
Benefits of Chlorella vulgaris against Cadmium Chloride-Induced Hepatic and Renal Toxicities via Restoring the Cellular Redox Homeostasis and Modulating Nrf2 and NF-KB Pathways in Male Rats


 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Husbandry
2.2. Tested Compounds
2.3. Experimental Design
2.4. Blood Collecting and Tissue Preservation
2.5. Hematological Analysis
2.6. Biochemical Analysis
2.7. Inflammatory Markers Evaluation in the Serum
2.8. Oxidative Stress Markers Detection in the Serum
2.9. Determination of Stress-Related Hormones
2.10. Separation of mRNA and Quantitative Real-Time PCR (qRT-PCR)
2.11. Histopathological Investigation
2.12. Statistical Analysis
3. Results
3.1. Chlorella vulgaris Improved the Hematological Parameters Altered by Cadmium
3.2. C. vulgaris Restores the Body and Organs Weight Affected by Cd Treatment
3.3. C. vulgaris Modulates the Serum Biochemical Parameters Altered by Cd Treatment
3.4. Cd Administration Induces the Whole Body’s Oxidative Stress
3.5. Cd Administration Elevates the Innate Immunity Response
3.6. Cd Administration Stimulates the Pro-Inflammatory Mediators
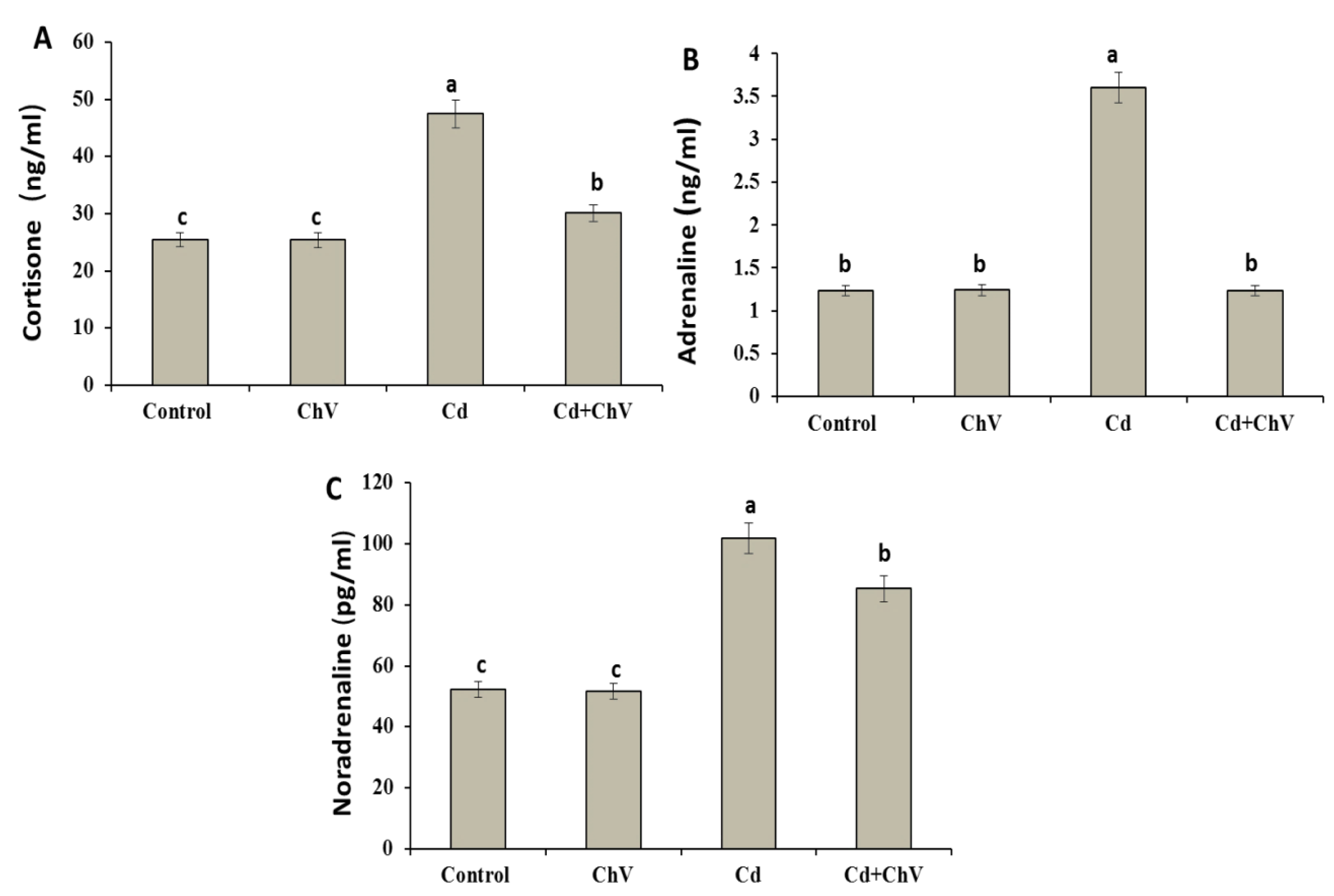
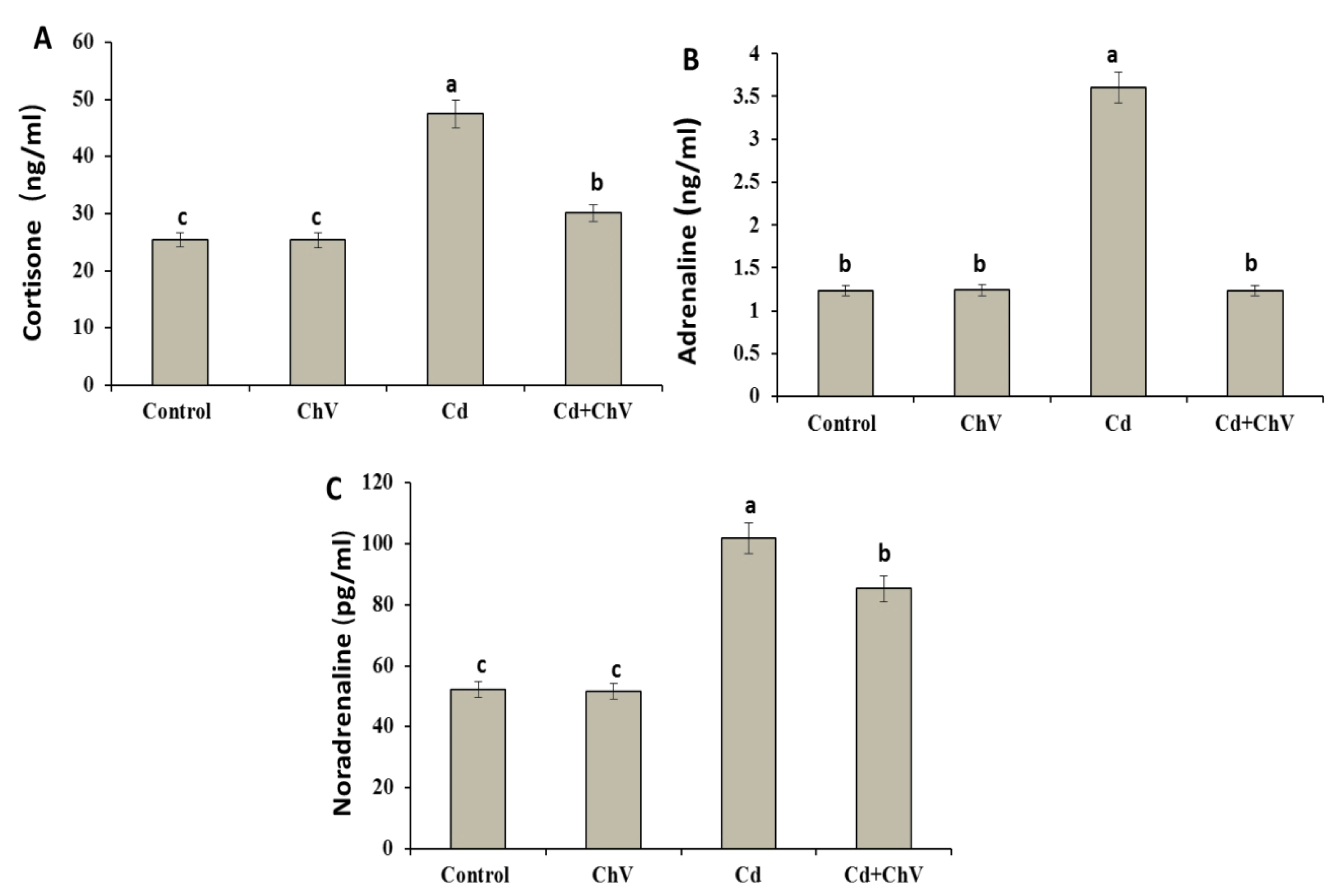
3.7. C. vulgaris Additive Modulates the Adrenal Hormones Induced by Cd Administration
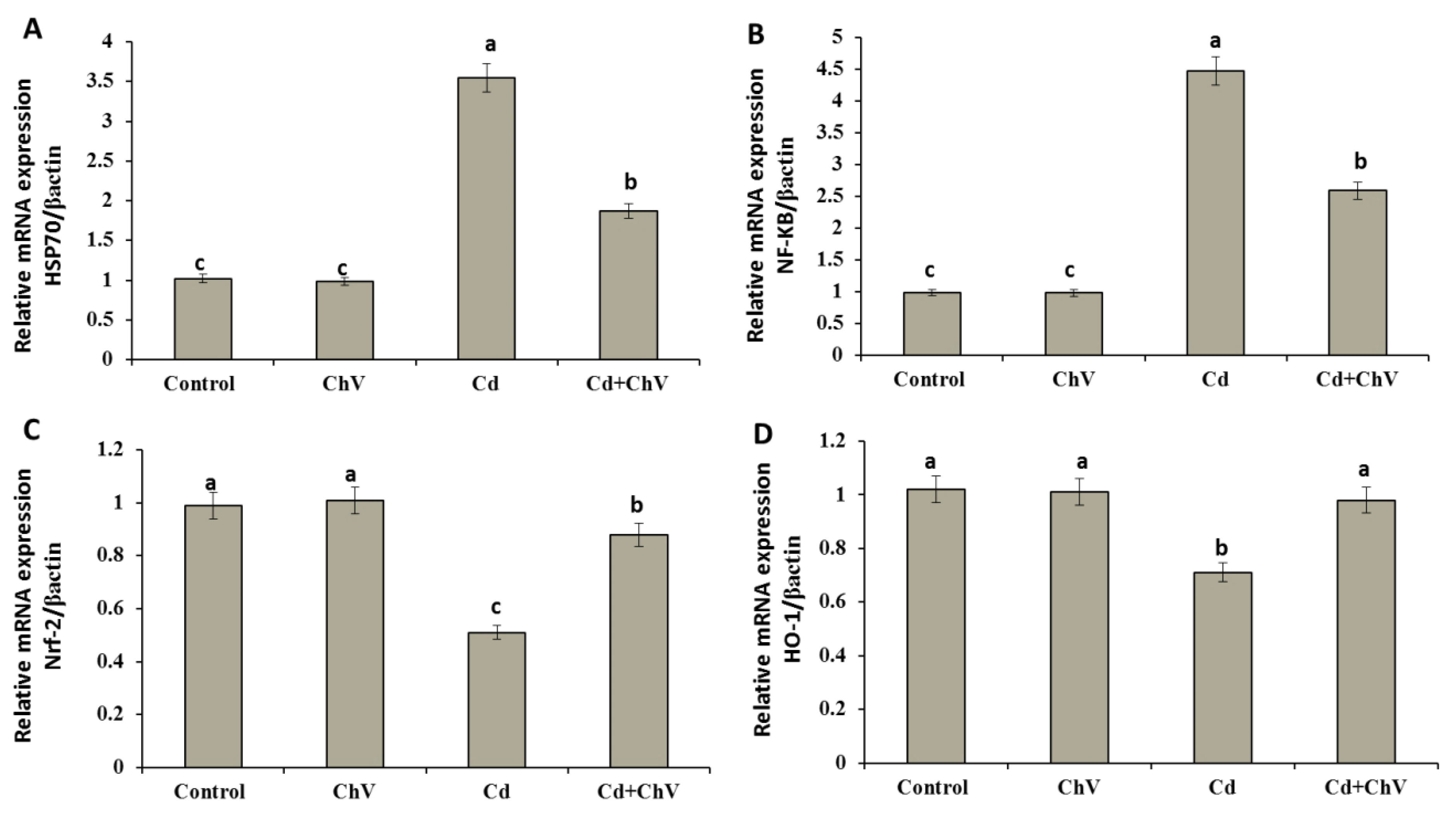
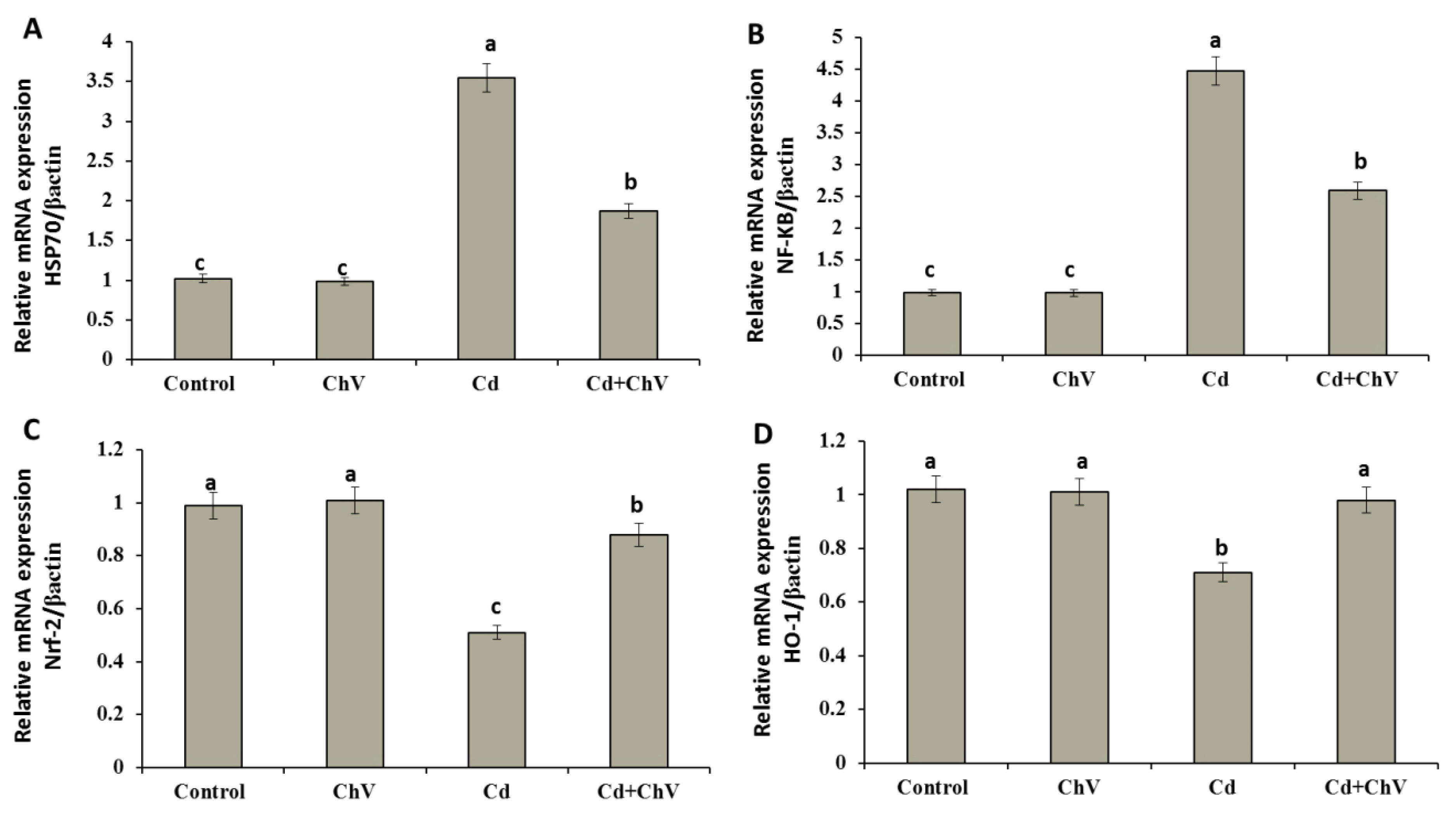
3.8. Chlorella vulgaris Restores the Cellular Redox Homeostasis by Modulating Stress Key Mediators at the Genetic Level
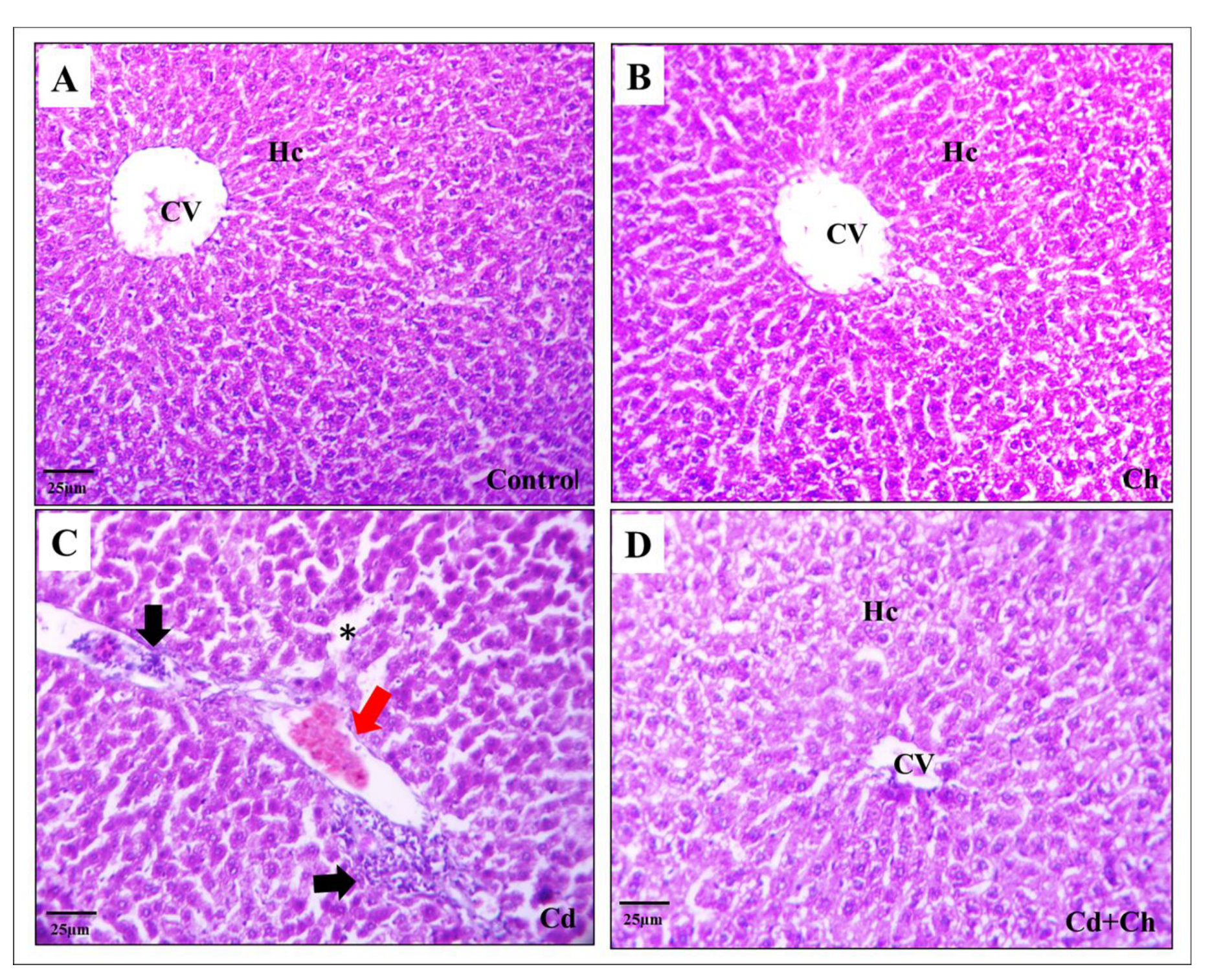
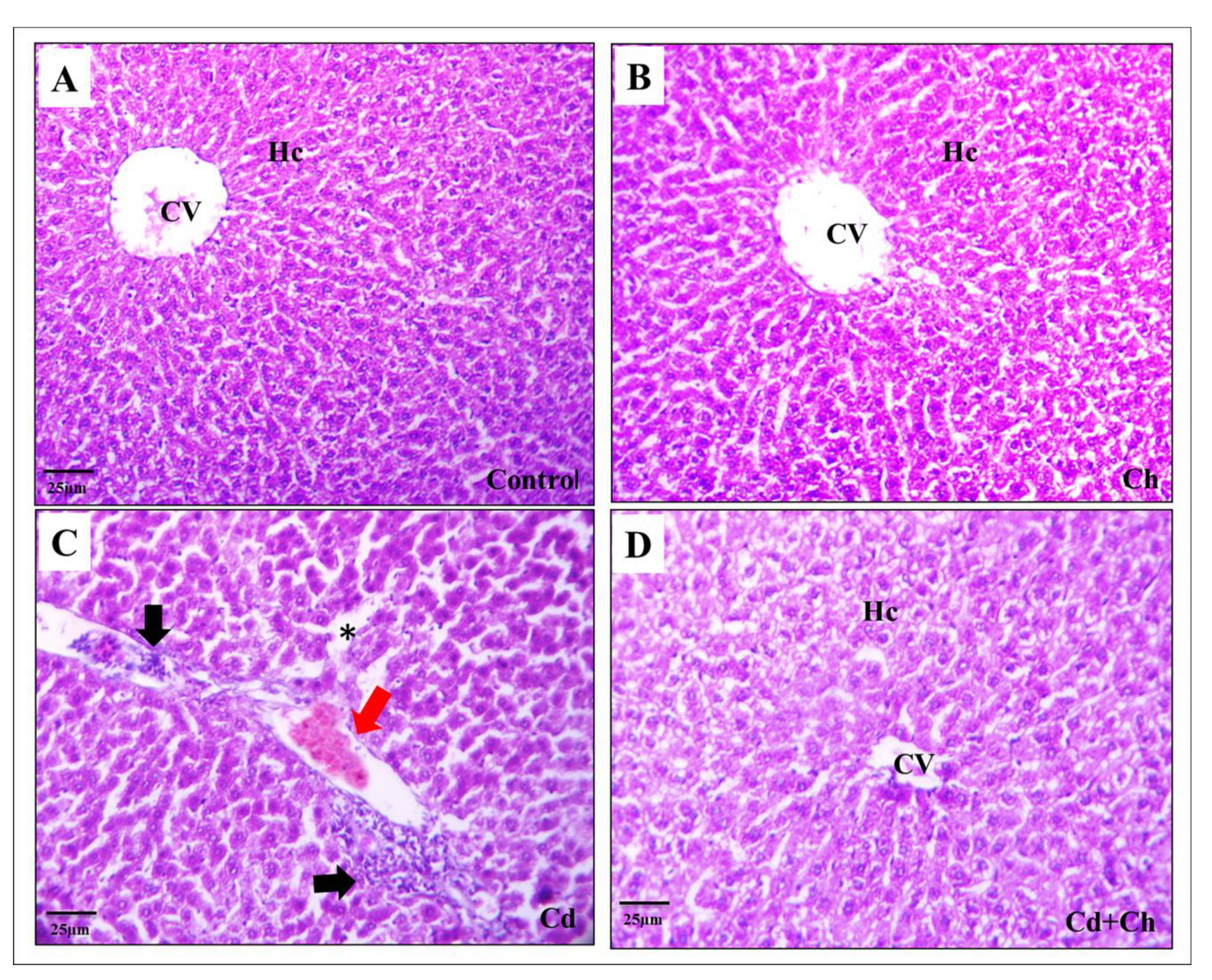
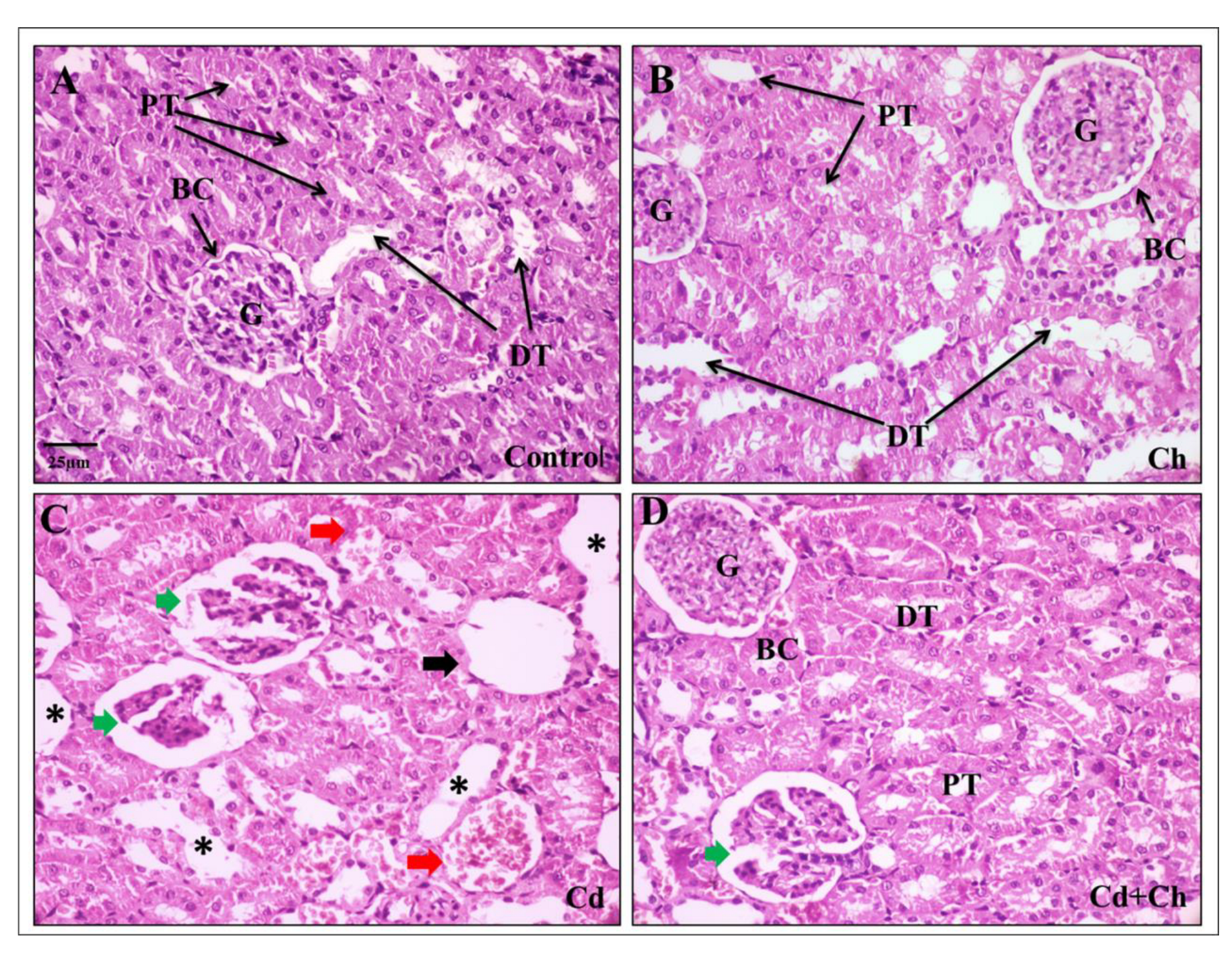
3.9. C. vulgaris Relief the Toxicity of CADMIUM in the Hepatic and Renal Tissues at the Histological Levels
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pandey, G.; Madhuri, S. Heavy metals causing toxicity in animals and fishes. Res. J. Anim. Vet. Fish. Sci. 2014, 2, 17–23. [Google Scholar]
- Järup, L.; Åkesson, A. Current status of cadmium as an environmental health problem. Toxicol. Appl. Pharmacol. 2009, 238, 201–208. [Google Scholar] [CrossRef]
- Gaurav, D.; Preet, S.; Dua, K. Protective effect of Spirulina platensis on cadmium induced renal toxicity in wistar rats. Arch. Appl. Sci. Res. 2010, 2, 390–397. [Google Scholar]
- Bernard, A. Renal dysfunction induced by cadmium: Biomarkers of critical effects. Biometals 2004, 17, 519–523. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Gao, X.; Luo, X.; Li, L.; Ma, M.; Zhu, Y.; Zhao, L.; Li, R. The effects of long-term exposure to low doses of cadmium on the health of the next generation of mice. Chem.-Biol. Interact. 2019, 312, 108792. [Google Scholar] [CrossRef] [PubMed]
- Lane, E.; Canty, M.J.; More, S.J. Cadmium exposure and consequence for the health and productivity of farmed ruminants. Res. Vet. Sci. 2015, 101, 132–139. [Google Scholar] [CrossRef]
- Souza, V.; Bucio, L.; Gutierrez-Ruiz, M. Cadmium uptake by a human hepatic cell line (WRL-68 cells). Toxicology 1997, 120, 215–220. [Google Scholar] [CrossRef]
- Koyu, A.; Gokcimen, A.; Ozguner, F.; Bayram, D.S.; Kocak, A. Evaluation of the effects of cadmium on rat liver. Mol. Cell. Biochem. 2006, 284, 81–85. [Google Scholar] [CrossRef]
- Kang, M.-Y.; Cho, S.-H.; Lim, Y.-H.; Seo, J.-C.; Hong, Y.-C. Effects of environmental cadmium exposure on liver function in adults. Occup. Environ. Med. 2013, 70, 268–273. [Google Scholar] [CrossRef]
- Elazab, S.T.; Hsu, W.H. Antagonism of cadmium-induced liver injury in ducks by α-bisabolol. Front. Vet. Sci. 2022, 9, 1024549. [Google Scholar] [CrossRef]
- Al-Nasser, I.A.; Al-Nasser, I. Cadmium hepatotoxicity and alterations of the mitochondrial function. J. Toxicol. Clin. Toxicol. 2000, 38, 407–413. [Google Scholar] [CrossRef] [PubMed]
- Berzina, N.; Markovs, J.; Isajevs, S.; Apsite, M.; Smirnova, G. Cadmium-Induced Enteropathy in Domestic Cocks: A Biochemical and Histological Study after Subchronic Exposure. Basic Clin. Pharmacol. Toxicol. 2007, 101, 29–34. [Google Scholar] [CrossRef]
