
Induction of Skin Cancer by Long-Term Blue Light Irradiation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Experiments
2.2. Preparation and Staining of Dorsal Skin
2.3. Western Blotting of Dorsal Skin Proteins
2.4. Measurement of IL-10, TGF-β, IL-6, and IL-23 Levels in Plasma and Ki67, Cyclin D1, Neutrophil Elastase, and Reactive Oxygen Species Levels (ROS) in Dorsal Skin
2.5. Statistical Analysis
3. Results
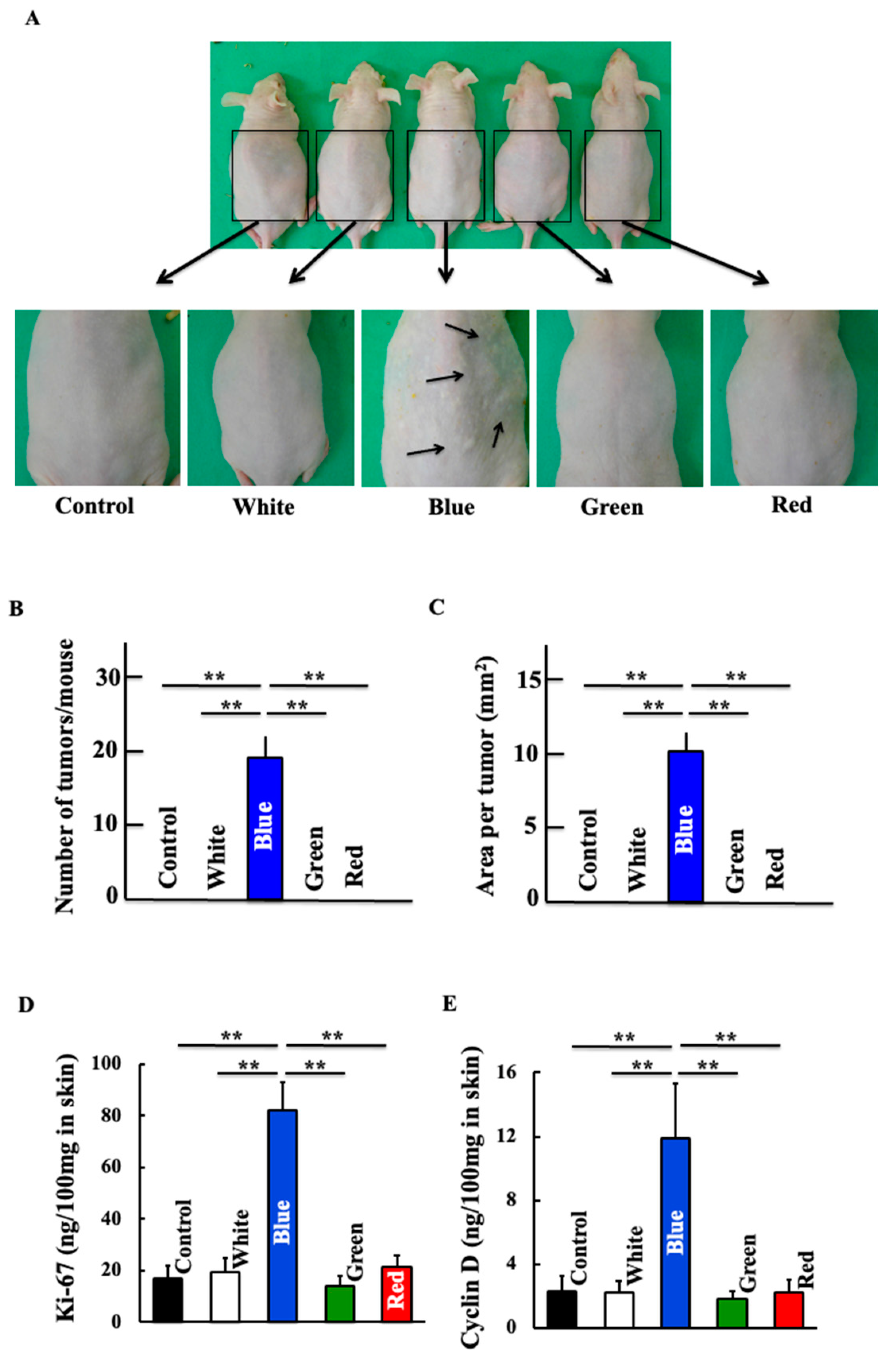
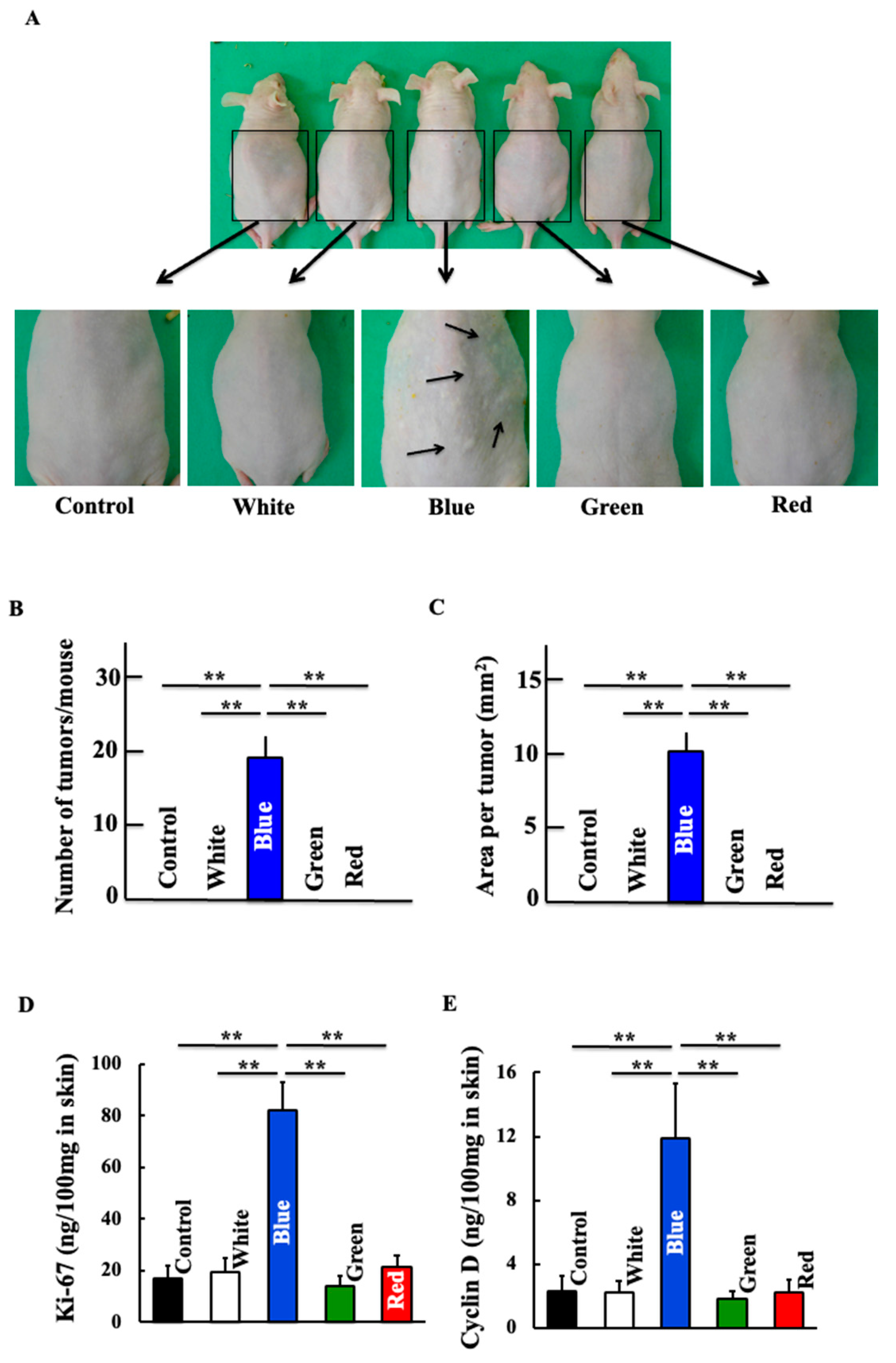
3.1. Effects of Blue Light on Skin Cancer
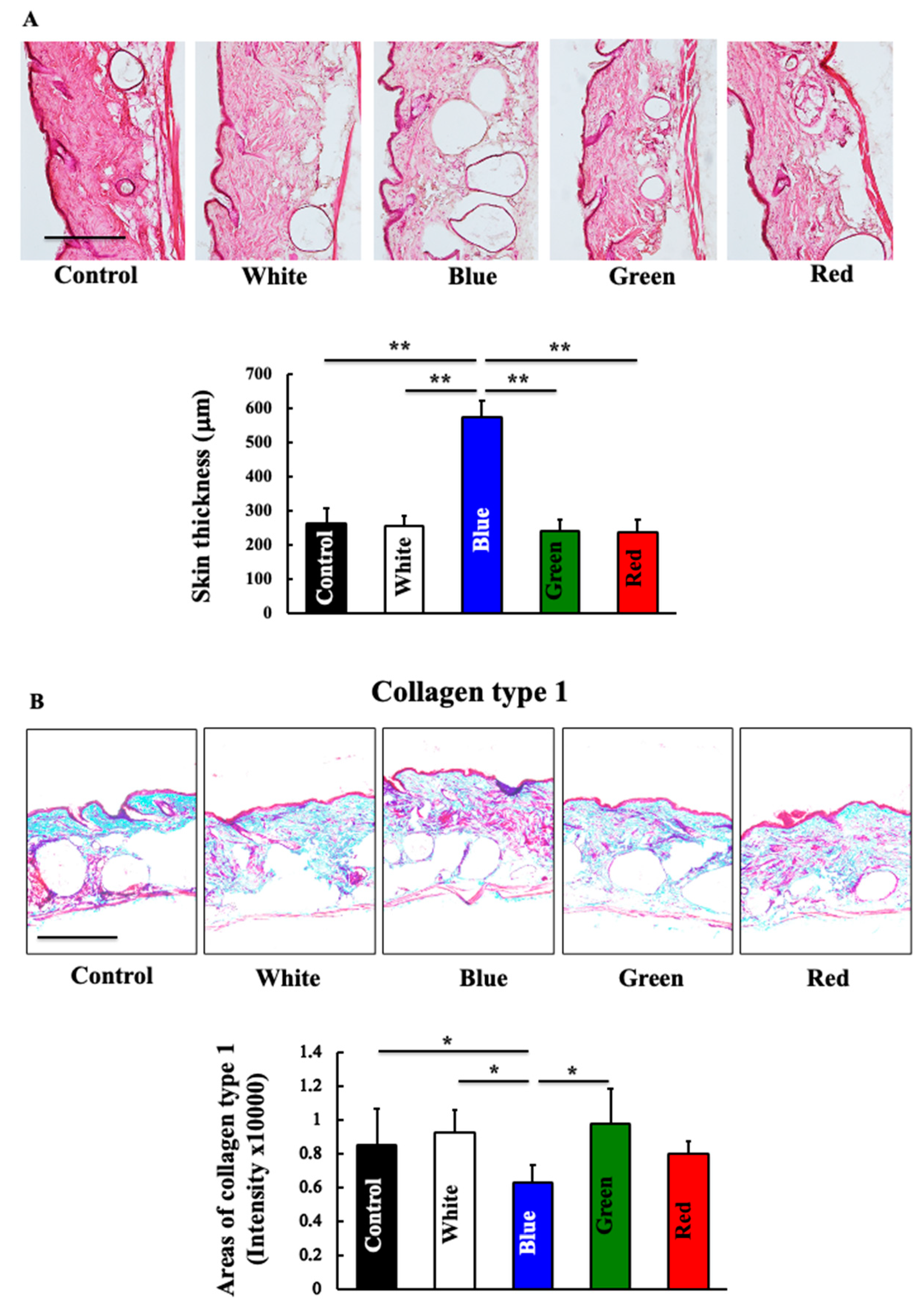
3.2. Effects of Blue Light on Total Skin Collagen
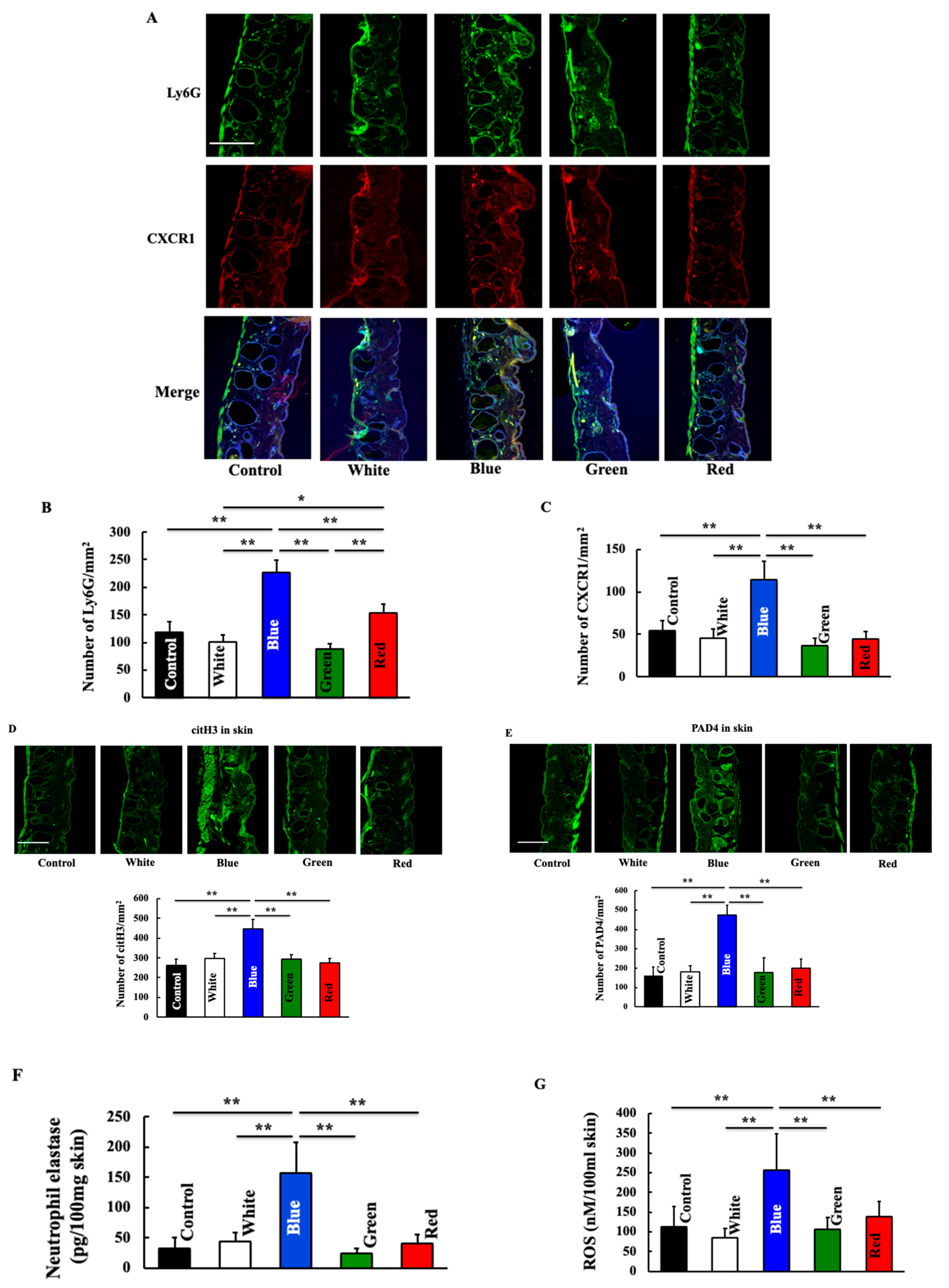
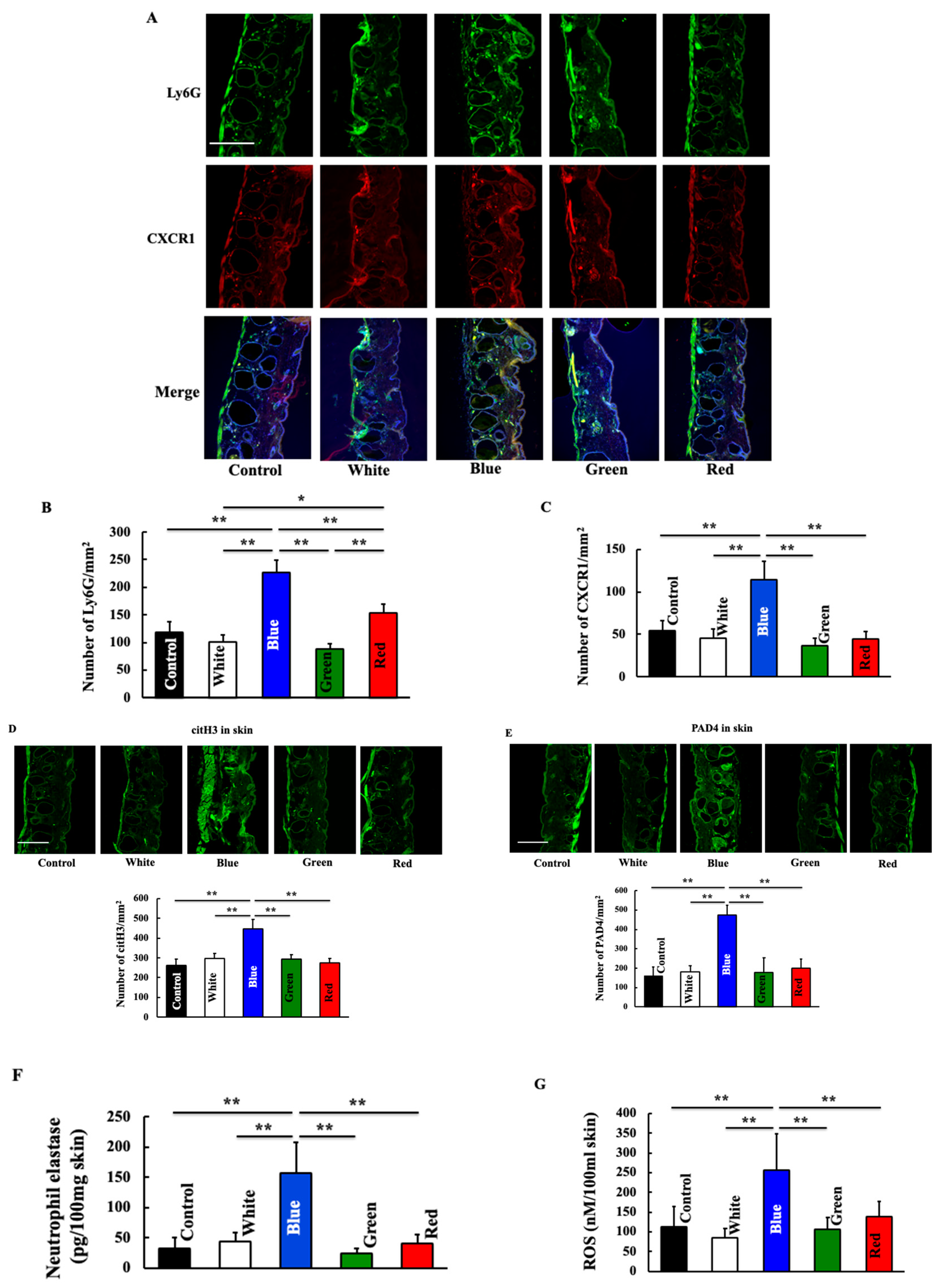
3.3. Effects of Blue Light on Expression of Neutrophil-Related Substances
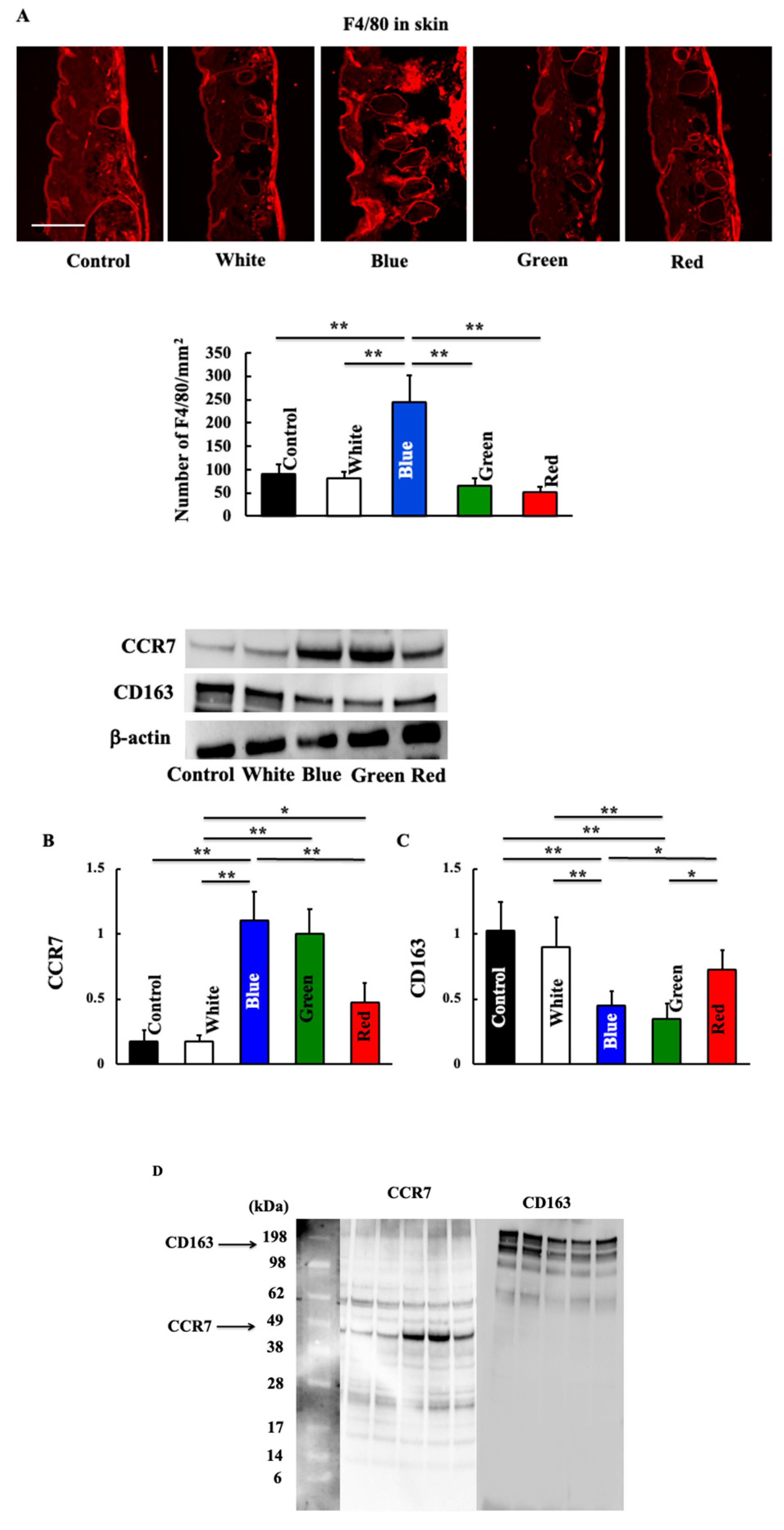
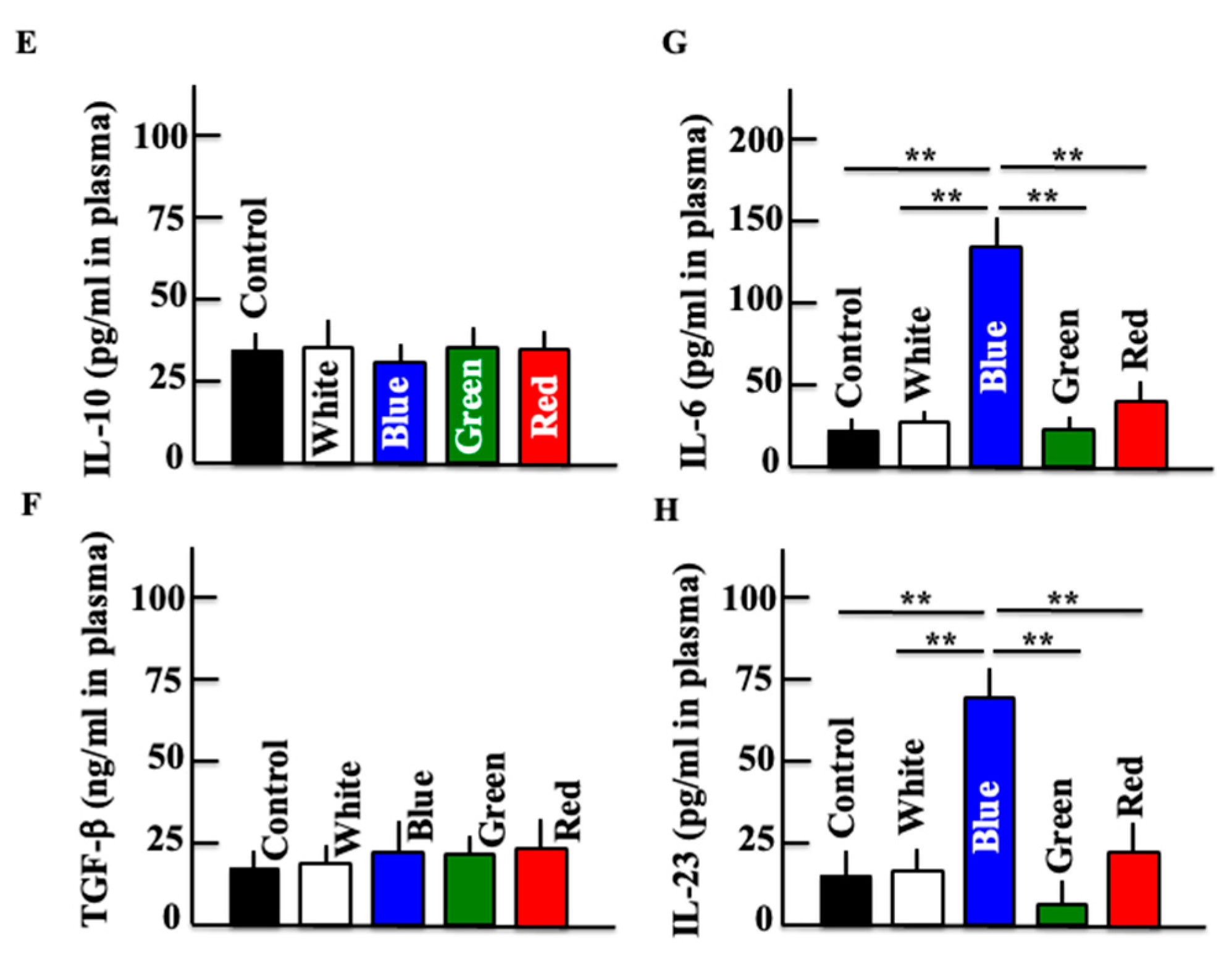
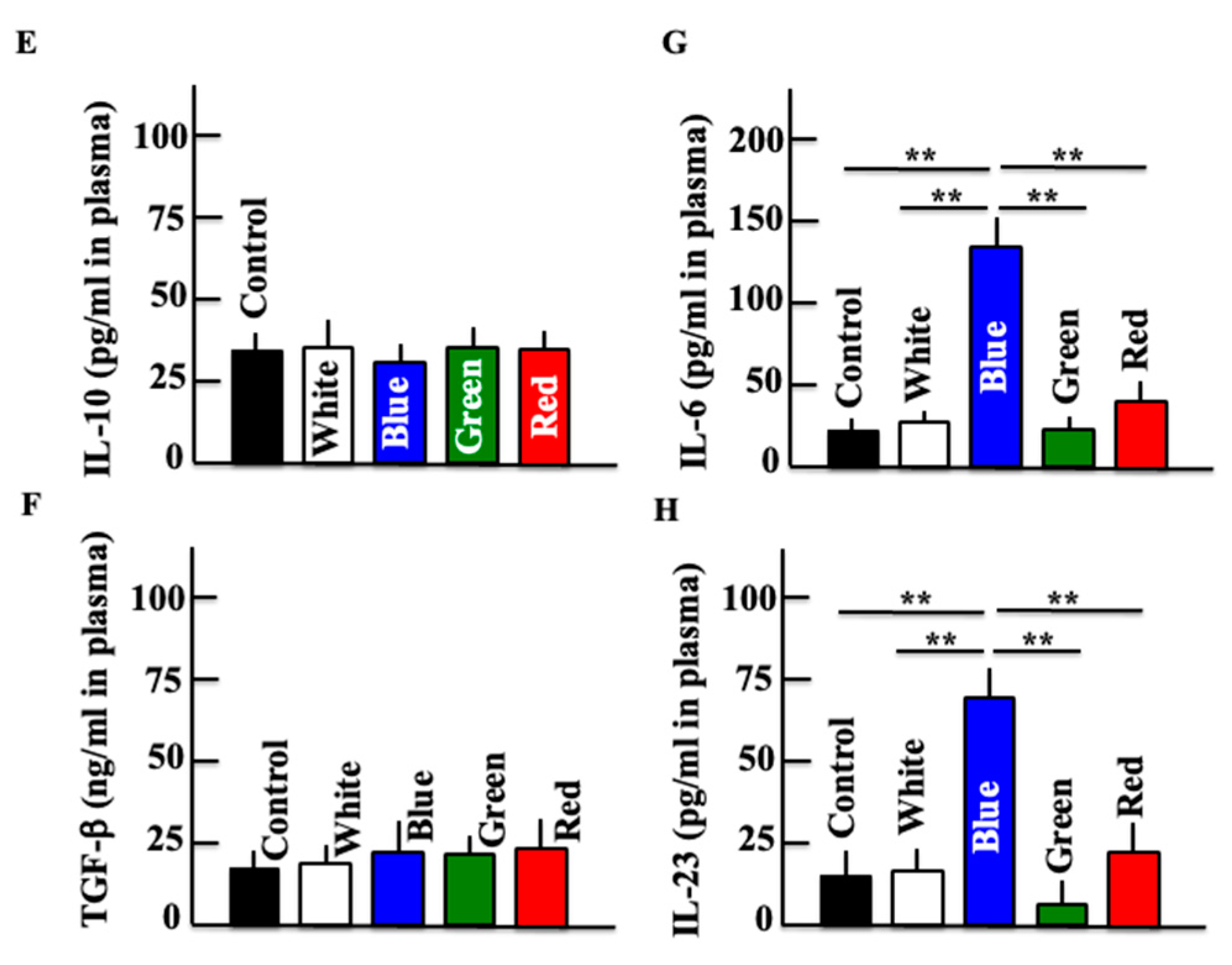
3.4. Effects of Blue Light on Expression of Macrophage-Related Substances
