

Cannabinerol (CBNR) Influences Synaptic Genes Associated with Cytoskeleton and Ion Channels in NSC-34 Cell Line: A Transcriptomic Study
 , , and
, , and
Abstract
1. Introduction
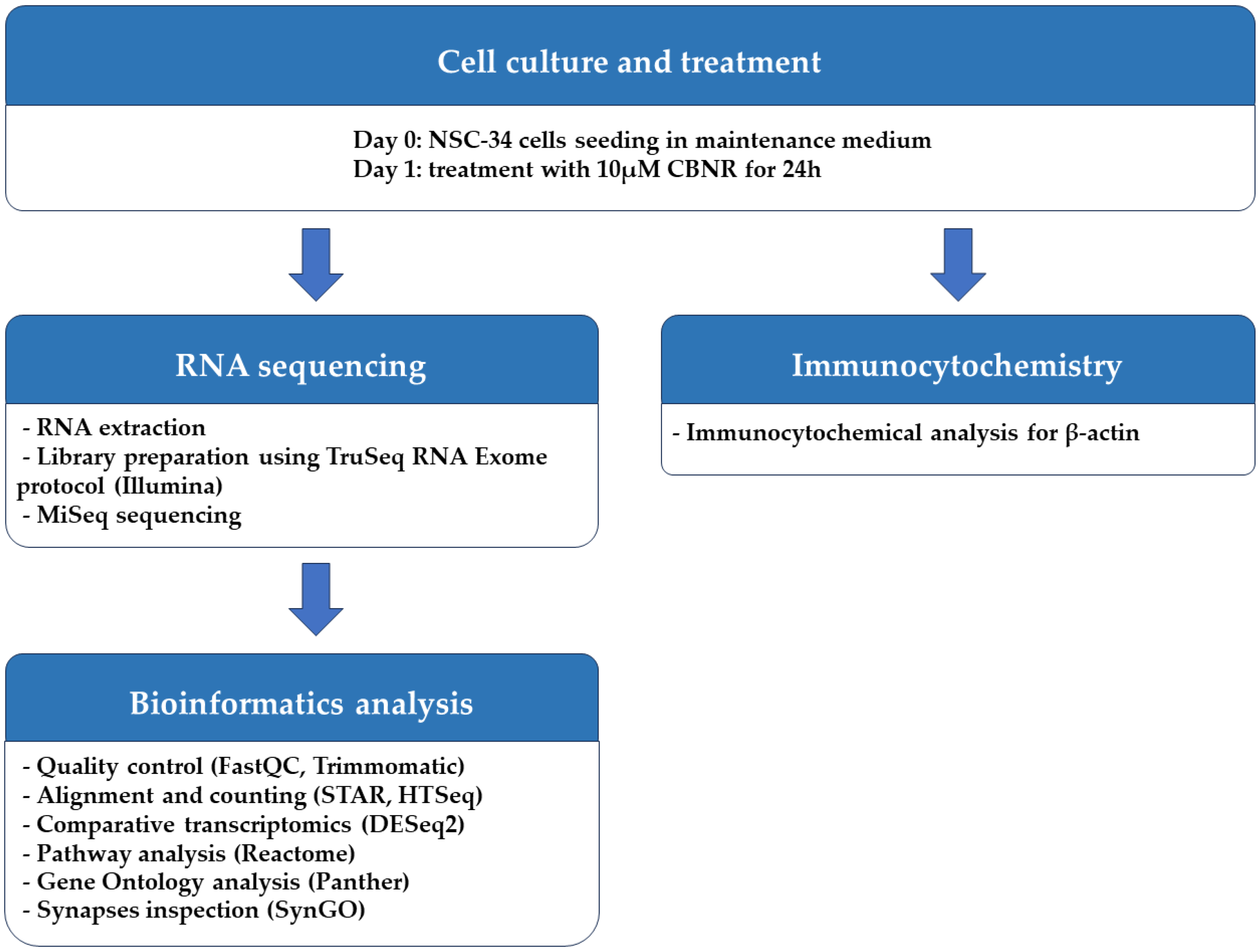
2. Materials and Methods
2.1. Cell Culture and Treatment
2.2. RNA Extraction and Library Preparation
2.3. Bioinformatics Analysis
2.4. Immunocytochemistry
3. Results
3.1. CBNR Modulates Synaptic Genes
3.2. CBNR Modulates Genes Associated with Cytoskeleton and Ion Channels
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ding, C.; Wu, Y.; Chen, X.; Chen, Y.; Wu, Z.; Lin, Z.; Kang, D.; Fang, W.; Chen, F. Global, regional, and national burden and attributable risk factors of neurological disorders: The Global Burden of Disease study 1990–2019. Front. Public Health 2022, 10, 952161. [Google Scholar] [CrossRef] [PubMed]
- Van Schependom, J.; D’Haeseleer, M. Advances in Neurodegenerative Diseases. J. Clin. Med. 2023, 12, 1709. [Google Scholar] [CrossRef] [PubMed]
- Dejanovic, B.; Sheng, M.; Hanson, J.E. Targeting synapse function and loss for treatment of neurodegenerative diseases. Nat. Rev. Drug Discov. 2024, 23, 23–42. [Google Scholar] [CrossRef]
- Stone, N.L.; Murphy, A.J.; England, T.J.; O’Sullivan, S.E. A systematic review of minor phytocannabinoids with promising neuroprotective potential. Br. J. Pharmacol. 2020, 177, 4330–4352. [Google Scholar] [CrossRef] [PubMed]
- Pennypacker, S.D.; Romero-Sandoval, E.A. CBD and THC: Do They Complement Each Other Like Yin and Yang? Pharmacotherapy 2020, 40, 1152–1165. [Google Scholar] [CrossRef] [PubMed]
- Voicu, V.; Brehar, F.M.; Toader, C.; Covache-Busuioc, R.A.; Corlatescu, A.D.; Bordeianu, A.; Costin, H.P.; Bratu, B.G.; Glavan, L.A.; Ciurea, A.V. Cannabinoids in Medicine: A Multifaceted Exploration of Types, Therapeutic Applications, and Emerging Opportunities in Neurodegenerative Diseases and Cancer Therapy. Biomolecules 2023, 13, 1388. [Google Scholar] [CrossRef] [PubMed]
- Mori, M.A.; Meyer, E.; Soares, L.M.; Milani, H.; Guimaraes, F.S.; de Oliveira, R.M.W. Cannabidiol reduces neuroinflammation and promotes neuroplasticity and functional recovery after brain ischemia. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2017, 75, 94–105. [Google Scholar] [CrossRef]
- Pazos, M.R.; Cinquina, V.; Gomez, A.; Layunta, R.; Santos, M.; Fernandez-Ruiz, J.; Martinez-Orgado, J. Cannabidiol administration after hypoxia-ischemia to newborn rats reduces long-term brain injury and restores neurobehavioral function. Neuropharmacology 2012, 63, 776–783. [Google Scholar] [CrossRef]
- Valdeolivas, S.; Navarrete, C.; Cantarero, I.; Bellido, M.L.; Munoz, E.; Sagredo, O. Neuroprotective properties of cannabigerol in Huntington’s disease: Studies in R6/2 mice and 3-nitropropionate-lesioned mice. Neurother. J. Am. Soc. Exp. Neurother. 2015, 12, 185–199. [Google Scholar] [CrossRef] [PubMed]
