

Genes Associated with Altered Brain Structure and Function in Obstructive Sleep Apnea
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
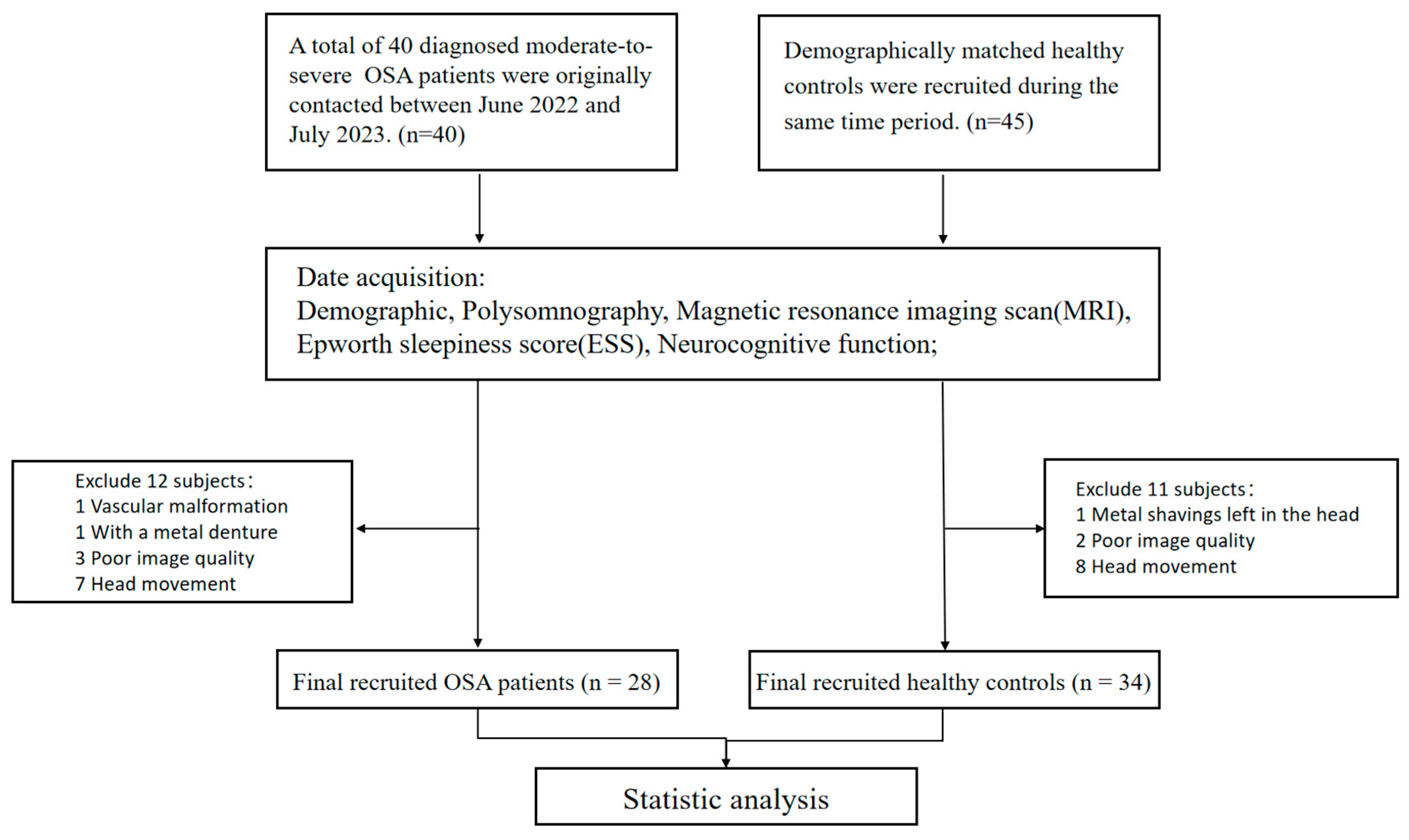
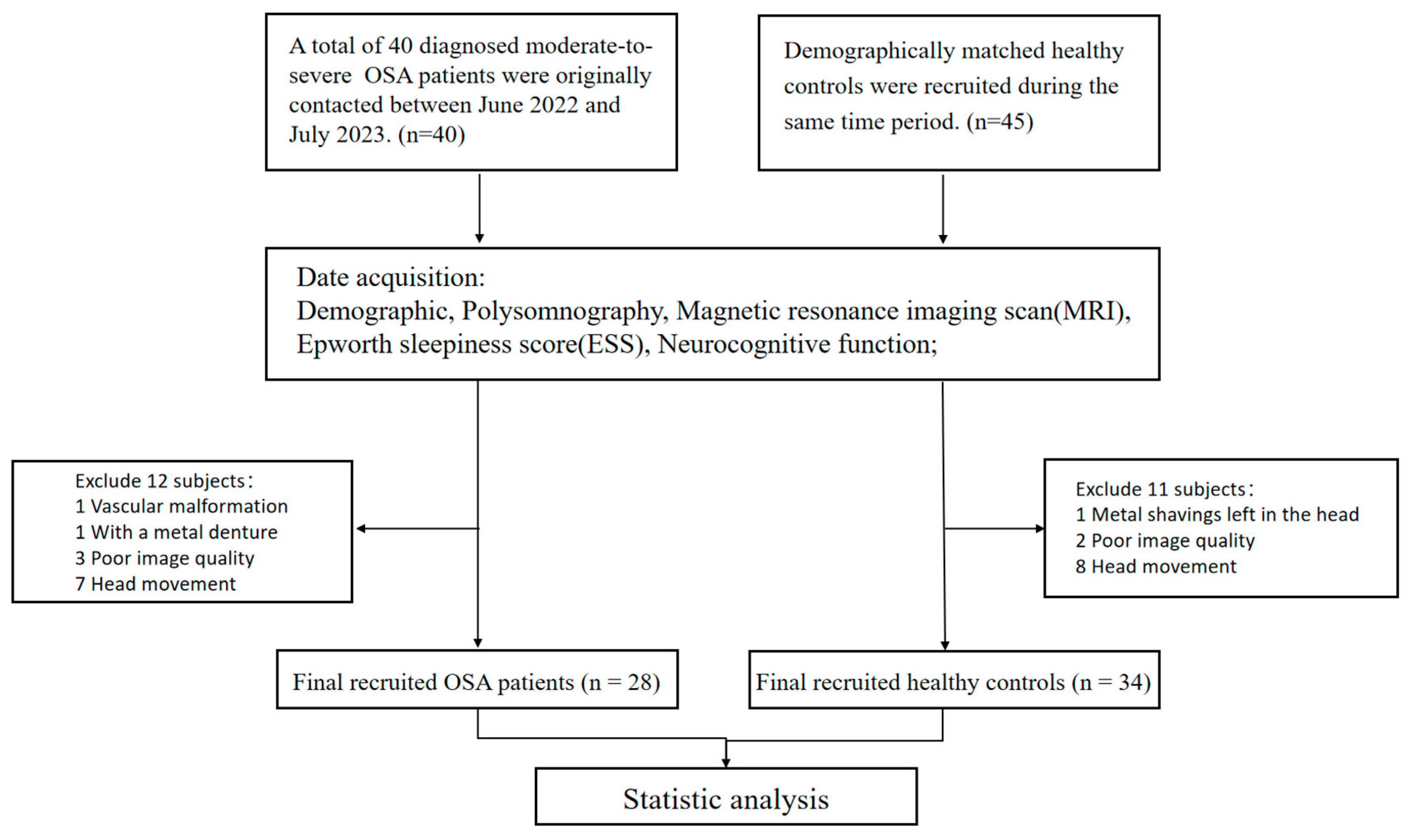
2.1. Participants
2.2. Polysomnography (PSG) and Questionnaires
2.3. Image Data Acquisition
2.4. Resting-State fMRI Preprocessing and ALFF Analysis
2.5. VBM Analysis
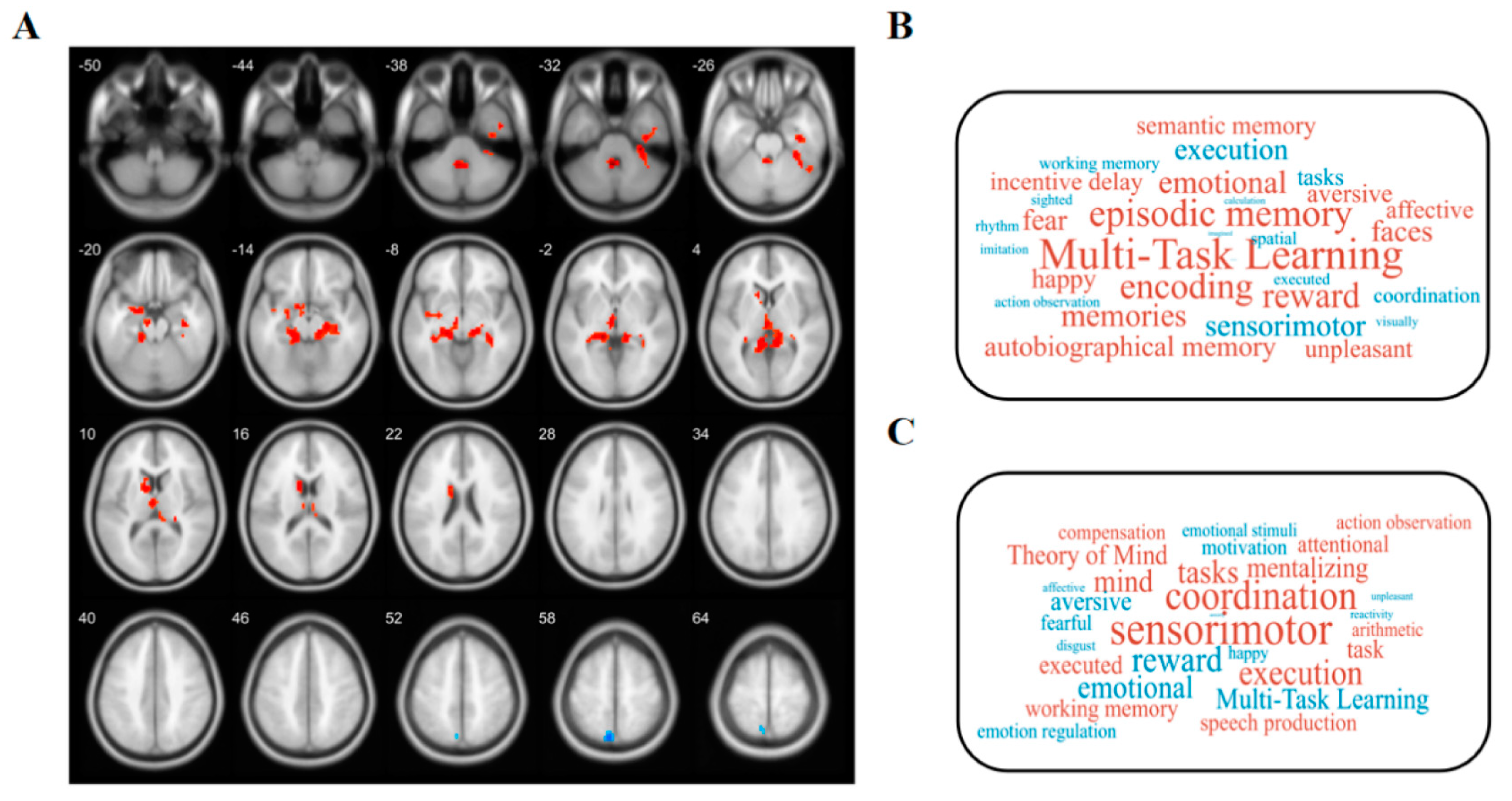
2.6. Meta-Analytical Decoding
2.7. Brain Gene Expression Data Processing
2.8. Gene Expression–Neuroimaging Spatial Correlation Analyses
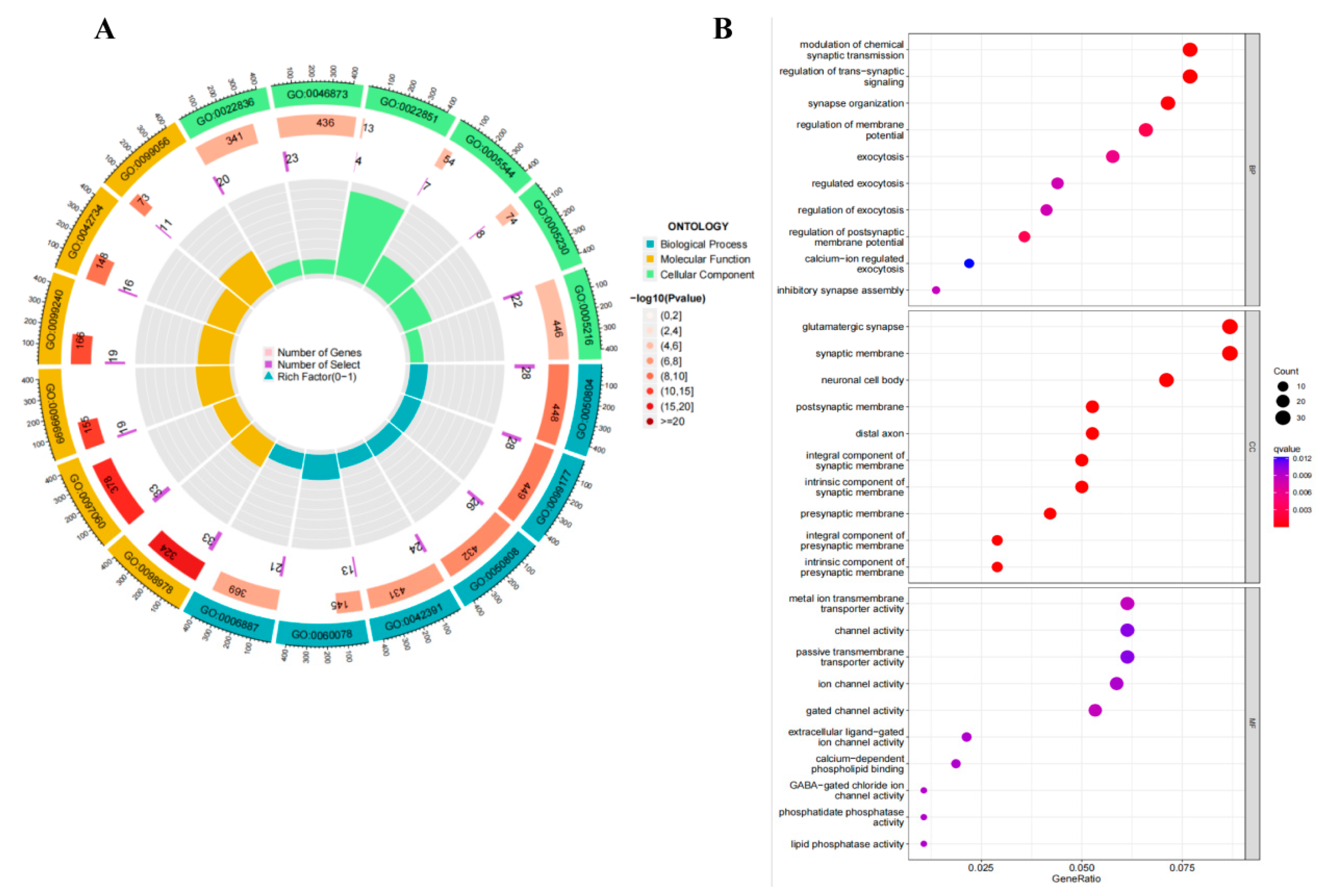
2.9. Pathway Analysis
2.10. Statistical Analysis
3. Results
3.1. Demographic and Clinical Characteristics
3.2. Altered ALFF
3.3. Changed GMV
3.4. Transcription–Neuroimaging Associations
3.5. Pathway Analysis
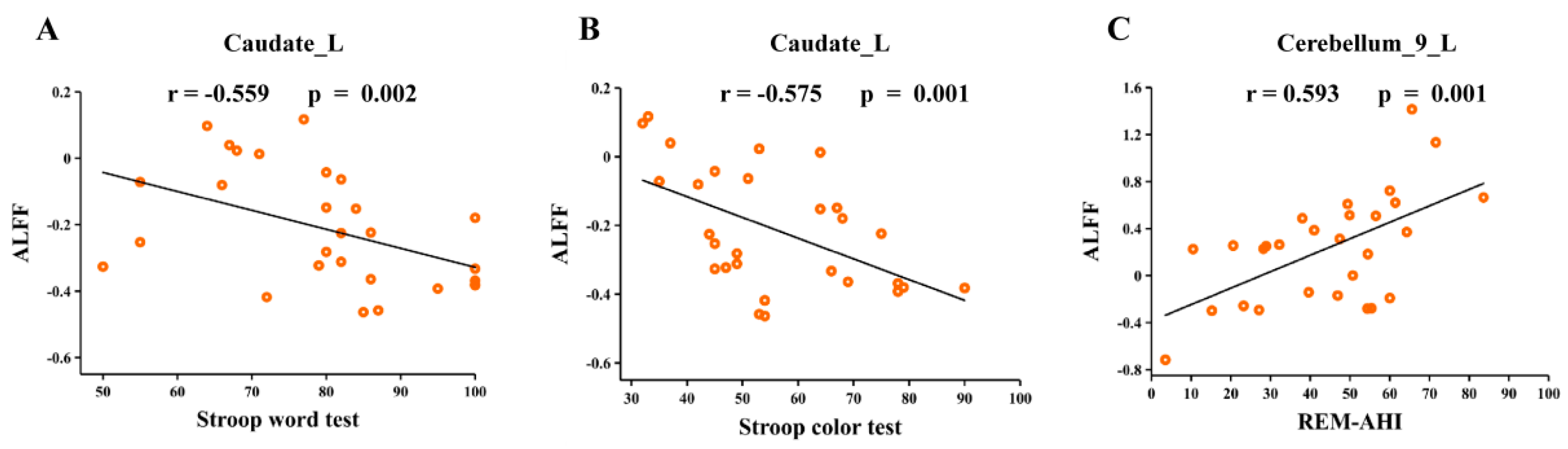
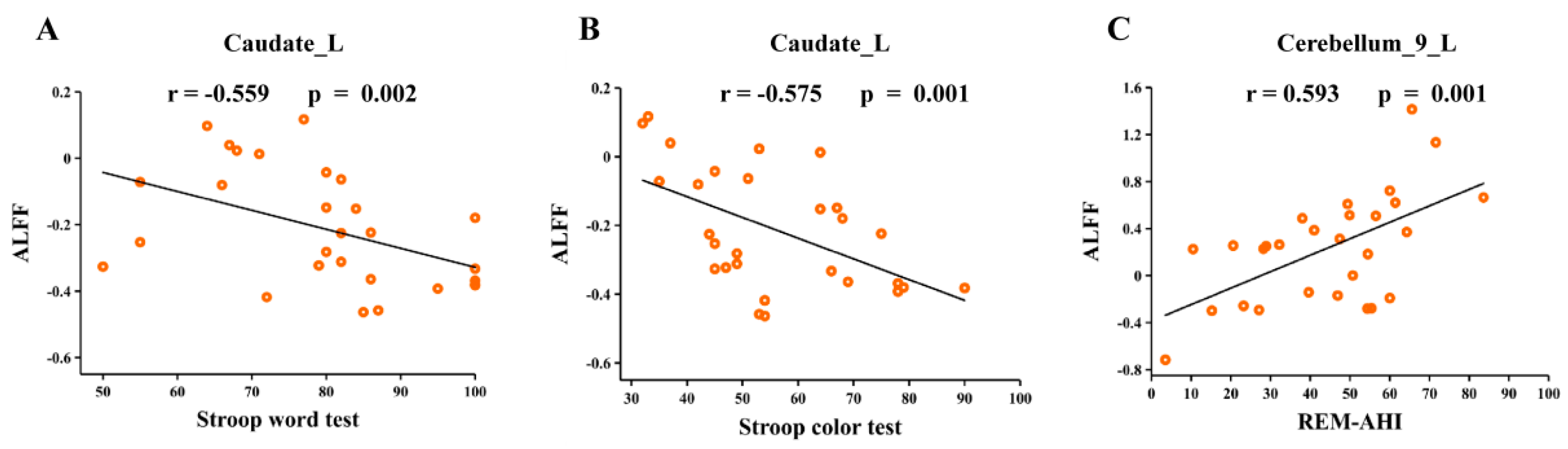
3.6. Correlation Analyses
4. Discussion
4.1. Brain Regions with Altered ALFF Values
4.2. Brain Regions with Altered GMV Values
4.3. Gene Enrichment Analysis
4.4. Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lv, R.; Liu, X.; Zhang, Y.; Dong, N.; Wang, X.; He, Y.; Yue, H.; Yin, Q. Pathophysiological mechanisms and therapeutic approaches in obstructive sleep apnea syndrome. Signal Transduct. Target. Ther. 2023, 8, 218. [Google Scholar] [CrossRef] [PubMed]
- Benjafield, A.V.; Ayas, N.T.; Eastwood, P.R.; Heinzer, R.; Ip, M.S.M.; Morrell, M.J.; Nunez, C.M.; Patel, S.R.; Penzel, T.; Pépin, J.L.; et al. Estimation of the global prevalence and burden of obstructive sleep apnoea: A literature-based analysis. Lancet Respir. Med. 2019, 7, 687–698. [Google Scholar] [CrossRef] [PubMed]
- Vanek, J.; Prasko, J.; Genzor, S.; Ociskova, M.; Kantor, K.; Holubova, M.; Slepecky, M.; Nesnidal, V.; Kolek, A.; Sova, M. Obstructive sleep apnea, depression and cognitive impairment. Sleep Med. 2020, 72, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Arnaud, C.; Dematteis, M.; Pepin, J.L.; Baguet, J.P.; Lévy, P. Obstructive sleep apnea, immuno-inflammation, and atherosclerosis. Semin Immunopathol. 2009, 31, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Ji, T.; Li, X.; Chen, J.; Ren, X.; Mei, L.; Qiu, Y.; Zhang, J.; Wang, S.; Xu, Z.; Li, H.; et al. Brain function in children with obstructive sleep apnea: A resting-state fMRI study. Sleep 2021, 44, zsab047. [Google Scholar] [CrossRef] [PubMed]