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ouyang, X.; Yang, J.; Hong, Z.; Wu, Y.; Xie, Y.; Wang, G. Mechanisms of blue light-induced eye hazard and protective measures: A review. Biomed. Pharmacother. 2020, 139, 110577. [Google Scholar] [CrossRef] [PubMed]
- Loh, K.; Redd, S. Understanding and preventing computer vision syndrome. Malays. Fam. Physician 2008, 3, 128–130. [Google Scholar] [PubMed]
- Jaiswal, S.; Asper, L.; Long, J.; Lee, A.; Harrison, K.; Gle-Biowski, B. Ocular and visual discomfort associated with smartphones, tablets and computers: What we do and do not know. Clin. Exp. Optom. 2019, 102, 463–477. [Google Scholar] [CrossRef] [PubMed]
- Tosini, G.; Ferguson, I.; Tsubota, K. Effects of blue light on the circadian system and eye physiology. Mol. Vis. 2016, 22, 61–72. [Google Scholar]
- Thresher, R.J.; Vitaterna, M.H.; Miyamoto, Y.; Kazantsev, A.; Hsu, D.S.; Petit, C.; Selby, C.P.; Dawut, L.; Smithies, O.; Takahashi, J.S.; et al. Role of mouse cryptochrome blue-light photoreceptor in circadian photoresponses. Science 1998, 282, 1490–1494. [Google Scholar] [CrossRef]