- Gugliandolo, A.; Silvestro, S.; Chiricosta, L.; Pollastro, F.; Bramanti, P.; Mazzon, E. The Transcriptomic Analysis of NSC-34 Motor Neuron-Like Cells Reveals That Cannabigerol Influences Synaptic Pathways: A Comparative Study with Cannabidiol. Life 2020, 10, 227. [Google Scholar] [CrossRef]
- Cashman, N.R.; Durham, H.D.; Blusztajan, J.K.; Oda, K.; Tabira, T.; Shaw, I.T.; Dahrouge, S.; Antel, J.P. Neuroblastoma X Spinal-Cord (Nsc) Hybrid Cell-Lines Resemble Developing Motor Neurons. Dev. Dyn. 1992, 194, 209–221. [Google Scholar] [CrossRef] [PubMed]
- Caprioglio, D.; Amin, H.I.M.; Taglialatela-Scafati, O.; Munoz, E.; Appendino, G. Minor Phytocannabinoids: A Misleading Name but a Promising Opportunity for Biomedical Research. Biomolecules 2022, 12, 1084. [Google Scholar] [CrossRef]
- Anchesi, I.; Schepici, G.; Chiricosta, L.; Gugliandolo, A.; Salamone, S.; Caprioglio, D.; Pollastro, F.; Mazzon, E. Delta(8)-THC Induces Up-Regulation of Glutamatergic Pathway Genes in Differentiated SH-SY5Y: A Transcriptomic Study. Int. J. Mol. Sci. 2023, 24, 9486. [Google Scholar] [CrossRef] [PubMed]
- Valeri, A.; Chiricosta, L.; Gugliandolo, A.; Pollastro, F.; Salamone, S.; Zingale, V.D.; Silvestro, S.; Mazzon, E. Cannabinerol and NSC-34 Transcriptomic Analysis: Is the Dose Who Makes Neuronal Differentiation? Int. J. Mol. Sci. 2022, 23, 7541. [Google Scholar] [CrossRef]
- Landeira, B.S.; Santana, T.; Araujo, J.A.M.; Tabet, E.I.; Tannous, B.A.; Schroeder, T.; Costa, M.R. Activity-Independent Effects of CREB on Neuronal Survival and Differentiation during Mouse Cerebral Cortex Development. Cereb. Cortex 2018, 28, 538–548. [Google Scholar] [CrossRef] [PubMed]
- Alberini, C.M. Transcription factors in long-term memory and synaptic plasticity. Physiol. Rev. 2009, 89, 121–145. [Google Scholar] [CrossRef]
- Durante, C.; Anceschi, L.; Brighenti, V.; Caroli, C.; Afezolli, C.; Marchetti, A.; Cocchi, M.; Salamone, S.; Pollastro, F.; Pellati, F. Application of experimental design in HPLC method optimisation for the simultaneous determination of multiple bioactive cannabinoids. J. Pharm. Biomed. Anal. 2022, 221, 115037. [Google Scholar] [CrossRef] [PubMed]
- Taura, F.; Morimoto, S.; Shoyama, Y. Cannabinerolic Acid, a Cannabinoid from Cannabis-Sativa. Phytochemistry 1995, 39, 457–458. [Google Scholar] [CrossRef]
- Hanus, L.O.; Levy, R.; De La Vega, D.; Katz, L.; Roman, M.; Tomícek, P. The main cannabinoids content in hashish samples seized in Israel and Czech Republic. Isr. J. Plant Sci. 2016, 63, 182–190. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, M.; Jassal, B.; Stephan, R.; Milacic, M.; Rothfels, K.; Senff-Ribeiro, A.; Griss, J.; Sevilla, C.; Matthews, L.; Gong, C.; et al. The reactome pathway knowledgebase 2022. Nucleic Acids Res. 2022, 50, D687–D692. [Google Scholar] [CrossRef] [PubMed]