- Taylor, K.S.; Millar, P.J.; Murai, H.; Haruki, N.; Kimmerly, D.S.; Bradley, T.D.; Floras, J.S. Cortical autonomic network gray matter and sympathetic nerve activity in obstructive sleep apnea. Sleep 2018, 41, zsx208. [Google Scholar] [CrossRef] [PubMed]
- Calhoun, V. Data-driven approaches for identifying links between brain structure and function in health and disease. Dialogues Clin. Neurosci. 2018, 20, 87–99. [Google Scholar] [CrossRef]
- Li, J.; Lv, Q.; Sun, H.; Yang, Y.; Jiao, X.; Yang, S.; Yu, H.; Qin, Y. Combined Association between ADIPOQ, PPARG, and TNF Genes Variants and Obstructive Sleep Apnea in Chinese Han Population. Nat. Sci. Sleep 2022, 14, 363–372. [Google Scholar] [CrossRef]
- Xu, H.; Liu, F.; Li, Z.; Li, X.; Liu, Y.; Li, N.; Zhang, X.; Gao, Z.; Zhang, X.; Liu, Y.; et al. Genome-Wide Association Study of Obstructive Sleep Apnea and Objective Sleep-related Traits Identifies Novel Risk Loci in Han Chinese Individuals. Am. J. Respir. Crit. Care Med. 2022, 206, 1534–1545. [Google Scholar] [CrossRef]
- Romero-Garcia, R.; Warrier, V.; Bullmore, E.T.; Baron-Cohen, S.; Bethlehem, R.A.I. Synaptic and transcriptionally downregulated genes are associated with cortical thickness differences in autism. Mol. Psychiatry 2019, 24, 1053–1064. [Google Scholar] [CrossRef]
- Xu, X.; Li, Q.; Qian, Y.; Cai, H.; Zhang, C.; Zhao, W.; Zhu, J.; Yu, Y. Genetic mechanisms underlying gray matter volume changes in patients with drug-naive first-episode schizophrenia. Cereb. Cortex 2023, 33, 2328–2341. [Google Scholar] [CrossRef] [PubMed]
- Kapur, V.K.; Auckley, D.H.; Chowdhuri, S.; Kuhlmann, D.C.; Mehra, R.; Ramar, K.; Harrod, C.G. Clinical Practice Guideline for Diagnostic Testing for Adult Obstructive Sleep Apnea: An American Academy of Sleep Medicine Clinical Practice Guideline. J. Clin. Sleep Med. 2017, 13, 479–504. [Google Scholar] [CrossRef] [PubMed]