- Skipper, A.; Sims, J.N.; Yedjou, C.G.; Tchounwou, P.B. Cadmium chloride induces DNA damage and apoptosis of human liver carcinoma cells via oxidative stress. Int. J. Environ. Res. Public Health 2016, 13, 88. [Google Scholar] [CrossRef]
- Rikans, L.E.; Yamano, T. Mechanisms of cadmium-mediated acute hepatotoxicity. J. Biochem. Mol. Toxicol. 2000, 14, 110–117. [Google Scholar] [CrossRef]
- Shaikh, Z.A.; Vu, T.T.; Zaman, K. Oxidative stress as a mechanism of chronic cadmium-induced hepatotoxicity and renal toxicity and protection by antioxidants. Toxicol. Appl. Pharmacol. 1999, 154, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Ma, H.; Liu, G.; Fan, S.; Guo, Z. Mechanism of cadmium exposure induced hepatotoxicity in the mud crab (Scylla paramamosain): Activation of oxidative stress and Nrf2 signaling pathway. Antioxidants 2022, 11, 978. [Google Scholar] [CrossRef]
- Abdel-Moneim, W.M.; Ghafeer, H.H. The potential protective effect of natural honey against cadmium-induced hepatotoxicity and nephrotoxicity. Mansoura J. Forensic Med. Clin. Toxicol. 2007, 15, 75–98. [Google Scholar] [CrossRef]
- Trzcinka-Ochocka, M.; Jakubowski, M.; Razniewska, G.; Halatek, T.; Gazewski, A. The effects of environmental cadmium exposure on kidney function: The possible influence of age. Environ. Res. 2004, 95, 143–150. [Google Scholar] [CrossRef]
- Dudley, R.E.; Gammal, L.M.; Klaassen, C.D. Cadmium-induced hepatic and renal injury in chronically exposed rats: Likely role of hepatic cadmium-metallothionein in nephrotoxicity. Toxicol. Appl. Pharmacol. 1985, 77, 414–426. [Google Scholar] [CrossRef]
- Renugadevi, J.; Prabu, S.M. Naringenin protects against cadmium-induced oxidative renal dysfunction in rats. Toxicology 2009, 256, 128–134. [Google Scholar] [CrossRef]
- Branca, J.J.V.; Pacini, A.; Gulisano, M.; Taddei, N.; Fiorillo, C.; Becatti, M. Cadmium-InducedCytotoxicity: Effects on MitochondrialElectron Transport Chain. Front. Cell Dev. Biol. 2020, 8, 604377. [Google Scholar] [CrossRef] [PubMed]
- Iwagami, Y. Changes in the ultrastructure of human cells related to certain biological responses under hyperthermic culture conditions. Human Cell 1996, 9, 353–366. [Google Scholar] [PubMed]
- Sabir, S.; Akash, M.S.H.; Fiayyaz, F.; Saleem, U.; Mehmood, M.H.; Rehman, K. Role of cadmium and arsenic as endocrine disruptors in the metabolism of carbohydrates: Inserting the association into perspectives. Biomed. Pharmacother. 2019, 114, 108802. [Google Scholar] [CrossRef] [PubMed]
- Das, S.C.; Al-Naemi, H.A. Cadmium toxicity: Oxidative stress, inflammation and tissue injury. Occup. Dis. Environ. Med. 2019, 7, 144–163. [Google Scholar] [CrossRef]
- Farag, M.R.; Elhady, W.M.; Ahmed, S.Y.A.; Taha, H.S.A.; Alagawany, M. Astragalus polysaccharides alleviate tilmicosin- induced toxicity in rats byinhibiting oxidative damage and modulating the expressions of HSP70, NF-kB and Nrf2/HO-1 pathway. Res. Vet. Sci. 2019, 124, 137–148. [Google Scholar] [CrossRef]
- Jeon, J.-Y.; Kim, K.-E.; Im, H.-J.; Oh, S.-T.; Lim, S.-U.; Kwon, H.-S.; Moon, B.-H.; Kim, J.-M.; An, B.-K.; Kang, C.-W. The production of lutein-enriched eggs with dietary Chlorella. Food Sci. Anim. Resour. 2012, 32, 13–17. [Google Scholar] [CrossRef]
- Buono, S.; Langellotti, A.L.; Martello, A.; Rinna, F.; Fogliano, V. Functional ingredients from microalgae. Food Funct. 2014, 5, 1669–1685. [Google Scholar] [CrossRef]
- Bauer, L.M.; Costa, J.A.V.; da Rosa, A.P.C.; Santos, L.O. Growth stimulation and synthesis of lipids, pigments and antioxidants with magnetic fields in Chlorella kessleri cultivations. Bioresour. Technol. 2017, 244, 1425–1432. [Google Scholar] [CrossRef]
- Bengwayan, P.T.; Laygo, J.C.; Pacio, A.E.; Poyaoan, J.L.Z.; Rebugio, J.F.; Yuson, A.L.L. A comparative study on the antioxidant property of Chlorella (Chlorella sp.) tablet and glutathione tablet. E-Int. Sci. Res. J. 2010, 2, 25–35. [Google Scholar]
- Plaza, M.; Herrero, M.; Cifuentes, A.; Ibanez, E. Innovative natural functional ingredients from microalgae. J. Agric. Food Chem. 2009, 57, 7159–7170. [Google Scholar] [CrossRef]