- Ozturk, N. Light-dependent reactions of animal circadian photoreceptor cryptochrome. FEBS J. 2022, 289, 6622–6639. [Google Scholar] [CrossRef]
- Lawrenson, J.G.; Hull, C.C.; Downie, L.E. The effect of blue-light blocking spectacle lenses on visual performance, macular health and the sleep-wake cycle: A systematic review of the literature. Ophthalmic Physiol. Opt. 2017, 37, 644–654. [Google Scholar] [CrossRef]
- Touitou, Y.; Touitou, D.; Reinberg, A. Disruption of adolescents’ circadian clock: The vicious circle of media use, exposure to light at night, sleep loss and risk behaviors. J. Physiol. Paris 2016, 110, 467–479. [Google Scholar] [CrossRef]
- Auffret, E.; Gomart, G.; Bourcier, T.; Gaucher, D.; Speeg-Schatz, C.; Sauer, A. Digital eye strain. Symptoms, prevalence, pathophysiology, and management. J. Fr. Ophtalmol. 2021, 44, 1605–1610. [Google Scholar] [CrossRef]
- Baez, G.; De la Vega, E.; Castro, C.; Elizarraras, R. Use of color lights for the detection of anomalies in quality systems. Work 2012, 41, 5889–5891. [Google Scholar] [CrossRef]