- Hawrylycz, M.J.; Lein, E.S.; Guillozet-Bongaarts, A.L.; Shen, E.H.; Ng, L.; Miller, J.A.; van de Lagemaat, L.N.; Smith, K.A.; Ebbert, A.; Riley, Z.L.; et al. An anatomically comprehensive atlas of the adult human brain transcriptome. Nature 2012, 489, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Markello, R.D.; Arnatkeviciute, A.; Poline, J.B.; Fulcher, B.D.; Fornito, A.; Misic, B. Standardizing workflows in imaging transcriptomics with the abagen toolbox. Elife 2021, 10, e72129. [Google Scholar] [CrossRef] [PubMed]
- Xue, K.; Guo, L.; Zhu, W.; Liang, S.; Xu, Q.; Ma, L.; Liu, M.; Zhang, Y.; Liu, F. Transcriptional signatures of the cortical morphometric similarity network gradient in first-episode, treatment-naive major depressive disorder. Neuropsychopharmacology 2023, 48, 518–528. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. ClusterProfiler: An R package for comparing biological themes among gene clusters. Omics 2012, 16, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Lucke-Wold, B.P.; Smith, K.E.; Nguyen, L.; Turner, R.C.; Logsdon, A.F.; Jackson, G.J.; Huber, J.D.; Rosen, C.L.; Miller, D.B. Sleep disruption and the sequelae associated with traumatic brain injury. Neurosci. Biobehav. Rev. 2015, 55, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Turner, R.C.; Lucke-Wold, B.; Lucke-Wold, N.; Elliott, A.S.; Logsdon, A.F.; Rosen, C.L.; Huber, J.D. Neuroprotection for ischemic stroke: Moving past shortcomings and identifying promising directions. Int. J. Mol. Sci. 2013, 14, 1890–1917. [Google Scholar] [CrossRef]
- Fang, H.; Lu, C.; Hong, F.; Jiang, W.; Wang, T. Sleep Apnea Detection Based on Multi-Scale Residual Network. Life 2022, 12, 119. [Google Scholar] [CrossRef]
- Gozal, D.; Row, B.W.; Kheirandish, L.; Liu, R.; Guo, S.Z.; Qiang, F.; Brittian, K.R. Increased susceptibility to intermittent hypoxia in aging rats: Changes in proteasomal activity, neuronal apoptosis and spatial function. J. Neurochem. 2003, 86, 1545–1552. [Google Scholar] [CrossRef]
- Cavanna, A.E.; Trimble, M.R. The precuneus: A review of its functional anatomy and behavioural correlates. Brain 2006, 129 Pt 3, 564–583. [Google Scholar] [CrossRef] [PubMed]