- Sheih, I.-C.; Fang, T.J.; Wu, T.-K. Isolation and characterisation of a novel angiotensin I-converting enzyme (ACE) inhibitory peptide from the algae protein waste. Food Chem. 2009, 115, 279–284. [Google Scholar] [CrossRef]
- Ko, S.-C.; Kim, D.; Jeon, Y.-J. Protective effect of a novel antioxidative peptide purified from a marine Chlorella ellipsoidea protein against free radical-induced oxidative stress. Food Chem. Toxicol. 2012, 50, 2294–2302. [Google Scholar] [CrossRef]
- Cherng, J.-Y.; Shih, M.-F. Preventing dyslipidemia by Chlorella pyrenoidosa in rats and hamsters after chronic high fat diet treatment. Life Sci. 2005, 76, 3001–3013. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, X. Separation, antitumor activities, and encapsulation of polypeptide from Chlorella pyrenoidosa. Biotechnol. Prog. 2013, 29, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Cherng, J.-Y.; Shih, M.-F. Improving glycogenesis in Streptozocin (STZ) diabetic mice after administration of green algae Chlorella. Life Sci. 2006, 78, 1181–1186. [Google Scholar] [CrossRef]
- Peng, H.-Y.; Chu, Y.-C.; Chen, S.-J.; Chou, S.-T. Hepatoprotection of chlorella against carbon tetrachloride-induced oxidative damage in rats. In Vivo 2009, 23, 747–754. [Google Scholar] [PubMed]
- Guzman, S.; Gato, A.; Calleja, J. Antiinflammatory, analgesic and free radical scavenging activities of the marine microalgae Chlorella stigmatophora and Phaeodactylum tricornutum. Phytother. Res. 2001, 15, 224–230. [Google Scholar] [CrossRef]
- Teresa, V.-O.M.; Octavio, G.-F.; Nayelly, J.-S.; Mayumi, R.-V.; Alan, M.-S.; Yvonne, T.-J.; Edgar, M.-R.; Santos, B.-S.; Angel, G.-C.C. The administration of Chlorella vulgaris protects against nefrotoxicity caused by CCl4. RETEL Revista de Toxicologia 2011, 35, 1–13. [Google Scholar]
- Huang, Z.; Li, L.; Huang, G.; Yan, Q.; Shi, B.; Xu, X. Growth-inhibitory and metal-binding proteins in Chlorella vulgaris exposed to cadmium or zinc. Aquat. Toxicol. 2009, 91, 54–61. [Google Scholar] [CrossRef]
- Wang, L.; Liu, J.; Filipiak, M.; Mungunkhuyag, K.; Jedynak, P.; Burczyk, J.; Fu, P.; Malec, P. Fast and efficient cadmium biosorption by Chlorella vulgaris K-01 strain: The role of cell walls in metal sequestration. Algal Res. 2021, 60, 102497. [Google Scholar] [CrossRef]
- Yadav, M.; Kumar, V.; Sandal, N.; Chauhan, M.K. Quantitative evaluation of Chlorella vulgaris for removal of toxic metals from body. J. Appl. Phycol. 2022, 34, 2743–2754. [Google Scholar] [CrossRef]
- Dacie, J.V.; Lewis, S.M. Practical Haematology; Churchill Livingstone: London, UK; New York, NY, USA, 1984. [Google Scholar]
- Armstrong, W.D.; Carr, C.W. Physiological Chemistry Laboratory Direction, 3rd ed.; Burses Publishing Co.: Minneopollis, MN, USA, 1964. [Google Scholar]
- Wise, W.A. Determination of Serum Albumen; King: London, UK, 1965; p. 273. [Google Scholar]
- Deeg, R.; Ziegenohrm, J. Kinetic enzymatic method forautomated determination of total cholesterol in serum. J. Clin. Chem. 1983, 29, 1798–1802. [Google Scholar] [CrossRef]
- Friedewald, W.T.; Levy, R.I.; Fredrickson, D.S. Estimation ofthe concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin. Chem. 1972, 18, 499–502. [Google Scholar] [CrossRef] [PubMed]
- Fossati, P.; Prencipe, L. Serum triglycerides determined colourimetrically with an enzyme that produces hydrogen peroxide. Clin. Chem. 1982, 28, 2077–2080. [Google Scholar] [CrossRef] [PubMed]
- Belfield, A.; Goldberg, D.M. Revised assay for serum phenylphosphatase activity using 4-amino-antipyrine. Enzyme 1971, 12, 561–573. [Google Scholar] [CrossRef]
- Reitman, S.; Frankel, S. A colorimetric method for thedetermination of serum glutamic oxaloacetic and glutamic pyruvic transaminases. Am. J. Clin. Pathol. 1957, 28, 56–63. [Google Scholar] [CrossRef]
- Bianchi, A.T.; Moonen-Leusen, H.W.; Van der Heijden, P.J.; Bokhout, B.A. The use of a double antibody sandwich ELISA and monoclonal antibodies for the assessment of porcine IgM, IgG and IgA concentrations. Vet. Immunol. Immunopathol. 1995, 44, 309–317. [Google Scholar] [CrossRef]