- Mi, H.; Ebert, D.; Muruganujan, A.; Mills, C.; Albou, L.P.; Mushayamaha, T.; Thomas, P.D. PANTHER version 16: A revised family classification, tree-based classification tool, enhancer regions and extensive API. Nucleic Acids Res. 2021, 49, D394–D403. [Google Scholar] [CrossRef] [PubMed]
- Koopmans, F.; van Nierop, P.; Andres-Alonso, M.; Byrnes, A.; Cijsouw, T.; Coba, M.P.; Cornelisse, L.N.; Farrell, R.J.; Goldschmidt, H.L.; Howrigan, D.P.; et al. SynGO: An Evidence-Based, Expert-Curated Knowledge Base for the Synapse. Neuron 2019, 103, 217–234 e214. [Google Scholar] [CrossRef]
- Cheever, T.R.; Ervasti, J.M. Actin isoforms in neuronal development and function. Int. Rev. Cell Mol. Biol. 2013, 301, 157–213. [Google Scholar] [CrossRef]
- Batool, S.; Raza, H.; Zaidi, J.; Riaz, S.; Hasan, S.; Syed, N.I. Synapse formation: From cellular and molecular mechanisms to neurodevelopmental and neurodegenerative disorders. J. Neurophysiol. 2019, 121, 1381–1397. [Google Scholar] [CrossRef]
- Gugliandolo, A.; Pollastro, F.; Grassi, G.; Bramanti, P.; Mazzon, E. In Vitro Model of Neuroinflammation: Efficacy of Cannabigerol, a Non-Psychoactive Cannabinoid. Int. J. Mol. Sci. 2018, 19, 1992. [Google Scholar] [CrossRef]
- Mammana, S.; Cavalli, E.; Gugliandolo, A.; Silvestro, S.; Pollastro, F.; Bramanti, P.; Mazzon, E. Could the Combination of Two Non-Psychotropic Cannabinoids Counteract Neuroinflammation? Effectiveness of Cannabidiol Associated with Cannabigerol. Medicina 2019, 55, 747. [Google Scholar] [CrossRef]
- Valeri, A.; Chiricosta, L.; Gugliandolo, A.; Pollastro, F.; Mazzon, E. Will Cannabigerol Trigger Neuroregeneration after a Spinal Cord Injury? An In Vitro Answer from NSC-34 Scratch-Injured Cells Transcriptome. Pharmaceuticals 2022, 15, 117. [Google Scholar] [CrossRef]
- Qaisar, R. Targeting neuromuscular junction to treat neuromuscular disorders. Life Sci. 2023, 333, 122186. [Google Scholar] [CrossRef] [PubMed]
- Hill, M. The neuromuscular junction disorders. J. Neurol. Neurosurg. Psychiatry 2003, 74 (Suppl. S2), ii32–ii37. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gugliandolo, A.; Blando, S.; Salamone, S.; Caprioglio, D.; Pollastro, F.; Mazzon, E.; Chiricosta, L. Delta(8)-THC Protects against Amyloid Beta Toxicity Modulating ER Stress In Vitro: A Transcriptomic Analysis. Int. J. Mol. Sci. 2023, 24, 6598. [Google Scholar] [CrossRef]
- Kim, J.; Choi, P.; Park, Y.T.; Kim, T.; Ham, J.; Kim, J.C. The Cannabinoids, CBDA and THCA, Rescue Memory Deficits and Reduce Amyloid-Beta and Tau Pathology in an Alzheimer’s Disease-like Mouse Model. Int. J. Mol. Sci. 2023, 24, 6827. [Google Scholar] [CrossRef]