- Ylä-Herttuala, S.; Hakulinen, M.; Poutiainen, P.; Laitinen, T.M.; Koivisto, A.M.; Remes, A.M.; Hallikainen, M.; Lehtola, J.M.; Saari, T.; Korhonen, V.; et al. Severe Obstructive Sleep Apnea and Increased Cortical Amyloid-β Deposition. J. Alzheimers Dis. 2021, 79, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Gozal, E.; Row, B.W.; Schurr, A.; Gozal, D. Developmental differences in cortical and hippocampal vulnerability to intermittent hypoxia in the rat. Neurosci. Lett. 2001, 305, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Wiltgen, B.J.; Zhou, M.; Cai, Y.; Balaji, J.; Karlsson, M.G.; Parivash, S.N.; Li, W.; Silva, A.J. The hippocampus plays a selective role in the retrieval of detailed contextual memories. Curr. Biol. 2010, 20, 1336–1344. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.H.; Sin, S.; Lee, S.; Wagshul, M.E.; Zimmerman, M.E.; Arens, R. Cortical thickness and hippocampal volume in adolescent children with obstructive sleep apnea. Sleep 2023, 46, zsac201. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Roy, B.; Kang, D.W.; Aysola, R.S.; Macey, P.M.; Woo, M.A.; Yan-Go, F.L.; Harper, R.M.; Kumar, R. Altered resting-state hippocampal and caudate functional networks in patients with obstructive sleep apnea. Brain Behav. 2018, 8, e00994. [Google Scholar] [CrossRef] [PubMed]
- Ochsner, K.N.; Ray, R.R.; Hughes, B.; McRae, K.; Cooper, J.C.; Weber, J.; Gabrieli, J.D.; Gross, J.J. Bottom-up and top-down processes in emotion generation: Common and distinct neural mechanisms. Psychol. Sci. 2009, 20, 1322–1331. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Chen, L.; Li, H.; Xin, H.; Zhang, J.; Wei, Z.; Peng, D. Abnormal resting-state functional connectivity of amygdala subregions in patients with obstructive sleep apnea. Neuropsychiatr. Dis. Treat. 2019, 15, 977–987. [Google Scholar] [CrossRef]
- Tahmasian, M.; Rosenzweig, I.; Eickhoff, S.B.; Sepehry, A.A.; Laird, A.R.; Fox, P.T.; Morrell, M.J.; Khazaie, H.; Eickhoff, C.R. Structural and functional neural adaptations in obstructive sleep apnea: An activation likelihood estimation meta-analysis. Neurosci. Biobehav. Rev. 2016, 65, 142–156. [Google Scholar] [CrossRef]
- Luppi, P.H.; Billwiller, F.; Fort, P. Selective activation of a few limbic structures during paradoxical (REM) sleep by the claustrum and the supramammillary nucleus: Evidence and function. Curr. Opin. Neurobiol. 2017, 44, 59–64. [Google Scholar] [CrossRef]