- Vidal-Vanaclocha, F.; Fantuzzi, G.; Mendoza, L.; Fuentes, A.M.; Anasagasti, M.J.; Martin, J.; Carrascal, T.; Walsh, P.; Reznikov, L.L.; Kim, S.-H.; et al. IL-18 regulates IL-1-beta-dependent hepatic melanoma metastasis via vascular cell adhesion molecule-1. Proc. Nat. Acad. Sci. USA 2000, 97, 734–739. [Google Scholar] [CrossRef]
- Koracevic, D.; Koracevic, G.; Djordjevic, V.; Andrejevic, S.; Cosic, V. Method for the measurement of antioxidant activity in human fluids. J. Clin. Pathol. 2001, 54, 356–361. [Google Scholar] [CrossRef]
- Preuss, H.G.; Jarrell, S.T.; Scheckenbach, R.; Lieberman, S.; Anderson, R.A. Comparative effects of chromium, vanadium andgymnema sylvestre on sugar-induced blood pressure elevations in SHR. J. Am. Coll. Nutr. 1998, 17, 116–123. [Google Scholar] [CrossRef]
- Khalil, S.R.; Awad, A.; Mohammed, H.H.; Nassan, M.A. Imidacloprid insecticide exposure induces stress and disrupts glucose homeostasis inmale rats. Environ. Toxicol. Pharmacol. 2017, 55, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Pakzad, M.; Fouladdel, S.; Nili-Ahmadabadi, A.; Pourkhalili, N.; Baeeri, M.; Azizi, E.; Sabzevari, O.; Ostad, S.N.; Abdollahi, M. Sublethal exposures of diazinon alters glucose homostasis in Wistar rats: Biochemical and molecular evidences of oxidative stress in adipose tissues. Pestic. Biochem. Physiol. 2013, 105, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Zhang, X.; Cheng, Y.; Abdelnasir, A.; Tang, S.; Kemper, N.; Hartung, J.; Bao, E. Association of heat shock protein 70 expression with rat myocardial cell damage during heat stress in vitro and in vivo. Genet. Mol. Res. 2015, 14, 1994–2005. [Google Scholar] [CrossRef] [PubMed]
- Sang, L.; Wang, X.-M.; Xu, D.-Y.; Sang, L.-X.; Han, Y.; Jiang, L.-Y. Morin enhances hepatic Nrf2 expression in a liver fibrosis rat model. World J. Gastroenterol. 2017, 23, 8334. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Suvarna, K.S.; Layton, C.; Bancroft, J.D. Bancroft’s Theory and Practice of Histological Techniques E-Book; Elsevier Health Sciences: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Shen, H.-H.; Huang, S.-Y.; Cheng, P.-Y.; Chu, Y.-J.; Chen, S.-Y.; Lam, K.-K.; Lee, Y.-M. Involvement of HSP70 and HO-1 in the protective effects of raloxifene on multiple organ dysfunction syndrome by endotoxemia in ovariectomized rats. Menopause 2017, 24, 959–969. [Google Scholar] [CrossRef]
- Luu Quoc, Q.; Cao Thi Bich, T.; Kim, S.H.; Park, H.S.; Shin, Y.S. Administration of vitamin E attenuates airway inflammation through restoration of Nrf2 in a mouse model of asthma. J. Cell. Mol. Med. 2021, 25, 6721–6732. [Google Scholar] [CrossRef]
- Saygi, A.; Deniz, G.; Kutsal, O.; Vural, N. Chronic effects of cadmium on kidney, liver, testis, and fertility of male rats. Biol. Trace Elem. Res. 1991, 31, 209–214. [Google Scholar] [CrossRef]
- Gonick, H. Nephrotoxicity of cadmium & lead. Indian J. Med. Res. 2008, 128, 335–352. [Google Scholar]
- Branca, J.J.V.; Morucci, G.; Pacini, A. Cadmium-induced neurotoxicity: Still much ado. Neural Regen. Res. 2018, 13, 1879. [Google Scholar]
- Huff, J.; Lunn, R.M.; Waalkes, M.P.; Tomatis, L.; Infante, P.F. Cadmium-induced cancers in animals and in humans. Int. J. Occup. Environ. Health 2007, 13, 202–212. [Google Scholar] [CrossRef]
- Hassoun, E.A.; Stohs, S.J. Cadmium-induced production of superoxide anion and nitric oxide, DNA single strand breaks and lactate dehydrogenase leakage in J774A. 1 cell cultures. Toxicology 1996, 112, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Hart, B.; Lee, C.; Shukla, G.; Shukla, A.; Osier, M.; Eneman, J.; Chiu, J.-F. Characterization of cadmium-induced apoptosis in rat lung epithelial cells: Evidence for the participation of oxidant stress. Toxicology 1999, 133, 43–58. [Google Scholar] [CrossRef] [PubMed]
