

Intestinal Region-Dependent Impact of NFκB-Nrf Crosstalk in Myenteric Neurons and Adjacent Muscle Cells in Type 1 Diabetic Rats

 ,
,  , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Model
2.2. Tissue Handling
2.3. RNA Extraction, Reverse Transcription and Quantitative PCR
2.4. Primers
2.5. Measurement of Tissue NFκB and Nrf2 Concentrations
2.6. Bradford Protein Micromethod
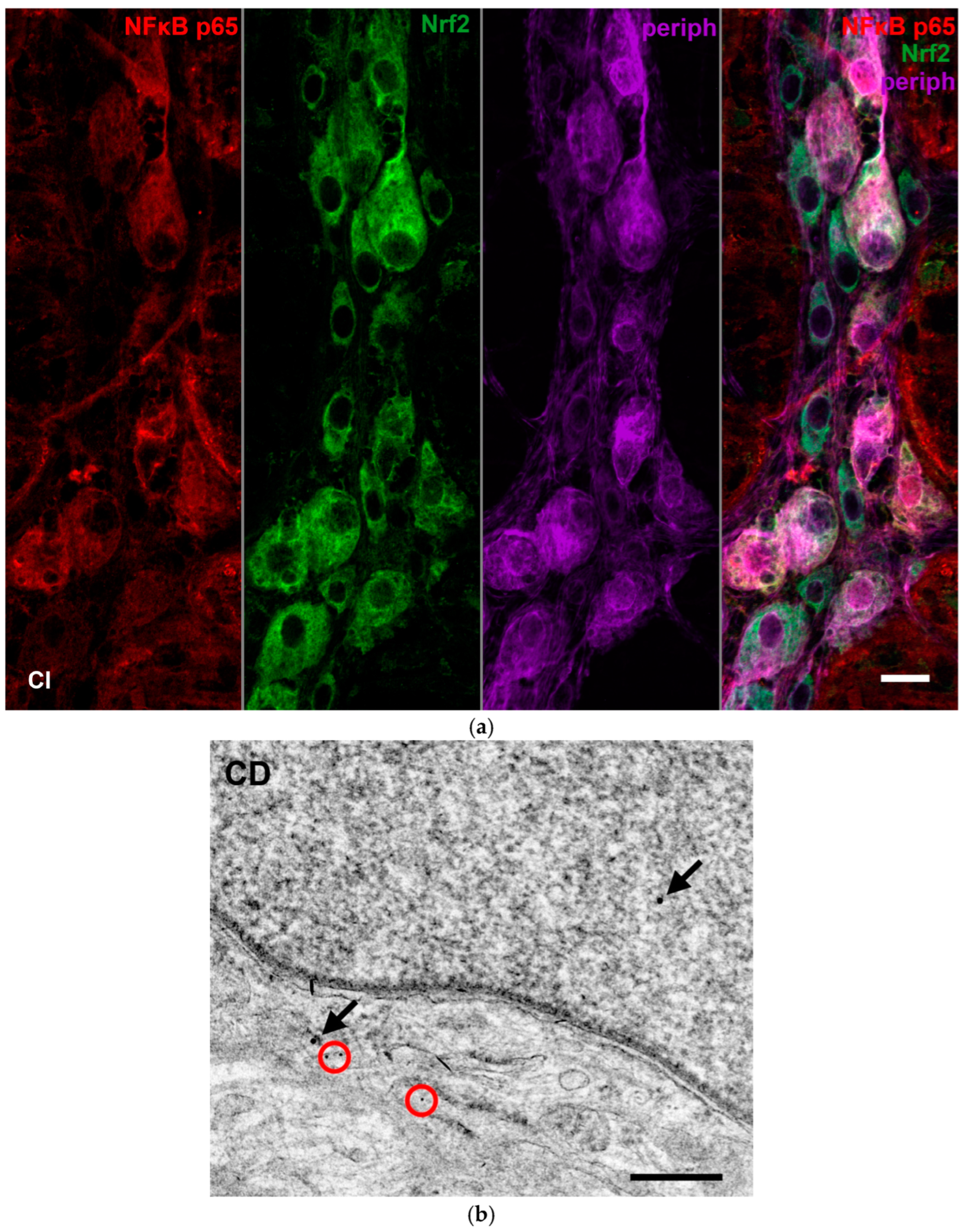
2.7. Fluorescent Immunohistochemistry
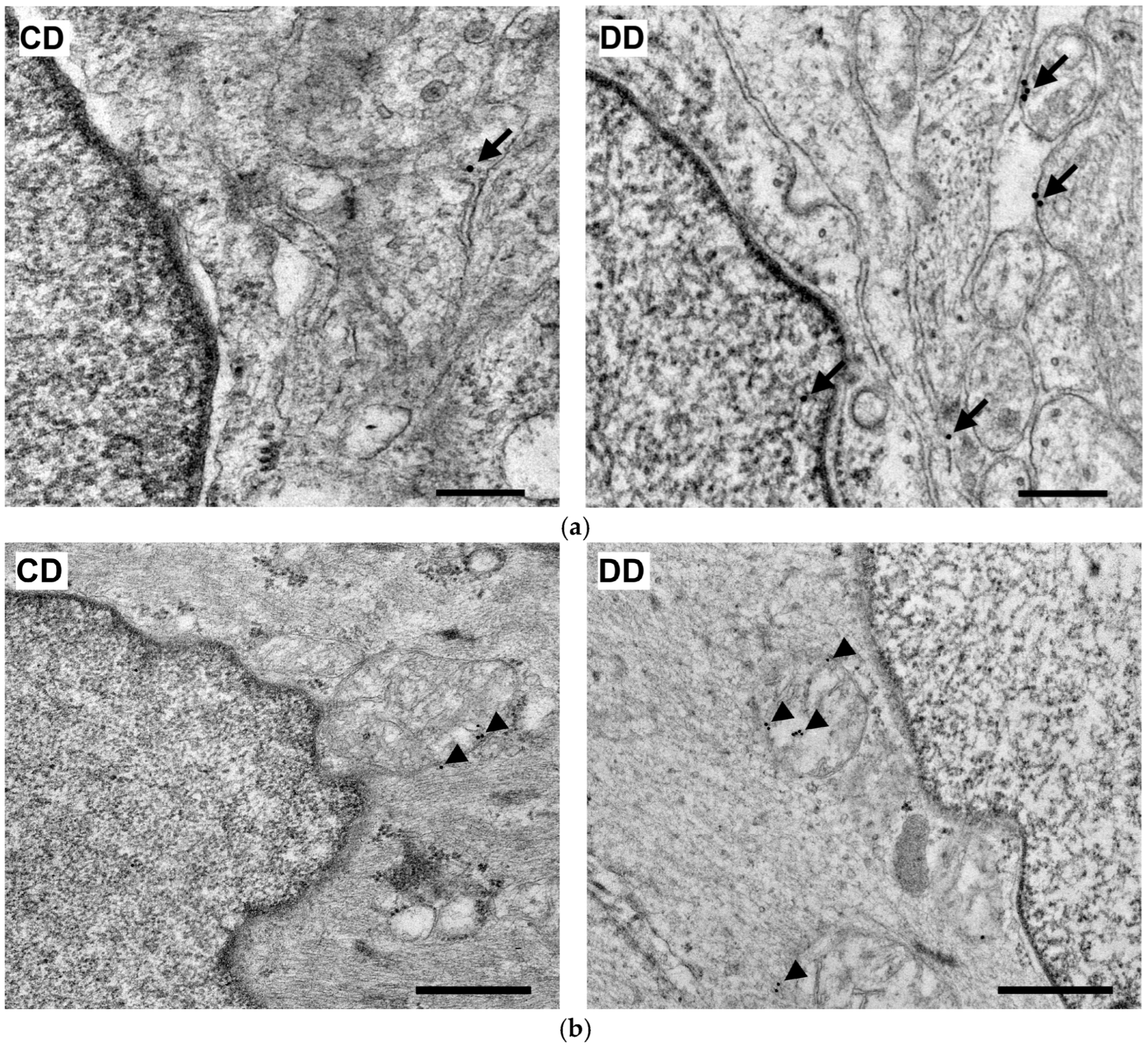
2.8. Quantitative Post-Embedding Immunohistochemistry
2.9. Statistical Analysis
3. Results
3.1. Disease Characteristics of Type 1 Diabetic Rats
3.2. Diabetic Alterations of Intestinal NFκB Expression
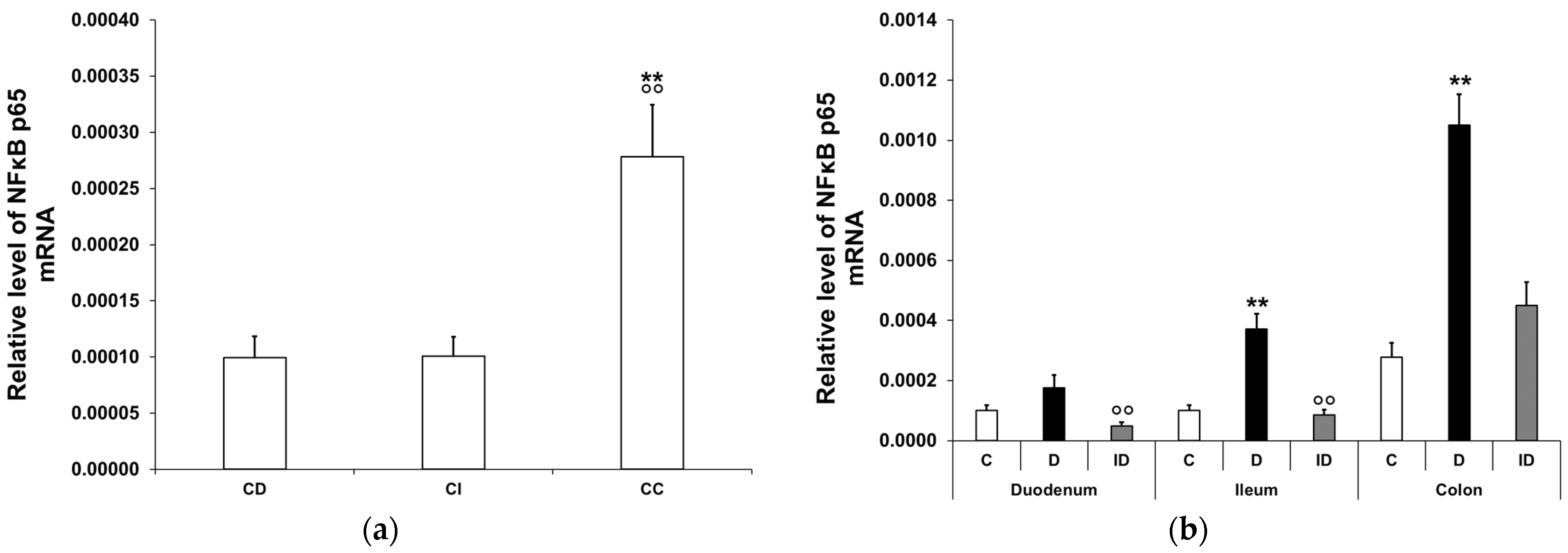