- Zhang, A.; Wang, X.; Li, J.; Jing, L.; Hu, X.; Li, H.; Yang, C.; Zhang, K.; Sun, N. Resting-State fMRI in Predicting Response to Treatment With SSRIs in First-Episode, Drug-Naive Patients With Major Depressive Disorder. Front. Neurosci. 2022, 16, 831278. [Google Scholar] [CrossRef] [PubMed]
- Khalifa, M.K.A.; Salem, H.A.; Shawky, S.M.; Eassa, H.A.; Elaidy, A.M. Enhancement of zaleplon oral bioavailability using optimized self-nano emulsifying drug delivery systems and its effect on sleep quality among a sample of psychiatric patients. Drug Deliv. 2019, 26, 1243–1253. [Google Scholar] [CrossRef] [PubMed]
- Crane, N.A.; Jenkins, L.M.; Dion, C.; Meyers, K.K.; Weldon, A.L.; Gabriel, L.B.; Walker, S.J.; Hsu, D.T.; Noll, D.C.; Klumpp, H.; et al. Comorbid anxiety increases cognitive control activation in Major Depressive Disorder. Depress. Anxiety 2016, 33, 967–977. [Google Scholar] [CrossRef] [PubMed]
- Scarpina, F.; Tagini, S. The Stroop Color and Word Test. Front. Psychol. 2017, 8, 557. [Google Scholar] [CrossRef] [PubMed]
- Canto, C.B.; Onuki, Y.; Bruinsma, B.; van der Werf, Y.D.; De Zeeuw, C.I. The Sleeping Cerebellum. Trends Neurosci. 2017, 40, 309–323. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, J.; Sun, H.; Yang, J.; Ma, Y.; Chen, K.; Su, J.; Yu, X.; Yang, F.; Zhang, Z.; et al. Cerebellum drives functional dysfunctions in restless leg syndrome. Sleep Med. 2023, 110, 172–178. [Google Scholar] [CrossRef]
- Shechter, A.; Foster, G.D.; Lang, W.; Reboussin, D.M.; St-Onge, M.P.; Zammit, G.; Newman, A.B.; Millman, R.P.; Wadden, T.A.; Jakicic, J.M.; et al. Effects of a lifestyle intervention on REM sleep-related OSA severity in obese individuals with type 2 diabetes. J. Sleep Res. 2017, 26, 747–755. [Google Scholar] [CrossRef]
- Numssen, O.; Bzdok, D.; Hartwigsen, G. Functional specialization within the inferior parietal lobes across cognitive domains. Elife 2021, 10, e63591. [Google Scholar] [CrossRef]
- Krause, A.J.; Simon, E.B.; Mander, B.A.; Greer, S.M.; Saletin, J.M.; Goldstein-Piekarski, A.N.; Walker, M.P. The sleep-deprived human brain. Nat. Rev. Neurosci. 2017, 18, 404–418. [Google Scholar] [CrossRef]
- Zhong, M.; Zeng, H.; Wang, D.; Li, J.; Duan, X.; Li, Y. Structure and activity alteration in adult highland residents’ cerebrum: Voxel-based morphometry and amplitude of low-frequency fluctuation study. Front. Neurosci. 2022, 16, 1035308. [Google Scholar] [CrossRef]