- Yang, S.; Du, Y.; Zhao, X.; Tang, Q.; Su, W.; Hu, Y.; Yu, P. Cannabidiol Enhances Microglial Beta-Amyloid Peptide Phagocytosis and Clearance via Vanilloid Family Type 2 Channel Activation. Int. J. Mol. Sci. 2022, 23, 5367. [Google Scholar] [CrossRef] [PubMed]
- Hao, F.; Feng, Y. Cannabidiol (CBD) enhanced the hippocampal immune response and autophagy of APP/PS1 Alzheimer’s mice uncovered by RNA-seq. Life Sci. 2021, 264, 118624. [Google Scholar] [CrossRef]
- Kanchi, P.K.; Dasmahapatra, A.K. Destabilization of the Alzheimer’s amyloid-beta protofibrils by THC: A molecular dynamics simulation study. J. Mol. Graph. Model. 2021, 105, 107889. [Google Scholar] [CrossRef]
- Paskus, J.D.; Herring, B.E.; Roche, K.W. Kalirin and Trio: RhoGEFs in Synaptic Transmission, Plasticity, and Complex Brain Disorders. Trends Neurosci. 2020, 43, 505–518. [Google Scholar] [CrossRef]
- Terry-Lorenzo, R.T.; Torres, V.I.; Wagh, D.; Galaz, J.; Swanson, S.K.; Florens, L.; Washburn, M.P.; Waites, C.L.; Gundelfinger, E.D.; Reimer, R.J.; et al. Trio, a Rho Family GEF, Interacts with the Presynaptic Active Zone Proteins Piccolo and Bassoon. PLoS ONE 2016, 11, e0167535. [Google Scholar] [CrossRef]
- Montenegro-Venegas, C.; Guhathakurta, D.; Pina-Fernandez, E.; Andres-Alonso, M.; Plattner, F.; Gundelfinger, E.D.; Fejtova, A. Bassoon controls synaptic vesicle release via regulation of presynaptic phosphorylation and cAMP. EMBO Rep. 2022, 23, e53659. [Google Scholar] [CrossRef] [PubMed]
- Waites, C.L.; Leal-Ortiz, S.A.; Okerlund, N.; Dalke, H.; Fejtova, A.; Altrock, W.D.; Gundelfinger, E.D.; Garner, C.C. Bassoon and Piccolo maintain synapse integrity by regulating protein ubiquitination and degradation. EMBO J. 2013, 32, 954–969. [Google Scholar] [CrossRef]
- Hodges, J.L.; Vilchez, S.M.; Asmussen, H.; Whitmore, L.A.; Horwitz, A.R. alpha-Actinin-2 mediates spine morphology and assembly of the post-synaptic density in hippocampal neurons. PLoS ONE 2014, 9, e101770. [Google Scholar] [CrossRef]
- Angliker, N.; Ruegg, M.A. In vivo evidence for mTORC2-mediated actin cytoskeleton rearrangement in neurons. Bioarchitecture 2013, 3, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Mironova, Y.A.; Giger, R.J. Where no synapses go: Gatekeepers of circuit remodeling and synaptic strength. Trends Neurosci. 2013, 36, 363–373. [Google Scholar] [CrossRef] [PubMed]
- Petrinovic, M.M.; Hourez, R.; Aloy, E.M.; Dewarrat, G.; Gall, D.; Weinmann, O.; Gaudias, J.; Bachmann, L.C.; Schiffmann, S.N.; Vogt, K.E.; et al. Neuronal Nogo-A negatively regulates dendritic morphology and synaptic transmission in the cerebellum. Proc. Natl. Acad. Sci. USA 2013, 110, 1083–1088. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Ong, S.E.; Bajjalieh, S. Huntingtin is associated with cytomatrix proteins at the presynaptic terminal. Mol. Cell. Neurosci. 2014, 63, 96–100. [Google Scholar] [CrossRef]