- Gallagher, R.P.; Lee, T.K. Adverse effects of ultraviolet radiation: A brief review. Prog. Biophys. Mol. Biol. 2006, 92, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Yannuzzi, L.A.; Fisher, Y.L.; Slakter, J.S.; Krueger, A. Solar retinopathy: A photobiologic and geophysical analysis. 1989. Retina 2012, 32, 28–43. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.C.; Zhou, Y.; Tan, G.; Li, J. Research progress about the effect and orevention of blue light on eyes. Int. J. Ophthalmol. 2018, 11, 1999–2003. [Google Scholar] [PubMed]
- Margrain, T.H.; Boulton, M.; Marshell, J.; Sliney, D.H. Do blue light filters confer protection against age-related macular degeneration? Prog. Retin. Eye Res. 2004, 23, 523–531. [Google Scholar] [CrossRef]
- Daneault, V.; Hebert, M.; Albouy, G.; Doyon, J.; Dumont, M.; Carrier, J.; Vandewalle, G. Aging reduces the stimulating effect of blue light on cognitive brain functions. Sleep 2014, 37, 85–96. [Google Scholar] [CrossRef]
- Avola, R.; Graziano, A.C.E.; Pannuzzo, G.; Bonina, F.; Cardile, V. Hydroxytyrosol from olive fruits prevents blue-light-induced damage in human keratinocytes and fibroblasts. J. Cell. Physiol. 2019, 34, 9065–9076. [Google Scholar] [CrossRef]
- Mann, T.; Eggers, K.; Rippke, F.; Tesch, M.; Buerger, A.; Darvin, M.E.; Schanzer, S.; Meinke, M.C.; Lademann, J.; Kolbe, L. High-energy visible light at ambient doses and intensities induces oxidative stress of skin-Protective effects of the antioxidant and Nrf2 inducer Licochalcone A in vitro and in vivo. Photodermatol. Photoimmunol. Photomed. 2020, 36, 135–144. [Google Scholar] [CrossRef]
- Austin, E.; Huang, A.; Adar, T.; Wang, E.; Jagdeo, J. Electronic device generated light increases reactive oxygen species in human fibroblasts. Lasers Surg. Med. 2018, 50, 689–695. [Google Scholar] [CrossRef]
- Schutz, R. Blue light and the skin. Curr. Probl. Dermatol. 2021, 55, 354–373. [Google Scholar]
- Sadowska, M.; Narbutt, J.; Lesiak, A. Blue light in dermatology. Life 2021, 11, 670. [Google Scholar] [CrossRef]