- Patra, R.; Rautray, A.K.; Swarup, D. Oxidative stress in lead and cadmium toxicity and its amelioration. Vet. Med. Int. 2011, 2011, 457327. [Google Scholar] [CrossRef] [PubMed]
- Khan, T.A.; Zafar, F. Haematological study in response to varying doses of estrogen in broiler chicken. Int. J. Poult. Sci. 2005, 4, 748–751. [Google Scholar]
- Andjelkovic, M.; Buha Djordjevic, A.; Antonijevic, E.; Antonijevic, B.; Stanic, M.; Kotur-Stevuljevic, J.; Spasojevic-Kalimanovska, V.; Jovanovic, M.; Boricic, N.; Wallace, D. Toxic effect of acute cadmium and lead exposure in rat blood, liver, and kidney. Int. J. Environ. Res. Public Health 2019, 16, 274. [Google Scholar] [CrossRef]
- Dökmeci, A.H.; Karaboğa, İ.; Güzel, S.; Erboğa, Z.F.; Yılmaz, A. Toxicological assessment of low-dose bisphenol A, lead and endosulfan combination: Chronic toxicity study in male rats. Environ. Sci. Pollut. Res. 2021, 29, 10558–10574. [Google Scholar] [CrossRef]
- Farag, M.R.; Alagawany, M.; Abd El-Hack, M.E.; El-Sayed, S.A.; Ahmed, S.Y.; Samak, D.H. Yucca schidigera extract modulates the lead-induced oxidative damage, nephropathy and altered inflammatory response and glucose homeostasis in Japanese quails. Ecotoxicol. Environ. Saf. 2018, 156, 311–321. [Google Scholar] [CrossRef]
- Pawlikowska-Pawlęga, B.; Gruszecki, W.I.; Misiak, L.; Paduch, R.; Piersiak, T.; Zarzyka, B.; Pawelec, J.; Gawron, A. Modification of membranes by quercetin, a naturally occurring flavonoid, via its incorporation in the polar head group. Biochim. Biophys. Acta (BBA)-Biomembr. 2007, 1768, 2195–2204. [Google Scholar] [CrossRef]
- Safi, C.; Zebib, B.; Merah, O.; Pontalier, P.-Y.; Vaca-Garcia, C. Morphology, composition, production, processing and applications of Chlorella vulgaris: A review. Renew. Sustain. Energy Rev. 2014, 35, 265–278. [Google Scholar] [CrossRef]
- Mathew, B.; Sankaranarayanan, R.; Nair, P.P.; Varghese, C.; Somanathan, T.; Amma, B.P.; Amma, N.S.; Nair, M.K. Evaluation of chemoprevention of oral cancer with Spirulina fusiformis. Nutr. Cancer 1995, 24, 197–202. [Google Scholar] [CrossRef]
- Rana, S.; Singh, R.; Verma, S. Protective effects of few antioxidants on liver function in rats treated with cadmium and mercury. Indian. J. Exp. Biol. 1996, 34, 177–179. [Google Scholar] [PubMed]
- Patil, G. Role of ascorbic acid on mercuric chloride toxicity in vital organs of mice. Indian J. Environ. Toxicol. 1999, 9, 53–55. [Google Scholar]
- Xu, W.; Gao, Z.; Qi, Z.; Qiu, M.; Peng, J.-q.; Shao, R. Effect of dietary Chlorella on the growth performance and physiological parameters of gibel carp, Carassius auratus gibelio. Turk. J. Fish. Aquat. Sci. 2014, 14, 53–57. [Google Scholar]
- Kang, H.; Salim, H.; Akter, N.; Kim, D.; Kim, J.; Bang, H.; Kim, M.; Na, J.; Hwangbo, J.; Choi, H. Effect of various forms of dietary Chlorella supplementation on growth performance, immune characteristics, and intestinal microflora population of broiler chickens. J. Appl. Poult. Res. 2013, 22, 100–108. [Google Scholar] [CrossRef]
- Yap, C.Y.; Aw, T.C. Liver function tests (LFTs). Proc. Singap. Healthc. 2010, 19, 80–82. [Google Scholar] [CrossRef]
- Philip, G.; Reddy, P.; Sridevi, G. Cypermethrin-induced in vivo alterations in the carbohydrate metabolism of freshwater fish, Labeo rohita. Ecotoxicol. Environ. Saf. 1995, 31, 173–178. [Google Scholar] [CrossRef]
- Shimada, Y.; Wiget, P.; Gulli, M.P.; Bi, E.; Peter, M. The nucleotide exchange factor Cdc24p may be regulated by auto-inhibition. EMBO J. 2004, 23, 1051–1062. [Google Scholar] [CrossRef]
- Renugadevi, J.; Prabu, S.M. Cadmium-induced hepatotoxicity in rats and the protective effect of naringenin. Exp. Toxicol. Pathol. 2010, 62, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Farag, M.R.; Alagawany, M. Erythrocytes as a biological model for screening of xenobiotics toxicity. Chem.-Biol. Interact. 2018, 279, 73–83. [Google Scholar] [CrossRef]
- Alagawany, M.; Abd El-Hack, M.; Farag, M.; Elnesr, S.; El-Kholy, M.; Saadeldin, I.; Swelum, A. Dietary supplementation of Yucca schidigera extract enhances productive and reproductive performances, blood profile, immune function, and antioxidant status in laying Japanese quails exposed to lead in the diet. Poult. Sci. 2018, 97, 3126–3137. [Google Scholar] [CrossRef]