3.2.1. Quantitative Changes in NFκB p65 mRNA Expression
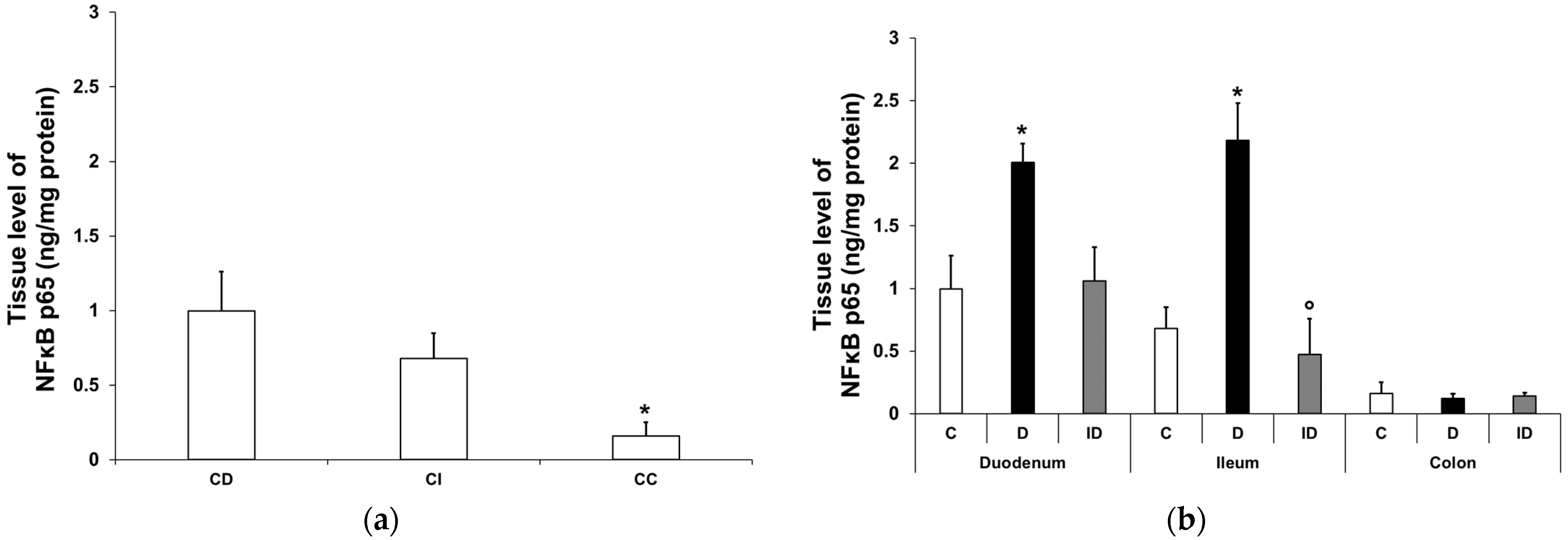
3.2.2. NFκB p65 Protein Level in Smooth Muscle/Myenteric Plexus Homogenates
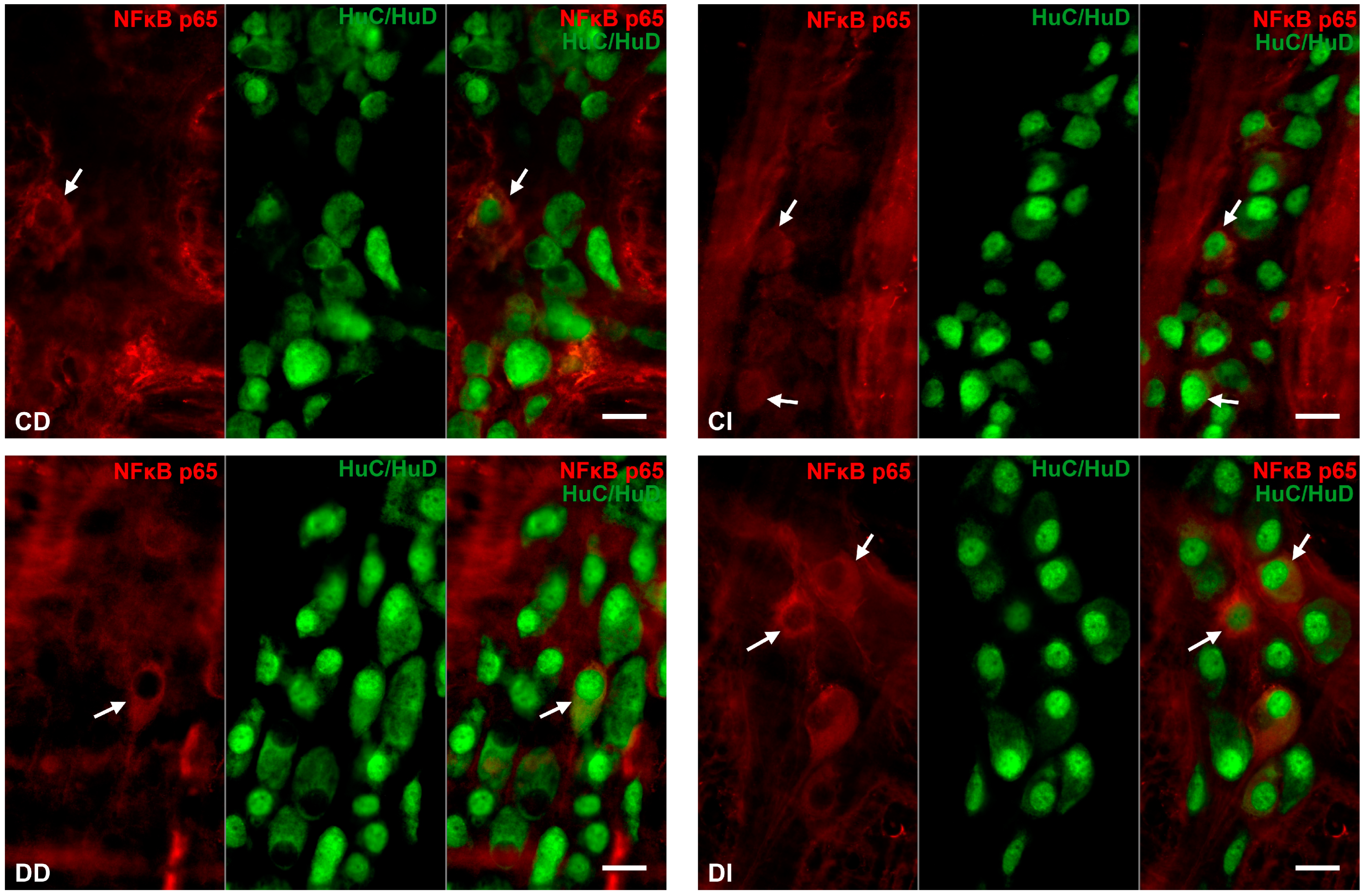
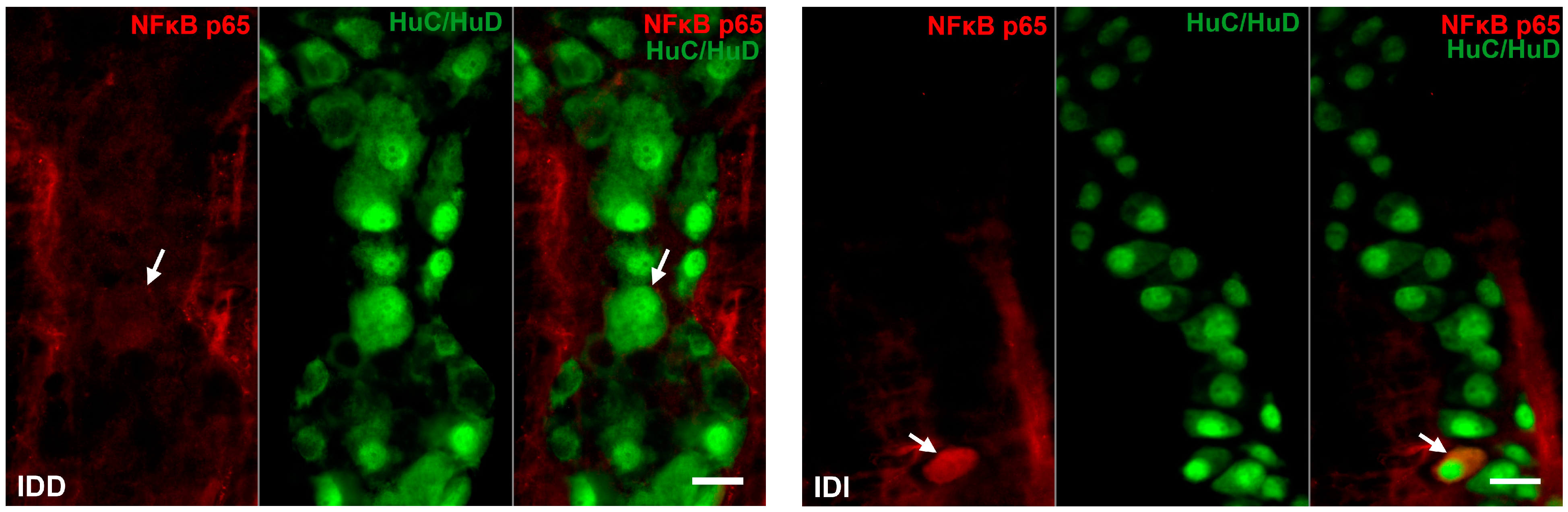
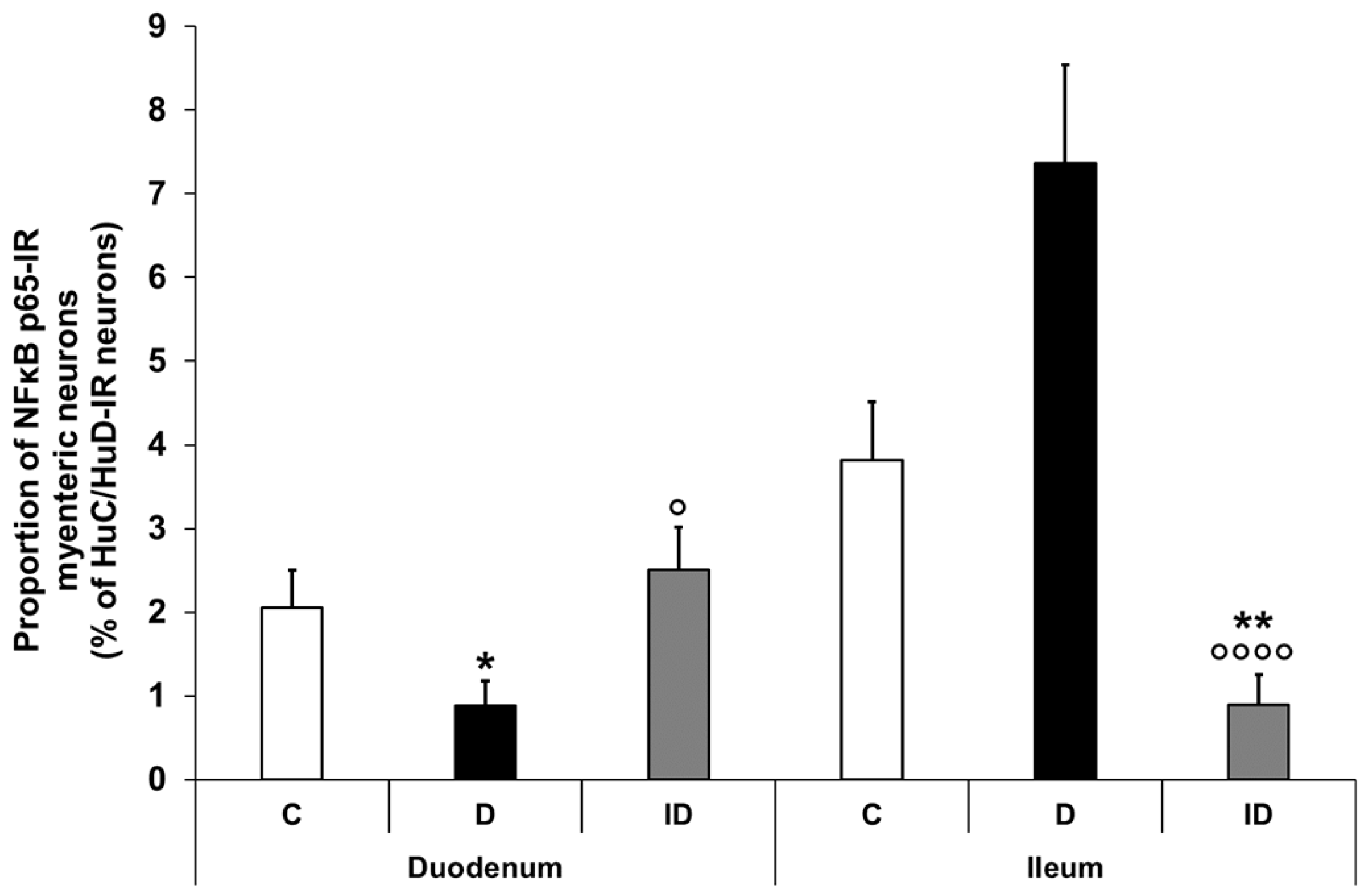
3.2.3. Proportion of NFκB p65-Immunoreactive Myenteric Neurons
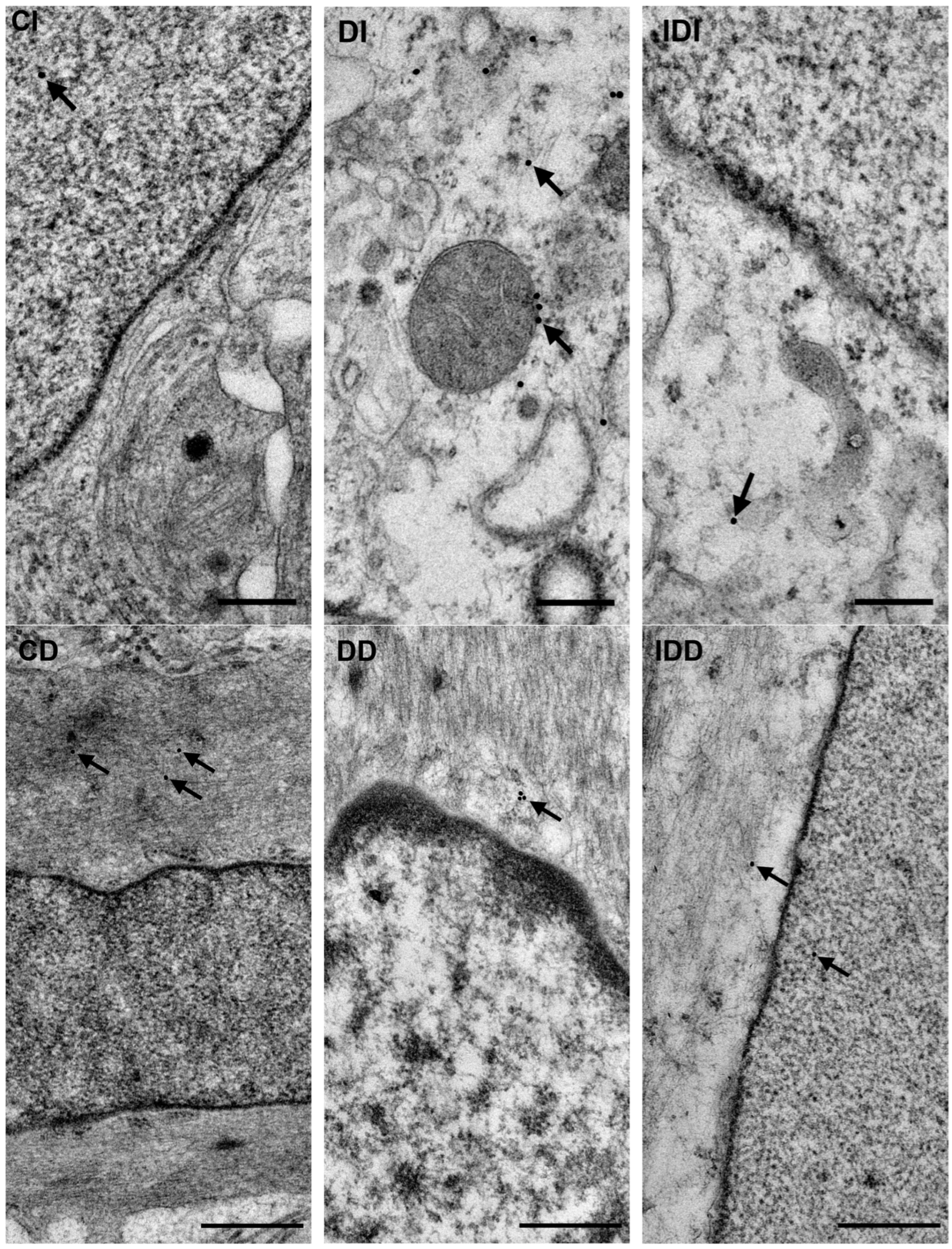