- Barron, J.C.; Hurley, E.P.; Parsons, M.P. Huntingtin and the Synapse. Front. Cell. Neurosci. 2021, 15, 689332. [Google Scholar] [CrossRef]
- Kim, K.; Yang, J.; Zhong, X.P.; Kim, M.H.; Kim, Y.S.; Lee, H.W.; Han, S.; Choi, J.; Han, K.; Seo, J.; et al. Synaptic removal of diacylglycerol by DGKzeta and PSD-95 regulates dendritic spine maintenance. EMBO J. 2009, 28, 1170–1179. [Google Scholar] [CrossRef]
- Pannaccione, A.; Piccialli, I.; Secondo, A.; Ciccone, R.; Molinaro, P.; Boscia, F.; Annunziato, L. The Na(+)/Ca(2+)exchanger in Alzheimer’s disease. Cell Calcium 2020, 87, 102190. [Google Scholar] [CrossRef]
- Liao, X.; Li, Y. Genetic associations between voltage-gated calcium channels and autism spectrum disorder: A systematic review. Mol. Brain 2020, 13, 96. [Google Scholar] [CrossRef] [PubMed]
- Andrade, A.; Brennecke, A.; Mallat, S.; Brown, J.; Gomez-Rivadeneira, J.; Czepiel, N.; Londrigan, L. Genetic Associations between Voltage-Gated Calcium Channels and Psychiatric Disorders. Int. J. Mol. Sci. 2019, 20, 3537. [Google Scholar] [CrossRef] [PubMed]
- Paoletti, P.; Bellone, C.; Zhou, Q. NMDA receptor subunit diversity: Impact on receptor properties, synaptic plasticity and disease. Nat. Rev. Neurosci. 2013, 14, 383–400. [Google Scholar] [CrossRef] [PubMed]
- Bunton-Stasyshyn, R.K.A.; Wagnon, J.L.; Wengert, E.R.; Barker, B.S.; Faulkner, A.; Wagley, P.K.; Bhatia, K.; Jones, J.M.; Maniaci, M.R.; Parent, J.M.; et al. Prominent role of forebrain excitatory neurons in SCN8A encephalopathy. Brain 2019, 142, 362–375. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, F.A.; Andrade, D.M. Myoclonus epilepsy and ataxia due to potassium channel mutation (MEAK) is caused by heterozygous KCNC1 mutations. Epileptic Disord. Int. Epilepsy J. Videotape 2016, 18, 135–138. [Google Scholar] [CrossRef] [PubMed]
- Jeremic, D.; Sanchez-Rodriguez, I.; Jimenez-Diaz, L.; Navarro-Lopez, J.D. Therapeutic potential of targeting G protein-gated inwardly rectifying potassium (GIRK) channels in the central nervous system. Pharmacol. Ther. 2021, 223, 107808. [Google Scholar] [CrossRef]
- Ames, J.B. L-Type Ca(2+) Channel Regulation by Calmodulin and CaBP1. Biomolecules 2021, 11, 1811. [Google Scholar] [CrossRef]
- Liang, Z.; Zhan, Y.; Shen, Y.; Wong, C.C.; Yates, J.R., 3rd; Plattner, F.; Lai, K.O.; Ip, N.Y. The pseudokinase CaMKv is required for the activity-dependent maintenance of dendritic spines. Nat. Commun. 2016, 7, 13282. [Google Scholar] [CrossRef]