- Wu, C.; Zhang, Y.; Chai, L.; Wang, H. Histological changes, lipid metabolism and oxidative stress in the liver of Bufo gargarizans exposed to cadmium concentrations. Chemosphere 2017, 179, 337–346. [Google Scholar] [CrossRef]
- Theocharis, S.; Margeli, A.; Fasitsas, C.; Loizidou, M.; Deliconstantinos, G. Acute exposure to cadmium causes time-dependent liver injury in rats. Comp. Biochem. Physiol. Part C Comp. Pharmacol. 1991, 99, 127–130. [Google Scholar] [CrossRef]
- Sudo, J.-i.; Hayashi, T.; Kimura, S.-i.; Kakuno, K.; Terui, J.; Takashima, K.; Soyama, M. Mechanism of nephrotoxicity induced by repeated administration of cadmium chloride in rats. J. Toxicol. Environ. Health 1996, 48, 333–348. [Google Scholar] [CrossRef] [PubMed]
- Thijssen, S.; Maringwa, J.; Faes, C.; Lambrichts, I.; Van Kerkhove, E. Chronic exposure of mice to environmentally relevant, low doses of cadmium leads to early renal damage, not predicted by blood or urine cadmium levels. Toxicology 2007, 229, 145–156. [Google Scholar] [CrossRef]
- Choi, J.-H.; Rhee, S.-J. Effects of vitamin E on renal dysfunction in chronic cadmium-poisoned rats. J. Med. Food 2003, 6, 209–215. [Google Scholar] [CrossRef]
- Damek-Poprawa, M.; Sawicka-Kapusta, K. Histopathological changes in the liver, kidneys, and testes of bank voles environmentally exposed to heavy metal emissions from the steelworks and zinc smelter in Poland. Environ. Res. 2004, 96, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Jemai, H.; Lachkar, H.A.; Messaoudi, I.; Kerkeni, A. Effects of zinc pre-treatment on blood glutathione, serum zinc and kidney histological organisation in male rats exposed to cadmium. J. Trace Elem. Med. Biol. 2010, 24, 277–282. [Google Scholar] [CrossRef]
- Herman, J.P.; McKlveen, J.M.; Ghosal, S.; Kopp, B.; Wulsin, A.; Makinson, R.; Scheimann, J.; Myers, B. Regulation of the hypothalamic-pituitary-adrenocortical stress response. Compr. Physiol. 2016, 6, 603. [Google Scholar] [PubMed]
- Queiroz, J.S.; Blasco, I.M.; Gagliano, H.; Daviu, N.; Román, A.G.; Belda, X.; Carrasco, J.; Rocha, M.C.; Neto, J.P.; Armario, A. Chlorella vulgaris reduces the impact of stress on hypothalamic–pituitary–adrenal axis and brain c-fos expression. Psychoneuroendocrinology 2016, 65, 1–8. [Google Scholar] [CrossRef]
- Sahin, K.; Orhan, C.; Akdemir, F.; Tuzcu, M.; Iben, C.; Sahin, N. Resveratrol protects quail hepatocytes against heat stress: Modulation of the Nrf2 transcription factor and heat shock proteins. J. Anim. Physiol. Anim. Nutr. 2012, 96, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Coronado-Reyes, J.A.; Acosta-Ramírez, E.; Martínez-Olguín, M.V.; González-Hernández, J.C. Antioxidant Activity and Kinetic Characterization of Chlorella vulgaris Growth under Flask-Level Photoheterotrophic Growth Conditions. Molecules 2022, 27, 6346. [Google Scholar] [CrossRef] [PubMed]
- Dodson, M.; Redmann, M.; Rajasekaran, N.S.; Darley-Usmar, V.; Zhang, J. KEAP1–NRF2 signalling and autophagy in protection against oxidative and reductive proteotoxicity. Biochem. J. 2015, 469, 347–355. [Google Scholar] [CrossRef]
- Zhao, P.; Piao, X.; Pan, L.; Zeng, Z.; Li, Q.; Xu, X.; Wang, H. Forsythia suspensa extract attenuates lipopolysaccharide-induced inflammatory liver injury in rats via promoting antioxidant defense mechanisms. Anim. Sci. J. 2017, 88, 873–881. [Google Scholar] [CrossRef]
- Na, H.-K.; Surh, Y.-J. Oncogenic potential of Nrf2 and its principal target protein heme oxygenase-1. Free. Radic. Biol. Med. 2014, 67, 353–365. [Google Scholar] [CrossRef] [PubMed]
- Elsherbiny, N.M.; Eladl, M.A.; Al-Gayyar, M.M. Renal protective effects of arjunolic acid in a cisplatin-induced nephrotoxicity model. Cytokine 2016, 77, 26–34. [Google Scholar] [CrossRef]