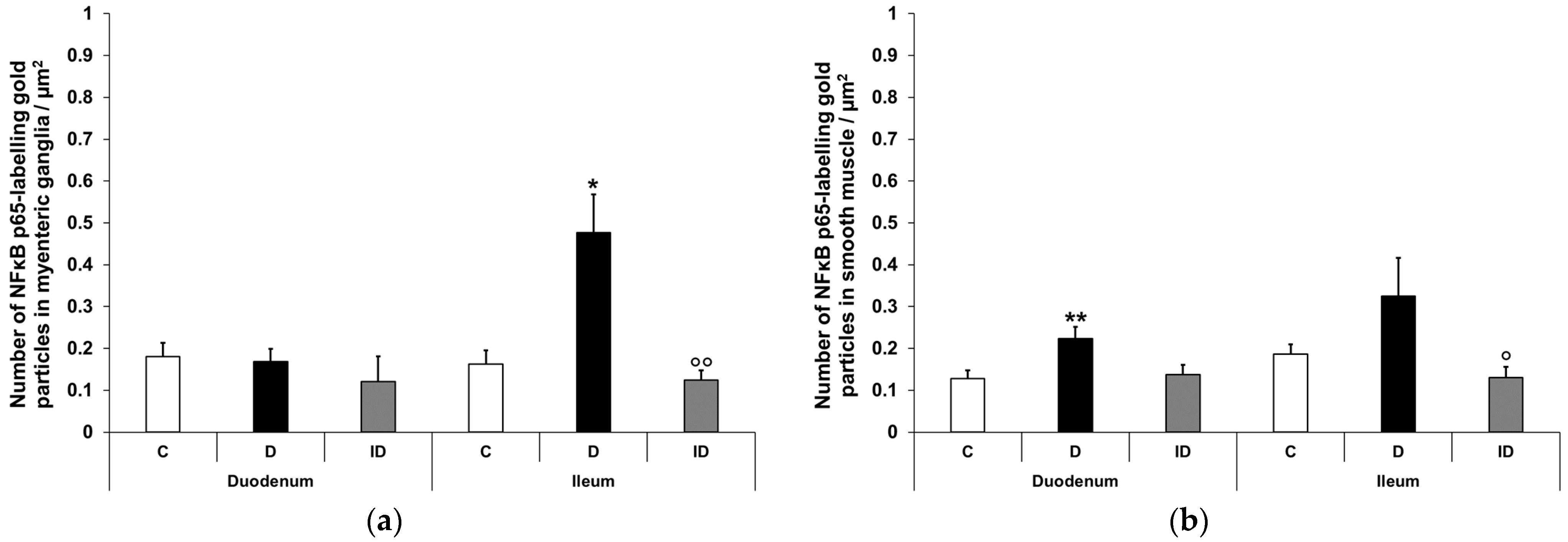
3.2.4. Quantitative Distribution of NFκB p65-Labelling Gold Particles in Myenteric Ganglia and Intestinal Smooth Muscle
3.3. Diabetic Alterations of Intestinal Nrf2 Expression
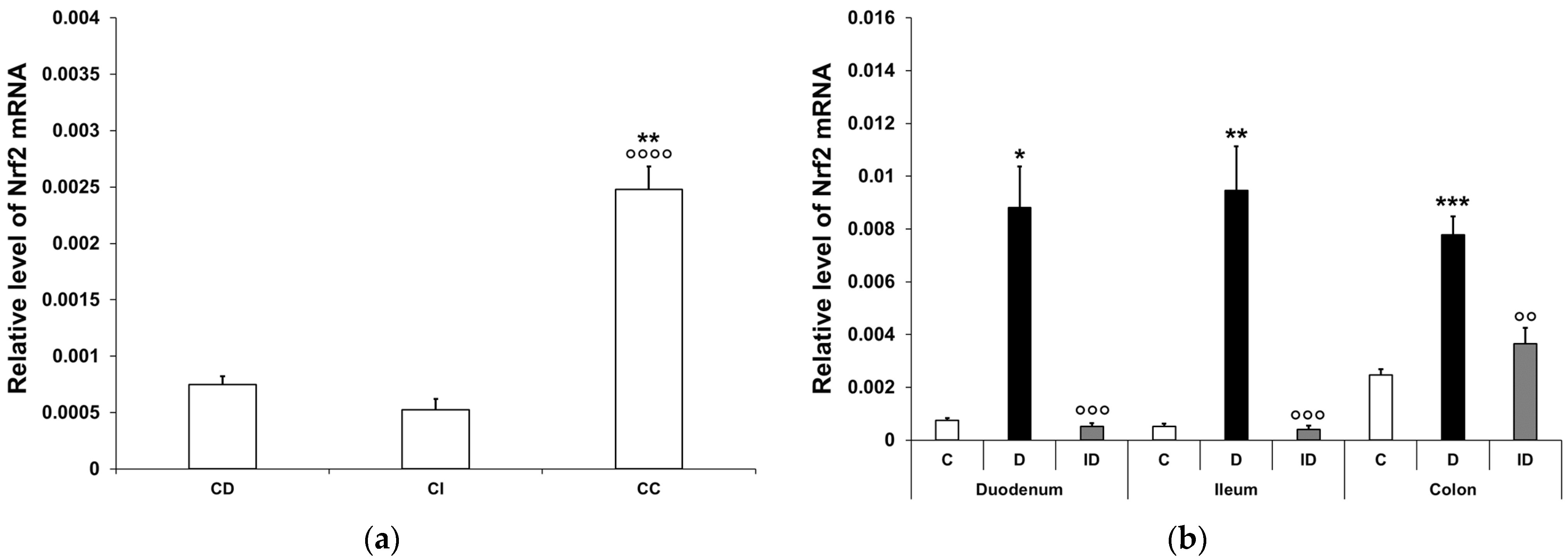
3.3.1. Quantitative Changes in Nrf2 mRNA Expression
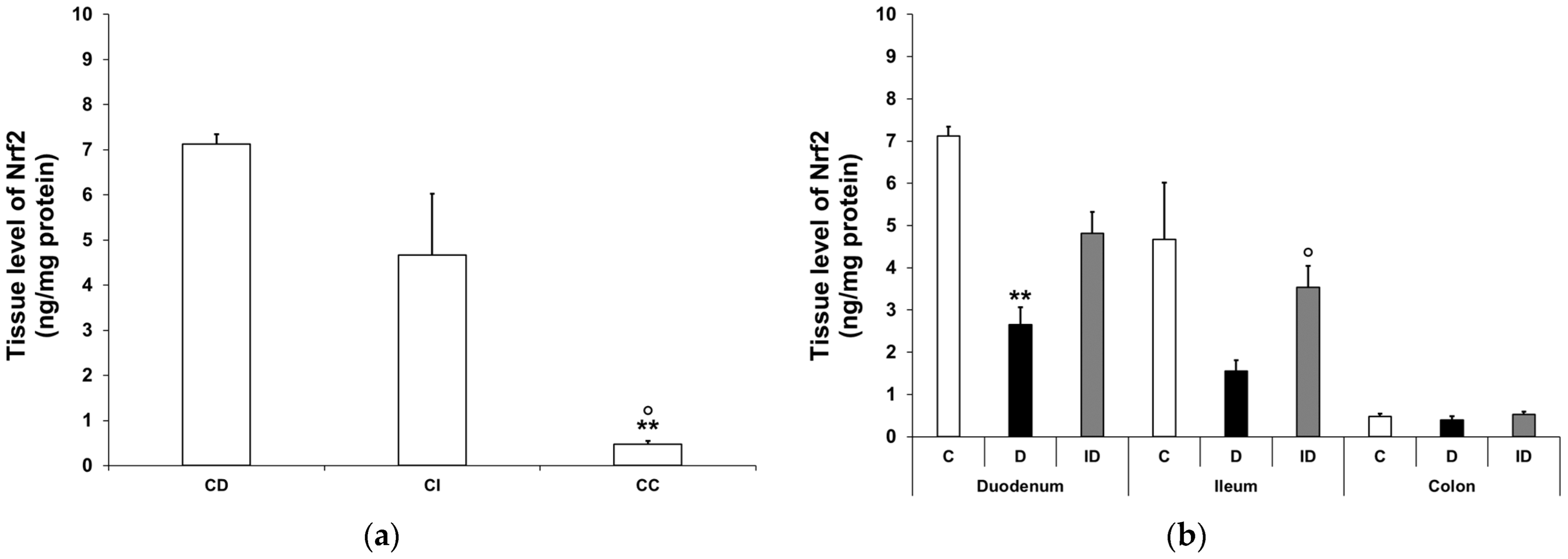