- Oku, Y.; Huganir, R.L. AGAP3 and Arf6 regulate trafficking of AMPA receptors and synaptic plasticity. J. Neurosci. Off. J. Soc. Neurosci. 2013, 33, 12586–12598. [Google Scholar] [CrossRef]
- Llamosas, N.; Arora, V.; Vij, R.; Kilinc, M.; Bijoch, L.; Rojas, C.; Reich, A.; Sridharan, B.; Willems, E.; Piper, D.R.; et al. SYNGAP1 Controls the Maturation of Dendrites, Synaptic Function, and Network Activity in Developing Human Neurons. J. Neurosci. Off. J. Soc. Neurosci. 2020, 40, 7980–7994. [Google Scholar] [CrossRef]
- Yang, R.; Feng, X.; Arias-Cavieres, A.; Mitchell, R.M.; Polo, A.; Hu, K.; Zhong, R.; Qi, C.; Zhang, R.S.; Westneat, N.; et al. Upregulation of SYNGAP1 expression in mice and human neurons by redirecting alternative splicing. Neuron 2023, 111, 1637–1650.e35. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Borgmann-Winter, K.E.; Wang, H.Y.; Ray, R.; Willis, B.R.; Moberg, P.J.; Rawson, N.E.; Gur, R.E.; Turetsky, B.I.; Hahn, C.G. Altered G Protein Coupling in Olfactory Neuroepithelial Cells From Patients With Schizophrenia. Schizophr. Bull. 2016, 42, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Bellon, A.; Wegener, A.; Lescallette, A.R.; Valente, M.; Yang, S.K.; Gardette, R.; Matricon, J.; Mouaffak, F.; Watts, P.; Vimeux, L.; et al. Transdifferentiation of Human Circulating Monocytes Into Neuronal-Like Cells in 20 Days and Without Reprograming. Front. Mol. Neurosci. 2018, 11, 323. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Fold Change | Biological Process |
|---|---|---|
| Actn2 | 0.54 | structural constituent of postsynaptic actin cytoskeleton |
| Actr3 | −0.12 | postsynapse organization; postsynaptic actin cytoskeleton organization |
| Agap3 | 0.21 | regulation of postsynaptic membrane neurotransmitter receptor levels |
| Arfgap1 | 0.29 | - |
| Arhgdia | −0.18 | regulation of synaptic vesicle cycle |
| Bsn | 0.49 | regulation of synaptic vesicle cycle; structural constituent of active zone; synaptic vesicle clustering |
| Cabp1 | 0.41 | modification of postsynaptic actin cytoskeleton; postsynapse to nucleus signaling pathway |
| Cacna1b | 0.23 | regulation of calcium-dependent activation of synaptic vesicle fusion; voltage-gated calcium channel activity involved in regulation of presynaptic cytosolic calcium levels |
| Cacna1c | 0.14 | - |
| Cacnb1 | 0.36 | - |
| Cadm1 | −0.14 | maintenance of postsynaptic specialization structure; presynapse assembly; retrograde trans-synaptic signaling by trans-synaptic protein complex; synapse adhesion between pre- and postsynapse; synapse assembly |
| Camkv | 0.25 | modulation of chemical synaptic transmission; regulation of modification of postsynaptic structure |
| Canx | −0.15 | synaptic vesicle endocytosis |
| Cd2ap | −0.41 | synapse organization |