- Oku, H.; Nakazato, H.; Horikawa, T.; Tsuruta, Y.; Suzuki, R. Pirfenidone suppresses tumor necrosis factor-α, enhances interleukin-10 and protects mice from endotoxic shock. Eur. J. Pharmacol. 2002, 446, 167–176. [Google Scholar] [CrossRef]
- Hsu, C.-M.; Wang, J.-S.; Liu, C.-H.; Chen, L.-W. Kupffer cells protect liver from ischemia-reperfusion injury by an inducible nitric oxide synthase-dependent mechanism. Shock 2002, 17, 280–285. [Google Scholar] [CrossRef]
- Hon, W.M.; Lee, K.H.; Khoo, H.E. Nitric oxide in liver diseases: Friend, foe, or just passerby? Ann. N. Y. Acad. Sci. 2002, 962, 275–295. [Google Scholar] [CrossRef]
- Tamura, Y.; Torigoe, T.; Kukita, K.; Saito, K.; Okuya, K.; Kutomi, G.; Hirata, K.; Sato, N. Heat-shock proteins as endogenous ligands building a bridge between innate and adaptive immunity. Immunotherapy 2012, 4, 841–852. [Google Scholar] [CrossRef]
- Ohashi, K.; Burkart, V.; Flohé, S.; Kolb, H. Cutting edge: Heat shock protein 60 is a putative endogenous ligand of the toll-like receptor-4 complex. J. Immunol. 2000, 164, 558–561. [Google Scholar] [CrossRef]
- Ahmed, M.B.; Khater, M.R. Evaluation of the protective potential of Ambrosia maritima extract on acetaminophen-induced liver damage. J. Ethnopharmacol. 2001, 75, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Garcia, I.; Guil-Guerrero, J.L. Evaluation of the antioxidant activity of three microalgal species for use as dietary supplements and in the preservation of foods. Food Chem. 2008, 108, 1023–1026. [Google Scholar] [CrossRef]
- Cheng, D.; Wan, Z.; Zhang, X.; Li, J.; Li, H.; Wang, C. Dietary Chlorella vulgaris ameliorates altered immunomodulatory functions in cyclophosphamide-induced immunosuppressive mice. Nutrients 2017, 9, 708. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, F.; Fanning, K.; Netzel, M.; Turner, W.; Li, Y.; Schenk, P.M. Profiling of carotenoids and antioxidant capacity of microalgae from subtropical coastal and brackish waters. Food Chem. 2014, 165, 300–306. [Google Scholar] [CrossRef]
- Machu, L.; Misurcova, L.; Vavra Ambrozova, J.; Orsavova, J.; Mlcek, J.; Sochor, J.; Jurikova, T. Phenolic content and antioxidant capacity in algal food products. Molecules 2015, 20, 1118–1133. [Google Scholar] [CrossRef] [PubMed]
- Martins, N.; Barros, L.; Ferreira, I.C. In vivo antioxidant activity of phenolic compounds: Facts and gaps. Trends Food Sci. Technol. 2016, 48, 1–12. [Google Scholar] [CrossRef]
- Renugadevi, K.; Nachiyar, C.V.; Sowmiya, P.; Sunkar, S. Antioxidant activity of phycocyanin pigment extracted from marine filamentous cyanobacteria Geitlerinema sp TRV57. Biocatal. Agric. Biotechnol. 2018, 16, 237–242. [Google Scholar] [CrossRef]
- El Latif, A.A.; Assar, D.H.; Elkaw, E.M.; Hamza, H.A.; Alkhalifah, D.H.; Hozzein, W.N.; Hamouda, R.A. Protective role of Chlorella vulgaris with Thiamine against Paracetamol induced toxic effects on haematological, biochemical, oxidative stress parameters and histopathological changesin Wistar rats. Sci. Rep. 2021, 11, 3911. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | ChV | Cd | Cd + ChV | p Value | |
|---|---|---|---|---|---|
| RBCs (106/mm3) | 8.35 ± 0.12 | 8.37 ± 0.12 | 7.21 ± 0.56 | 8.06 ± 0.06 | 0.074 |
| Hb (gm/dL) | 13.38 ± 0.35 a | 13.24 ± 0.16 a | 8.68 ± 0.29 b | 12.44 ± 0.12 a | <0.001 |
| PCV % | 50.39 ± 0.17 a | 50.91 ± 0.94 a | 43.44 ± 0.51 c | 46.66 ± 0.40 b | <0.001 |
| WBC (103/mm3) | 8.52 ± 0.07 a | 8.54 ± 0.09 a | 6.21 ± 0.01 c | 7.20 ± 0.13 b | <0.001 |
| Lymphocyte (103/mm3) | 4.34 ± 0.23 a | 4.69 ± 0.07 a | 3.38 ± 0.13 b | 4.18 ± 0.14 a | 0.002 |
| Neutrophil (103/mm3) | 2.71 ± 1.35 | 4.06 ± 0.01 | 4.02 ± 0.01 | 4.04 ± 0.02 | 0.452 |
| Esinophil (103/mm3) | 0.82 ± 0.01 | 0.83 ± 0.01 | 0.82 ± 0.01 | 0.82 ± 0.01 | 0.583 |
| Control | ChV | Cd | Cd + ChV | p Value | |
|---|---|---|---|---|---|
| Initial weight | 185.31 ± 0.08 | 185.29 ± 0.09 | 185.30 ± 0.08 | 185.30 ± 0.07 | 0.997 |
| Final weight | 209.59 ± 0.47 b | 215.55 ± 1.70 a | 200.84 ± 0.41 c | 209.14 ± 0.12 b | <0.001 |
| Relative liver weight | 3.30 ± 0.01 | 3.30 ± 0.01 | 3.29 ± 0.01 | 3.29 ± 0.02 | 0.802 |