3.3.2. Nrf2 Protein Level in Smooth Muscle/Myenteric Plexus Homogenates
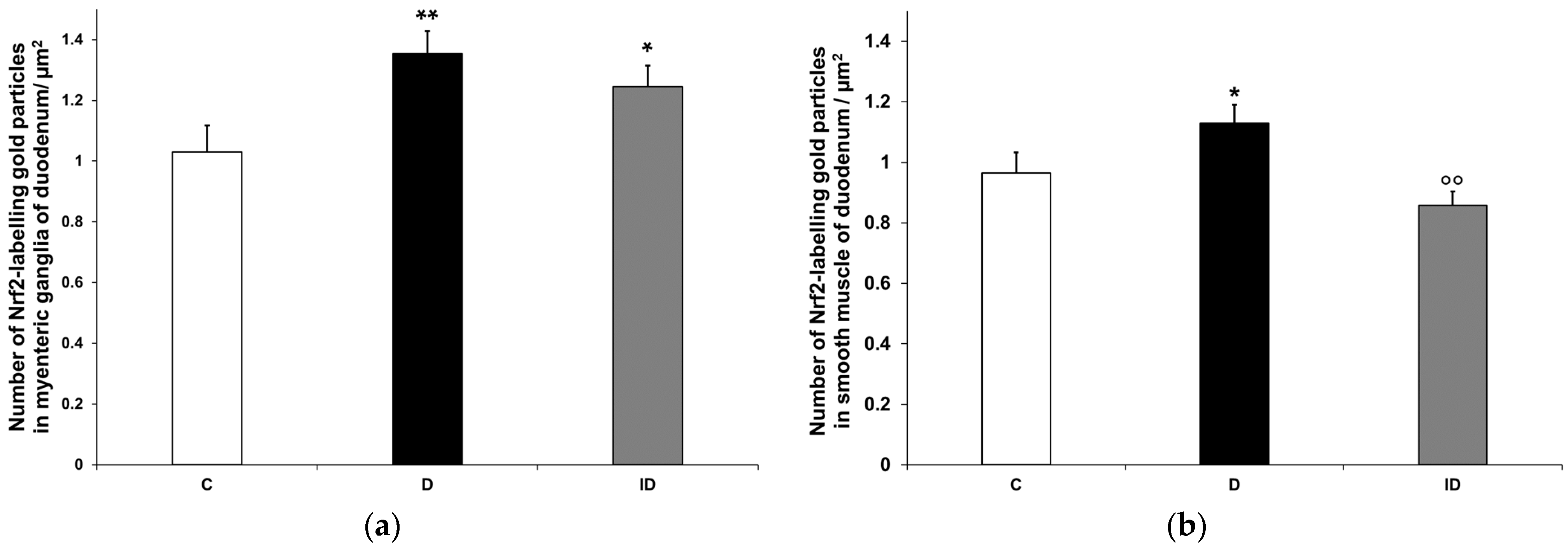
3.3.3. Quantitative Evaluation of Nrf2 Immunohistochemistry in Myenteric Ganglia and Intestinal Smooth Muscle
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Diabetes. Available online: https://www.who.int/news-room/fact-sheets/detail/diabetes (accessed on 16 September 2024).