- Bekeredjian, R.; Walton, C.B.; MacCannell, K.A.; Ecker, J.; Kruse, F.; Outten, T.; Sutcliffe, D.; Gerard, R.D.; Bruick, R.K.; Shohet, R.V. Conditional HIF-lalpha expression produces a reversible cardiomyopathy. PLoS ONE 2010, 5, e11693. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, S.; Hiramoto, K.; Koyama, M.; Ooi, K. Skin disruption is associated with indomethacine-induced small intestine injury in mice. Exp. Dermatol. 2014, 23, 659–663. [Google Scholar] [CrossRef] [PubMed]
- Hiramoto, K.; Sugiyama, D.; Takahashi, Y.; Mafune, E. The amelioration effect of tranexamic acid in wrinkles induced by skin dryness. Biomed. Pharmacother. 2016, 80, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Masaki, H. Skin aging mechanisms and materials. J. Cos. Sci. Soc. 2013, 37, 11–16. [Google Scholar]
- Nathan, C. Neutrophils and immunity: Challenges and opportunities. Nat. Rev. Immunol. 2006, 6, 173–182. [Google Scholar] [CrossRef]
- Brinkmann, V.; Reichard, V.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef]
- Fuchs, T.A.; Abed, V.; Goosmann, C.; Hurwitz, R.; Schulze, I.; Wahn, V.; Weinrauch, Y.; Brinkmann, V.; Zychlinsky, A. Novel cell death program leads to neutrophil extracellular traps. J. Cell Biol. 2007, 176, 231–241. [Google Scholar] [CrossRef]
- Fousert, E.; Toes, R.; Desai, J. Neutrophil extracellulars traps (NETs) take the central stage in driving autoimmune responses. Cells 2020, 9, 915. [Google Scholar] [CrossRef]
- Jin, W.; Yin, H.; Li, H.; Yu, X.J.; Xu, H.X.; Liu, L. Neutrophil extracellular DNA traps promote pancreatic cancer cells migration and invasion by activating EGFR/ERK pathway. J. Cell. Mol. Med. 2012, 25, 5443–5456. [Google Scholar] [CrossRef]
- Zhu, T.; Zou, X.; Yang, C.; Li, L.; Wang, B.; Li, R.; Li, H.; Xu, Z.; Huang, D.; Wu, Q. Neutrophil extracellular traps promote gastric cancer metastasis by inducing epithelial-mesenchymal transition. Int. J. Mol. Med. 2021, 48, 127. [Google Scholar] [CrossRef]