- Holt, W.V.; Gadea, J. Information Flow between Parents and Offspring Is Essential for Successful Development. Biomolecules 2023, 13, 1299. [Google Scholar] [CrossRef] [PubMed]
- Ko, G.Y.; Shi, L.; Ko, M.L. Circadian regulation of ion channels and their functions. J. Neurochem. 2009, 110, 1150–1169. [Google Scholar] [CrossRef] [PubMed]
- Forcaia, G.; Formicola, B.; Terribile, G.; Negri, S.; Lim, D.; Biella, G.; Re, F.; Moccia, F.; Sancini, G. Multifunctional Liposomes Modulate Purinergic Receptor-Induced Calcium Wave in Cerebral Microvascular Endothelial Cells and Astrocytes: New Insights for Alzheimer’s disease. Mol. Neurobiol. 2021, 43, 2824–2835. [Google Scholar] [CrossRef] [PubMed]
- McMoneagle, E.; Zhou, J.; Zhang, S.; Huang, W.; Josiah, S.S.; Ding, K.; Wang, Y.; Zhang, J. Neuronal K(+)-Cl(-) cotransporter KCC2 as a promising drug target for epilepsy treatment. Acta Pharmacol. Sin. 2023. [Google Scholar] [CrossRef] [PubMed]
- Rubin, B.R.; Milner, T.A.; Pickel, V.M.; Coleman, C.G.; Marques-Lopes, J.; Van Kempen, T.A.; Kazim, S.F.; McEwen, B.S.; Gray, J.D.; Pereira, A.C. Sex and age differentially affect GABAergic neurons in the mouse prefrontal cortex and hippocampus following chronic intermittent hypoxia. Exp. Neurol. 2020, 325, 113075. [Google Scholar] [CrossRef] [PubMed]
- Mondino, A.; Hambrecht-Wiedbusch, V.S.; Li, D.; York, A.K.; Pal, D.; González, J.; Torterolo, P.; Mashour, G.A.; Vanini, G. Glutamatergic Neurons in the Preoptic Hypothalamus Promote Wakefulness, Destabilize NREM Sleep, Suppress REM Sleep, and Regulate Cortical Dynamics. J. Neurosci. 2021, 41, 3462–3478. [Google Scholar] [CrossRef] [PubMed]
- de Bartolomeis, A.; Barone, A.; Buonaguro, E.F.; Tomasetti, C.; Vellucci, L.; Iasevoli, F. The Homer1 family of proteins at the crossroad of dopamine-glutamate signaling: An emerging molecular “Lego” in the pathophysiology of psychiatric disorders. A systematic review and translational insight. Neurosci. Biobehav. Rev. 2022, 136, 104596. [Google Scholar] [CrossRef] [PubMed]