- Modarelli, R.; Brown, L.; Boyd, J.; Podd, B.; Willis, Z.; Levenson, A. Severe Multiple Organ Failure as a Consequence of Diabetic Ketoacidosis in an Adolescent with New-Onset Type 1 Diabetes: A Case Report. SAGE Open Med. Case Rep. 2023, 11, 2050313X231190004. [Google Scholar] [CrossRef] [PubMed]
- Avagimyan, A.; Fogacci, F.; Pogosova, N.; Kakrurskiy, L.; Kogan, E.; Urazova, O.; Kobalava, Z.; Mikhaleva, L.; Vandysheva, R.; Zarina, G.; et al. Diabetic Cardiomyopathy: 2023 Update by the International Multidisciplinary Board of Experts. Curr. Probl. Cardiol. 2024, 49, 102052. [Google Scholar] [CrossRef] [PubMed]
- Shivam; Gupta, A.K.; Kumar, S. Current Concepts in the Molecular Mechanisms and Management ofDiabetic Neuropathy by Pharmacotherapeutics and Natural Compounds. Cent. Nerv. Syst. Agents Med. Chem. 2024, 24, 264–280. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Zhang, C. Recent Advances in the Management of Diabetic Kidney Disease: Slowing Progression. Int. J. Mol. Sci. 2024, 25, 3086. [Google Scholar] [CrossRef] [PubMed]
- Izbéki, F.; Wittman, T.; Rosztóczy, A.; Linke, N.; Bódi, N.; Fekete, É.; Bagyánszki, M. Immediate Insulin Treatment Prevents Gut Motility Alterations and Loss of Nitrergic Neurons in the Ileum and Colon of Rats with Streptozotocin-Induced Diabetes. Diabetes Res. Clin. Pract. 2008, 80, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Bagyánszki, M.; Bódi, N. Key Elements Determining the Intestinal Region-Specific Environment of Enteric Neurons in Type 1 Diabetes. World J. Gastroenterol. 2023, 29, 2704–2716. [Google Scholar] [CrossRef] [PubMed]
- Ruder, B.; Atreya, R.; Becker, C. Tumour Necrosis Factor Alpha in Intestinal Homeostasis and Gut Related Diseases. Int. J. Mol. Sci. 2019, 20, 1887. [Google Scholar] [CrossRef]
- Jarret, A.; Jackson, R.; Duizer, C.; Healy, M.E.; Zhao, J.; Rone, J.M.; Bielecki, P.; Sefik, E.; Roulis, M.; Rice, T.; et al. Enteric Nervous System-Derived IL-18 Orchestrates Mucosal Barrier Immunity. Cell 2020, 180, 50–63. [Google Scholar] [CrossRef]
- Wei, H.-X.; Wang, B.; Li, B. IL-10 and IL-22 in Mucosal Immunity: Driving Protection and Pathology. Front. Immunol. 2020, 11, 1315. [Google Scholar] [CrossRef]
- Bessac, A.; Cani, P.D.; Meunier, E.; Dietrich, G.; Knauf, C. Inflammation and Gut-Brain Axis During Type 2 Diabetes: Focus on the Crosstalk Between Intestinal Immune Cells and Enteric Nervous System. Front. Neurosci. 2018, 12, 725. [Google Scholar] [CrossRef]
- Wang, H.; Foong, J.P.P.; Harris, N.L.; Bornstein, J.C. Enteric Neuroimmune Interactions Coordinate Intestinal Responses in Health and Disease. Mucosal Immunol. 2022, 15, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Dolga, A.M.; Granic, I.; Blank, T.; Knaus, H.; Spiess, J.; Luiten, P.G.M.; Eisel, U.L.M.; Nijholt, I.M. TNF-α-mediates Neuroprotection against Glutamate-induced Excitotoxicity via NF-κB-dependent Up-regulation of KCa 2.2 Channels. J. Neurochem. 2008, 107, 1158–1167. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, S.; Vargas, J.; Hoffmann, A. Signaling via the NFκB System. Wiley Interdiscip. Rev. Syst. Biol. Med. 2016, 8, 227–241. [Google Scholar] [CrossRef]
- Alam, M.; Javed, M.N.; Najmi, A.K.; Ahmad, F.J.; Imam, S.S.; Akhtar, M. Thymoquinone Lipid Nanoparticles Cut the Gordian Knots of Depression via NeuroprotectiveBDNF and Downregulation of Neuro-Inflammatory NF-κB, IL-6, andTNF-α in LPS Treated Rats. Curr. Drug Metab. 2021, 22, 978–988. [Google Scholar] [CrossRef] [PubMed]
- Wardyn, J.D.; Ponsford, A.H.; Sanderson, C.M. Dissecting Molecular Cross-Talk between Nrf2 and NF-κB Response Pathways. Biochem. Soc. Trans. 2015, 43, 621–626. [Google Scholar] [CrossRef]
- Zhang, H.; Sun, S.-C. NF-κB in Inflammation and Renal Diseases. Cell Biosci. 2015, 5, 63. [Google Scholar] [CrossRef]
- Baldwin, A.S. THE NF-κB AND IκB PROTEINS: New Discoveries and Insights. Annu. Rev. Immunol. 1996, 14, 649–681. [Google Scholar] [CrossRef]
- Dresselhaus, E.C.; Meffert, M.K. Cellular Specificity of NF-κB Function in the Nervous System. Front. Immunol. 2019, 10, 1043. [Google Scholar] [CrossRef] [PubMed]
- Giridharan, S.; Srinivasan, M. Mechanisms of NF-κB P65 and Strategies for Therapeutic Manipulation. J. Inflamm. Res. 2018, 11, 407–419. [Google Scholar] [CrossRef]
- Patel, S.; Santani, D. Role of NF-κB in the Pathogenesis of Diabetes and Its Associated Complications. Pharmacol. Rep. 2009, 61, 595–603. [Google Scholar] [CrossRef]
- Heimberg, H.; Heremans, Y.; Jobin, C.; Leemans, R.; Cardozo, A.K.; Darville, M.; Eizirik, D.L. Inhibition of Cytokine-Induced NF-κB Activation by Adenovirus-Mediated Expression of a NF-κB Super-Repressor Prevents β-Cell Apoptosis. Diabetes 2001, 50, 2219–2224. [Google Scholar] [CrossRef] [PubMed]
- Eldor, R.; Yeffet, A.; Baum, K.; Doviner, V.; Amar, D.; Ben-Neriah, Y.; Christofori, G.; Peled, A.; Carel, J.C.; Boitard, C.; et al. Conditional and Specific NF-κB Blockade Protects Pancreatic Beta Cells from Diabetogenic Agents. Proc. Natl. Acad. Sci. USA 2006, 103, 5072–5077. [Google Scholar] [CrossRef] [PubMed]