| Cltb | −0.22 | synaptic vesicle endocytosis |
| Cntnap1 | 0.28 | regulation of synapse maturation |
| Cpeb4 | 0.16 | - |
| Dbnl | 0.17 | postsynaptic actin cytoskeleton organization; structural constituent of postsynaptic actin cytoskeleton |
| Dgkz | 0.25 | maintenance of postsynaptic density structure |
| Dlgap3 | 0.23 | modification of postsynaptic structure; modulation of chemical synaptic transmission |
| Drosha | −0.14 | postsynaptic density |
| Dvl1 | 0.35 | postsynapse organization; presynapse assembly; regulation of postsynapse organization; regulation of synaptic vesicle exocytosis |
| Eif1ax | −0.15 | - |
| Eif4g2 | −0.14 | - |
| Eif5 | −0.13 | - |
| Gnb5 | 0.43 | - |
| Grin1 | 0.21 | ligand-gated ion channel activity involved in regulation of presynaptic membrane potential; transmitter-gated ion channel activity involved in regulation of postsynaptic membrane potential |
| Htt | 0.16 | postsynapse to nucleus signaling pathway |
| Igsf9b | 0.34 | synapse adhesion between pre- and postsynapse |
| Ina | 0.30 | postsynaptic modulation of chemical synaptic transmission; structural constituent of postsynaptic intermediate filament cytoskeleton |
| Kcnc1 | 1.23 | voltage-gated ion channel activity involved in regulation of presynaptic membrane potential |
| Kcnj9 | 1.38 | regulation of presynaptic membrane potential |
| L1cam | −0.17 | modulation of chemical synaptic transmission; regulation of synapse assembly |
| Map2k1 | 0.18 | regulation of neurotransmitter receptor localization to postsynaptic specialization membrane |
| Marcks | 0.41 | regulation of modification of postsynaptic actin cytoskeleton |
| Myo5a | −0.11 | establishment of endoplasmic reticulum localization to postsynapse; regulation of postsynaptic cytosolic calcium levels; transport |
| Nedd4 | −0.22 | regulation of postsynaptic neurotransmitter receptor endocytosis; regulation of protein catabolic process at postsynapse, modulating synaptic transmission; regulation protein catabolic process at postsynapse |
| Nptn | −0.18 | trans-synaptic signaling by trans-synaptic complex, modulating synaptic transmission |
| Ogt | 0.25 | regulation of neurotransmitter receptor localization to postsynaptic specialization membrane; regulation of synapse assembly |
| Pabpc1 | −0.09 | - |
| Parn | −0.30 | - |
| Ppfia3 | 0.49 | presynapse assembly; synaptic vesicle docking; synaptic vesicle exocytosis |
| Ppp2ca | −0.15 | - |
| Ppp2r1a | −0.11 | - |
| Ptprf | 0.21 | regulation of postsynapse organization; synapse adhesion between pre- and postsynapse; |
| Rab7 | −0.20 | neurotransmitter receptor transport, postsynaptic endosome to lysosome; synaptic vesicle recycling via endosome |
| Rheb | −0.16 | regulation of postsynapse organization |