- Korenic, S.A.; Klingaman, E.A.; Wickwire, E.M.; Gaston, F.E.; Chen, H.; Wijtenburg, S.A.; Rowland, L.M. Sleep quality is related to brain glutamate and symptom severity in schizophrenia. J. Psychiatr. Res. 2020, 120, 14–20. [Google Scholar] [CrossRef]
- Lodato, S.; Shetty, A.S.; Arlotta, P. Cerebral cortex assembly: Generating and reprogramming projection neuron diversity. Trends Neurosci. 2015, 38, 117–125. [Google Scholar] [CrossRef]
- Terra, H.; Bruinsma, B.; de Kloet, S.F.; van der Roest, M.; Pattij, T.; Mansvelder, H.D. Prefrontal Cortical Projection Neurons Targeting Dorsomedial Striatum Control Behavioral Inhibition. Curr. Biol. 2020, 30, 4188–4200.e4185. [Google Scholar] [CrossRef]
- Lyons, L.C.; Vanrobaeys, Y.; Abel, T. Sleep and memory: The impact of sleep deprivation on transcription, translational control, and protein synthesis in the brain. J. Neurochem. 2023, 166, 24–46. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Reiner, B.; Liu, J.; Xu, L.; Xiong, H. Methamphetamine augments HIV-1 gp120 inhibition of synaptic transmission and plasticity in rat hippocampal slices: Implications for methamphetamine exacerbation of HIV-associated neurocognitive disorders. Neurobiol. Dis. 2022, 168, 105712. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Demographic | OSA (n = 28) | HCs (n = 34) | p Value |
|---|---|---|---|
| Age (year) | 48 (36–53) | 41 (33–51) | 0.731 |
| Gender (male) (%) | 25 (89.3) | 26 (76.5) | 0.317 |
| BMI (kg/m2) | 27.9 (26.7–30.5) | 24.2 (22.3–26.4) | <0.001 |
| Education (year) | 15 (12–16) | 15 (12–16) | 0.344 |
| Smoker (%) | 19 (67.9) | 17 (50.0) | 0.200 |
| ESS scores | 10.0 (7.0–16.0) | 3.0 (2.0–4.5) | <0.001 |
| Sleep efficiency (%) | 84.6 (75.5–89.8) | 89.5 (86.2–91.3) | 0.004 |
| N3 period (%) | 6.7 (1.2–13.3) | 18.6 (12.4–20.4) | <0.001 |
| REM period (%) | 18.4 (14.8–21.8) | 20.3 (17.0–24.9) | 0.157 |
| AHI (events/hour) | 57.0 (31.8–76.4) | 3.9 (3.3–4.6) | <0.001 |
| REM-AHI (events/hour) | 47.2 (28.4–59.1) | 2.7 (2.0–3.5) | <0.001 |
| NREM-AHI (events/hour) | 53.6 (29.9–71.8) | 3.8 (3.1–4.7) | <0.001 |
| LSpO2 (%) | 75.5 (58.8–85) | 92.0 (91.5–95.2) | <0.001 |
| MSpO2 (%) | 94.0 (92.0–96.0) | 96.0 (95.5–97.0) | <0.001 |
| ODI (events/hour) | 45.4 (24.1–70.6) | 3.1 (1.8–4.5) | <0.001 |
| Arousal Index (events/hour) | 45.3 (27.7–71.3) | 10.8 (6.7–15.5) | <0.001 |