- Norlin, S.; Ahlgren, U.; Edlund, H. Nuclear Factor-κB Activity in β-Cells Is Required for Glucose-Stimulated Insulin Secretion. Diabetes 2005, 54, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Mattson, M.P.; Meffert, M.K. Roles for NF-κB in Nerve Cell Survival, Plasticity, and Disease. Cell Death Differ. 2006, 13, 852–860. [Google Scholar] [CrossRef]
- Qin, Z.; Tao, L.; Chen, X. Dual Roles of NF-κB in Cell Survival and Implications of NF-κB Inhibitors in Neuroprotective Therapy. Acta Pharmacol. Sin. 2007, 28, 1859–1872. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Singh, T.G. Role of Nuclear Factor Kappa B (NF-κB) Signalling in Neurodegenerative Diseases: An Mechanistic Approach. Curr. Neuropharmacol. 2020, 18, 918–935. [Google Scholar] [CrossRef]
- Jeffrey, S.; Samraj, P.I.; Raj, B.S. The Role of Alpha-Lipoic Acid Supplementation in the Prevention of Diabetes Complications: A Comprehensive Review of Clinical Trials. Curr. Diabetes Rev. 2021, 17, 87–98. [Google Scholar] [CrossRef]
- Guo, Q.; Jin, Y.; Chen, X.; Ye, X.; Shen, X.; Lin, M.; Zeng, C.; Zhou, T.; Zhang, J. NF-κB in Biology and Targeted Therapy: New Insights and Translational Implications. Signal Transduct. Target. Ther. 2024, 9, 53. [Google Scholar] [CrossRef]
- Gao, W.; Guo, L.; Yang, Y.; Wang, Y.; Xia, S.; Gong, H.; Zhang, B.-K.; Yan, M. Dissecting the Crosstalk Between Nrf2 and NF-κB Response Pathways in Drug-Induced Toxicity. Front. Cell Dev. Biol. 2022, 9, 809952. [Google Scholar] [CrossRef]
- Kobayashi, A.; Ohta, T.; Yamamoto, M. Unique Function of the Nrf2–Keap1 Pathway in the Inducible Expression of Antioxidant and Detoxifying Enzymes. Methods Enzymol. 2004, 378, 273–286. [Google Scholar] [CrossRef]
- Yanaka, A. Role of NRF2 in Protection of the Gastrointestinal Tract against Oxidative Stress. J. Clin. Biochem. Nutr. 2018, 63, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Hashemi, M.; Zandieh, M.A.; Ziaolhagh, S.; Mojtabavi, S.; Sadi, F.H.; Koohpar, Z.K.; Ghanbarirad, M.; Haghighatfard, A.; Behroozaghdam, M.; Khorrami, R.; et al. Nrf2 Signaling in Diabetic Nephropathy, Cardiomyopathy and Neuropathy: Therapeutic Targeting, Challenges and Future Prospective. Biochim. Et Biophys. Acta (BBA)-Mol. Basis Dis. 2023, 1869, 166714. [Google Scholar] [CrossRef] [PubMed]
- Bódi, N.; Chandrakumar, L.; Al Doghmi, A.; Mezei, D.; Szalai, Z.; Barta, B.P.; Balázs, J.; Bagyánszki, M. Intestinal Region-Specific and Layer-Dependent Induction of TNFα in Rats with Streptozotocin-Induced Diabetes and after Insulin Replacement. Cells 2021, 10, 2410. [Google Scholar] [CrossRef] [PubMed]
- Al Doghmi, A.; Barta, B.P.; Egyed-Kolumbán, A.; Onhausz, B.; Kiss, S.; Balázs, J.; Szalai, Z.; Bagyánszki, M.; Bódi, N. Gut Region-Specific Interleukin 1β Induction in Different Myenteric Neuronal Subpopulations of Type 1 Diabetic Rats. Int. J. Mol. Sci. 2023, 24, 5804. [Google Scholar] [CrossRef]
- Barta, B.P.; Onhausz, B.; Doghmi, A.A.; Szalai, Z.; Balázs, J.; Bagyánszki, M.; Bódi, N. Gut Region-Specific TNFR Expression: TNFR2 Is More Affected than TNFR1 in Duodenal Myenteric Ganglia of Diabetic Rats. World J. Diabetes 2023, 14, 48–61. [Google Scholar] [CrossRef]
- Chandrakumar, L.; Bagyánszki, M.; Szalai, Z.; Mezei, D.; Bódi, N. Diabetes-Related Induction of the Heme Oxygenase System and Enhanced Colocalization of Heme Oxygenase 1 and 2 with Neuronal Nitric Oxide Synthase in Myenteric Neurons of Different Intestinal Segments. Oxidative Med. Cell. Longev. 2017, 2017, 1–13. [Google Scholar] [CrossRef]
- Bódi, N.; Egyed-Kolumbán, A.; Onhausz, B.; Barta, B.P.; Doghmi, A.A.; Balázs, J.; Szalai, Z.; Bagyánszki, M. Intestinal Region-Dependent Alterations of Toll-Like Receptor 4 Expression in Myenteric Neurons of Type 1 Diabetic Rats. Biomedicines 2023, 11, 129. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Barta, B.P.; Onhausz, B.; AL Doghmi, A.; Egyed-Kolumbán, A.; Kiss, S.Z.; Szalai, Z.; Balázs, J.; Bagyánszki, M.; Bódi, N. Gut Region-Specific Expression of Nuclear Factor Kappa B P65 in the Myenteric Ganglia and Its Microenvironment of Streptozotocin Induced Diabetic Rats. In Proceedings of the 1st International Conference on Antioxidants, Barcelona, Spain, 10–12 May 2023. [Google Scholar]
- Stachon, T.; Latta, L.; Seitz, B.; Szentmáry, N. Hypoxic Stress Increases NF-κB and iNOS mRNA Expression in Normal, but Not in Keratoconus Corneal Fibroblasts. Graefe’s Arch. Clin. Exp. Ophthalmol. 2021, 259, 449–458. [Google Scholar] [CrossRef]
- Cuadrado, A.; Martín-Moldes, Z.; Ye, J.; Lastres-Becker, I. Transcription Factors NRF2 and NF-κB Are Coordinated Effectors of the Rho Family, GTP-Binding Protein RAC1 during Inflammation. J. Biol. Chem. 2014, 289, 15244–15258. [Google Scholar] [CrossRef]
- Luo, Q.; Cui, H.; Deng, H.; Kuang, P.; Liu, H.; Lu, Y.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; et al. Sodium Fluoride Induces Renal Inflammatory Responses by Activating NF-κB Signaling Pathway and Reducing Anti-Inflammatory Cytokine Expression in Mice. Oncotarget 2017, 8, 80192–80207. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Yang, X.-D.; Lamb, A.; Chen, L.-F. Posttranslational Modifications of NF-κB: Another Layer of Regulation for NF-κB Signaling Pathway. Cell. Signal. 2010, 22, 1282–1290. [Google Scholar] [CrossRef] [PubMed]