| Rpl10a | 0.40 | translation at postsynapse: translation at presynapse |
| Rpl13 | 0.21 | translation at postsynapse; translation at presynapse |
| Rpl14 | −0.19 | translation at postsynapse; translation at presynapse |
| Rpl19 | 0.21 | - |
| Rpl21 | −0.06 | - |
| Rpl31 | 0.09 | - |
| Rpl4 | −0.17 | translation at postsynapse; translation at presynapse |
| Rpl5 | −0.08 | translation at postsynapse; translation at presynapse |
| Rpl7 | −0.31 | translation at postsynapse; translation at presynapse |
| Rps17 | −0.09 | - |
| Rps19 | 0.23 | - |
| Rps24 | −0.15 | translation at postsynapse; translation at presynapse |
| Rps27 | 0.20 | translation at postsynapse; translation at presynapse |
| Rps4x | −0.13 | - |
| Rps8 | 0.23 | - |
| Rps9 | 0.27 | - |
| Rtn4 | −0.15 | modulation of chemical synaptic transmission; regulation of postsynapse assembly |
| Scn8a | 0.23 | - |
| Sgip1 | 0.21 | synaptic vesicle endocytosis |
| Slc6a9 | 0.33 | neurotransmitter uptake |
| Slk | −0.20 | modulation of chemical synaptic transmission |
| Syngap1 | 0.22 | maintenance of postsynaptic specialization structure; modulation of chemical synaptic transmission |
| Trio | 0.12 | modulation of chemical synaptic transmission; postsynaptic modulation of chemical synaptic transmission |
| Vps16 | 0.35 | - |
| Wasf1 | 0.17 | dendritic transport of mitochondrion; modification of postsynaptic actin cytoskeleton |
| Ybx1 | −0.06 | - |
| Ywhag | 0.11 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Artimagnella, O.; Mazzon, E.; Salamone, S.; Pollastro, F.; Gugliandolo, A.; Chiricosta, L. Cannabinerol (CBNR) Influences Synaptic Genes Associated with Cytoskeleton and Ion Channels in NSC-34 Cell Line: A Transcriptomic Study. Biomedicines 2024, 12, 189. https://doi.org/10.3390/biomedicines12010189
Artimagnella O, Mazzon E, Salamone S, Pollastro F, Gugliandolo A, Chiricosta L. Cannabinerol (CBNR) Influences Synaptic Genes Associated with Cytoskeleton and Ion Channels in NSC-34 Cell Line: A Transcriptomic Study. Biomedicines. 2024; 12(1):189. https://doi.org/10.3390/biomedicines12010189
Chicago/Turabian StyleArtimagnella, Osvaldo, Emanuela Mazzon, Stefano Salamone, Federica Pollastro, Agnese Gugliandolo, and Luigi Chiricosta. 2024. "Cannabinerol (CBNR) Influences Synaptic Genes Associated with Cytoskeleton and Ion Channels in NSC-34 Cell Line: A Transcriptomic Study" Biomedicines 12, no. 1: 189. https://doi.org/10.3390/biomedicines12010189
APA StyleArtimagnella, O., Mazzon, E., Salamone, S., Pollastro, F., Gugliandolo, A., & Chiricosta, L. (2024). Cannabinerol (CBNR) Influences Synaptic Genes Associated with Cytoskeleton and Ion Channels in NSC-34 Cell Line: A Transcriptomic Study. Biomedicines, 12(1), 189. https://doi.org/10.3390/biomedicines12010189