| Relative kidney weight | 0.79 ± 0.02 | 0.78 ± 0.02 | 0.79 ± 0.02 | 0.78 ± 0.03 | 0.995 |
| Relative spleen weight | 0.38 ± 0.01 | 0.36 ± 0.01 | 0.38 ± 0.01 | 0.37 ± 0.01 | 0.640 |
| Control | ChV | Cd | Cd + ChV | p Value | |
|---|---|---|---|---|---|
| Liver function markers | |||||
| Total protein(g/dL) | 8.04 ± 0.04 a | 8.12 ± 0.01 a | 5.05 ± 0.03 c | 7.28 ± 0.15 b | <0.001 |
| Albumin(g/dL) | 5.01 ± 0.01 a | 5.08 ± 0.06 a | 2.52 ± 0.01 c | 4.84 ± 0.04 b | <0.001 |
| ALT(U/L) | 28.35 ± 0.12 c | 28.16 ± 0.02 c | 121.17 ± 0.07 a | 82.87 ± 03.68 b | <0.001 |
| AST(U/L) | 67.45 ± 0.41 c | 66.55 ± 01.01 c | 151.44 ± 01.32 a | 113.90 ± 03.12 b | <0.001 |
| ALP(U/L) | 112.53 ± 0.25 c | 110.38 ± 0.08 c | 196.81 ± 01.14 a | 150.24 ± 05.51 b | <0.001 |
| Kidney function markers | |||||
| Urea(mg/dL) | 18.75 ± 0.12 c | 17.21 ± 0.01 c | 41.32 ± 0.61 a | 23.75 ± 1.21 b | <0.001 |
| Creatinine (mg/dL) | 0.69 ± 0.00 b | 0.68 ± 0.01 b | 1.01 ± 0.01 a | 0.69 ± 0.01 b | <0.001 |
| Lipid profile | |||||
| Triglycerides (mg/dL) | 100.20 ± 0.42 | 98.62 ± 0.69 | 105.39 ± 0.35 | 103.97 ± 01.26 | 0.447 |
| Total cholesterol (mg/dL) | 85.97 ± 0.92 c | 85.58 ± 0.32 c | 157.35 ± 0.82 a | 122.47 ± 1.52 b | <0.001 |
| HDL-cholesterol (mg/dL) | 50.28 ± 0.04 a | 50.12 ± 0.05 ab | 30.29 ± 0.09 c | 49.36 ± 0.41 b | <0.001 |
| LDL-cholesterol(mg/dL) | 30.21 ± 0.03 c | 30.18 ± 0.03 c | 122.59 ± 1.20 a | 70.35 ± 0.11 b | <0.001 |
| Control | ChV | Cd | Cd + V | p Value | |
|---|---|---|---|---|---|
| Oxidative stress markers | |||||
| TAC (μmol/mL) | 3.46 ± 0.13 b | 4.14 ± 0.12 a | 1.02 ± 0.01 d | 2.87 ± 0.01 c | <0.001 |
| ROS (U/mL) | 15.55 ± 0.10 c | 15.85 ± 0.09 c | 30.24 ± 0.02 a | 18.30 ± 0.06 b | <0.001 |
| MDA (nmol/mL) | 5.01 ± 0.01 b | 4.63 ± 0.20 b | 13.49 ± 0.58 a | 5.03 ± 0.01 b | <0.001 |
| Immunity biomarkers | |||||
| IgG (mg/dL) | 8.21 ± 0.01 b | 8.73 ± 0.09 a | 5.26 ± 0.10 c | 8.20 ± 0.01 b | <0.001 |
| IgM (mg/dL) | 1.33 ± 0.07 | 1.39 ± 0.11 | 1.30 ± 0.06 | 1.31 ± 0.06 | 0.843 |
| Inflammatory markers | |||||
| TNF-α (pg/mL) | 63.26± 1.54 c | 60.06 ± 0.10 c | 99.14 ± 0.27 a | 75.20 ± 2.51 b | <0.001 |
| IL-1β (pg/mL) | 72.02 ± 0.24 b | 70.17 ± 0.10 c | 131.04 ± 0.55 a | 131.04 ± 0.55 a | <0.001 |
| NO (Umol/L) | 51.19 ± 1.01 c | 50.13 ± 0.07 c | 81.83 ± 1.13 a | 56.38 ± 0.45 b | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farag, M.R.; Alagawany, M.; Mahdy, E.A.A.; El-Hady, E.; Abou-Zeid, S.M.; Mawed, S.A.; Azzam, M.M.; Crescenzo, G.; Abo-Elmaaty, A.M.A. Benefits of Chlorella vulgaris against Cadmium Chloride-Induced Hepatic and Renal Toxicities via Restoring the Cellular Redox Homeostasis and Modulating Nrf2 and NF-KB Pathways in Male Rats. Biomedicines 2023, 11, 2414. https://doi.org/10.3390/biomedicines11092414
Farag MR, Alagawany M, Mahdy EAA, El-Hady E, Abou-Zeid SM, Mawed SA, Azzam MM, Crescenzo G, Abo-Elmaaty AMA. Benefits of Chlorella vulgaris against Cadmium Chloride-Induced Hepatic and Renal Toxicities via Restoring the Cellular Redox Homeostasis and Modulating Nrf2 and NF-KB Pathways in Male Rats. Biomedicines. 2023; 11(9):2414. https://doi.org/10.3390/biomedicines11092414
Chicago/Turabian StyleFarag, Mayada R., Mahmoud Alagawany, Eman A. A. Mahdy, Enas El-Hady, Shimaa M. Abou-Zeid, Suzan A. Mawed, Mahmoud M. Azzam, Giuseppe Crescenzo, and Azza M. A. Abo-Elmaaty. 2023. "Benefits of Chlorella vulgaris against Cadmium Chloride-Induced Hepatic and Renal Toxicities via Restoring the Cellular Redox Homeostasis and Modulating Nrf2 and NF-KB Pathways in Male Rats" Biomedicines 11, no. 9: 2414. https://doi.org/10.3390/biomedicines11092414
APA StyleFarag, M. R., Alagawany, M., Mahdy, E. A. A., El-Hady, E., Abou-Zeid, S. M., Mawed, S. A., Azzam, M. M., Crescenzo, G., & Abo-Elmaaty, A. M. A. (2023). Benefits of Chlorella vulgaris against Cadmium Chloride-Induced Hepatic and Renal Toxicities via Restoring the Cellular Redox Homeostasis and Modulating Nrf2 and NF-KB Pathways in Male Rats. Biomedicines, 11(9), 2414. https://doi.org/10.3390/biomedicines11092414