- Meier-Soelch, J.; Mayr-Buro, C.; Juli, J.; Leib, L.; Linne, U.; Dreute, J.; Papantonis, A.; Schmitz, M.L.; Kracht, M. Monitoring the Levels of Cellular NF-κB Activation States. Cancers 2021, 13, 5351. [Google Scholar] [CrossRef]
- Kaltschmidt, C.; Kaltschmidt, B.; Neumann, H.; Wekerle, H.; Baeuerle, P.A. Constitutive NF-Kappa B Activity in Neurons. Mol. Cell. Biol. 1994, 14, 3981–3992. [Google Scholar] [CrossRef]
- Kracht, M.; Müller-Ladner, U.; Schmitz, M.L. Mutual Regulation of Metabolic Processes and Proinflammatory NF-κB Signaling. J. Allergy Clin. Immunol. 2020, 146, 694–705. [Google Scholar] [CrossRef]
- Zhu, J.; Cynader, M.S.; Jia, W. TDP-43 Inhibits NF-κB Activity by Blocking P65 Nuclear Translocation. PLoS ONE 2015, 10, e0142296. [Google Scholar] [CrossRef]
- Fagerlund, R.; Kinnunen, L.; Köhler, M.; Julkunen, I.; Melén, K. NF-κB Is Transported into the Nucleus by Importin A3 and Importin A4. J. Biol. Chem. 2005, 280, 15942–15951. [Google Scholar] [CrossRef]
- Jancsó, Z.; Bódi, N.; Borsos, B.; Fekete, É.; Hermesz, E. Gut Region-Specific Accumulation of Reactive Oxygen Species Leads to Regionally Distinct Activation of Antioxidant and Apoptotic Marker Molecules in Rats with STZ-Induced Diabetes. Int. J. Biochem. Cell Biol. 2015, 62, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Song, M.-Y.; Kim, E.-K.; Moon, W.-S.; Park, J.-W.; Kim, H.-J.; So, H.-S.; Park, R.; Kwon, K.-B.; Park, B.-H. Sulforaphane Protects against Cytokine- and Streptozotocin-Induced β-Cell Damage by Suppressing the NF-κB Pathway. Toxicol. Appl. Pharmacol. 2009, 235, 57–67. [Google Scholar] [CrossRef]
- Sandireddy, R.; Yerra, V.G.; Komirishetti, P.; Areti, A.; Kumar, A. Fisetin Imparts Neuroprotection in Experimental Diabetic Neuropathy by Modulating Nrf2 and NF-κB Pathways. Cell. Mol. Neurobiol. 2016, 36, 883–892. [Google Scholar] [CrossRef]
- Zhang, W.; Yu, H.; Lin, Q.; Liu, X.; Cheng, Y.; Deng, B. Anti-Inflammatory Effect of Resveratrol Attenuates the Severity of Diabetic Neuropathy by Activating the Nrf2 Pathway. Aging 2021, 13, 10659–10671. [Google Scholar] [CrossRef] [PubMed]
- Bódi, N.; Barta, B.P.; Onhausz, B.; Doghmi, A.A.L.; Egyed-Kolumbán, A.; Szalai, Z.; Balázs, J.; Bagyánszki, M. Diabetes-Related Alterations in the Expression of NFκB and Its Negative Regulator in the Myenteric Ganglia and Neighbouring Muscular Environment of Rat Duodenum. In Proceedings of the 48th FEBS Congress, Milan, Italy, 29 June–3 July 2024. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primers (5′ → 3′) |
|---|---|
| NFκB (NM_199267) | CCGGAACTCTGGGAGCTGCC AGGCTAGGGTCAGCGTATGG |
| NRF2 (XM_006234397) | GCAACTCCAGAAGGAACAGG GGAATGTCTCTGCCAAAAGC |
| Body Weight (g) | Blood Glucose Concentration (mmol/L) | |||
|---|---|---|---|---|
| Initial | Final | Initial | Final (Average) | |
| Controls (n = 14) | 256.7 ± 17.68 | 467 ± 16.13 **** | 5.96 ± 0.26 | 5.92 ± 0.11 |
| Diabetics (n = 14) | 223.9 ± 7.54 | 320.9 ± 14.73 | 6.09 ± 0.37 | 29.46 ± 0.87 ****,oooo |
| Insulin-treated diabetics (n = 13) | 233.7 ± 9.44 | 463.1 ± 14.87 **** | 5.86 ± 0.35 | 12.98 ± 1.5 *,o |
| Myenteric Neurons | Smooth Muscle Cells | |||
|---|---|---|---|---|
| Perikarya | Nuclei | Cytoplasm | Nuclei | |
| Controls | 1.12 ± 0.16 | 0.95 ± 0.06 | 0.67 ± 0.04 | 1.27 ± 0.11 |
| Diabetics | 1.43 ± 0.09 ** | 1.28 ± 0.08 * | 0.92 ± 0.05 *** | 1.35 ± 0.10 |
| Insulin-treated diabetics | 1.32 ± 0.09 * | 1.16 ± 0.07 | 0.75 ± 0.04 ° | 0.97 ± 0.08 ° |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barta, B.P.; Onhausz, B.; Egyed-Kolumbán, A.; AL Doghmi, A.; Balázs, J.; Szalai, Z.; Ferencz, Á.; Hermesz, E.; Bagyánszki, M.; Bódi, N. Intestinal Region-Dependent Impact of NFκB-Nrf Crosstalk in Myenteric Neurons and Adjacent Muscle Cells in Type 1 Diabetic Rats. Biomedicines 2024, 12, 2347. https://doi.org/10.3390/biomedicines12102347
Barta BP, Onhausz B, Egyed-Kolumbán A, AL Doghmi A, Balázs J, Szalai Z, Ferencz Á, Hermesz E, Bagyánszki M, Bódi N. Intestinal Region-Dependent Impact of NFκB-Nrf Crosstalk in Myenteric Neurons and Adjacent Muscle Cells in Type 1 Diabetic Rats. Biomedicines. 2024; 12(10):2347. https://doi.org/10.3390/biomedicines12102347
Chicago/Turabian StyleBarta, Bence Pál, Benita Onhausz, Abigél Egyed-Kolumbán, Afnan AL Doghmi, János Balázs, Zita Szalai, Ágnes Ferencz, Edit Hermesz, Mária Bagyánszki, and Nikolett Bódi. 2024. "Intestinal Region-Dependent Impact of NFκB-Nrf Crosstalk in Myenteric Neurons and Adjacent Muscle Cells in Type 1 Diabetic Rats" Biomedicines 12, no. 10: 2347. https://doi.org/10.3390/biomedicines12102347
APA StyleBarta, B. P., Onhausz, B., Egyed-Kolumbán, A., AL Doghmi, A., Balázs, J., Szalai, Z., Ferencz, Á., Hermesz, E., Bagyánszki, M., & Bódi, N. (2024). Intestinal Region-Dependent Impact of NFκB-Nrf Crosstalk in Myenteric Neurons and Adjacent Muscle Cells in Type 1 Diabetic Rats. Biomedicines, 12(10), 2347. https://doi.org/10.3390/biomedicines12102347