| MoCA | 22.5 (20.0–26.0) | 25.0 (24.0–27.0) | 0.003 |
| Trial Making Test-A | 37.0 (29.3–47.8) | 34.0 (29.0–41.5) | 0.442 |
| Symbol Coding | 39.0 (27.0–47.8) | 38.0 (36.0–43.5) | 0.723 |
| HVLT-R | 16.0 (14.3–22.0) | 21.0 (18.5–24.5) | 0.004 |
| Spatial Span | 16.0 (15.0–19.0) | 19.0 (17.5–22.0) | 0.002 |
| Mazes | 12.5 (10.0–18.0) | 19.0 (17.5–22.0) | <0.001 |
| BVMT-R | 16.0 (12.0–22.0) | 24.0 (21.0–28.0) | 0.002 |
| Category Fluency | 17.0 (15.0–21.5) | 22.0 (20.0–26.0) | 0.001 |
| CPT-IP | 2.2 (1.4–2.6) | 2.7 (2.3–3.1) | 0.031 |
| Stroop word test | 81.0 (69.5–88.5) | 92.0 (86.0–100.0) | 0.001 |
| Stroop color test | 53.0 (45.0–68.8) | 75.0 (69.0–80.0) | <0.001 |
| SCWT | 32.5 (25.5–37.8) | 39.0 (32.5–44.0) | 0.005 |
| Indices | Brain Regions | L/R | Peak MNI Coordinates | Cluster Size | t-Value | ||
|---|---|---|---|---|---|---|---|
| X | Y | Z | |||||
| ALFF | Hippocampus | L | −15 | −33 | −3 | 41 | 5.294 |
| Amygdala | L | −21 | −3 | −18 | 25 | 4.176 | |
| Caudate | L | −12 | 6 | 12 | 67 | 4.659 | |
| Precuneus | L | −6 | −72 | 57 | 50 | −4.776 | |
| Cerebellum_9 | L | −3 | −48 | −33 | 25 | 5.896 | |
| GMV | Parietal_Inf | R | 44 | −59 | 60 | 106 | 5.158 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, Y.; Shen, C.; Zhao, W.; Shang, Y.; Wang, Y.; Zhang, H.-T.; Ouyang, R.; Liu, J. Genes Associated with Altered Brain Structure and Function in Obstructive Sleep Apnea. Biomedicines 2024, 12, 15. https://doi.org/10.3390/biomedicines12010015
Huang Y, Shen C, Zhao W, Shang Y, Wang Y, Zhang H-T, Ouyang R, Liu J. Genes Associated with Altered Brain Structure and Function in Obstructive Sleep Apnea. Biomedicines. 2024; 12(1):15. https://doi.org/10.3390/biomedicines12010015
Chicago/Turabian StyleHuang, Yijie, Chong Shen, Wei Zhao, Youlan Shang, Yisong Wang, Hui-Ting Zhang, Ruoyun Ouyang, and Jun Liu. 2024. "Genes Associated with Altered Brain Structure and Function in Obstructive Sleep Apnea" Biomedicines 12, no. 1: 15. https://doi.org/10.3390/biomedicines12010015
APA StyleHuang, Y., Shen, C., Zhao, W., Shang, Y., Wang, Y., Zhang, H.-T., Ouyang, R., & Liu, J. (2024). Genes Associated with Altered Brain Structure and Function in Obstructive Sleep Apnea. Biomedicines, 12(1), 15. https://doi.org/10.3390/biomedicines12010015