- Albrengues, J.; Shields, M.A.; Ng, D.; Park, C.G.; Ambrico, A.; Poindexter, M.E.; Upadhyay, P.; Uyeminami, D.L.; Pommier, A.; Kuttner, V.; et al. Neutrophil extracellular traps produced during inflammation awaken dormant cancer cells in mice. Science 2018, 361, eaao4227. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Zhou, Y.; Bu, H.; Lv, T.; Shi, Y.; Yang, J. Deletion of interleukin-6 in monocytes/macrophages suppresses the initiation of hepatocellular carcinoma in mice. J. Exp. Clin. Cancer Res. 2016, 35, 131. [Google Scholar] [CrossRef] [PubMed]
- Crivennikov, S.I.; Wang, K.; Mucida, D.; Stewart, C.A.; Schnabl, B.; Jauch, D.; Taniguchi, K.; Yu, G.Y.; Osterreicher, C.H.; Hung, K.E.; et al. Adenoma-linked barrier defects and microbial products drive IL-23/IL-17-mediated tumor growth. Nature 2012, 491, 254–258. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.H.; Britton, G.J.; Hill, E.V.; Verhagen, J.; Burton, B.R.; Wraith, D.C. Regulation of adaptive immunity: The role of imterleukin-10. Front. Immunol. 2013, 4, 129. [Google Scholar] [CrossRef]
- Savage, N.D.L.; de Boer, T.; Walburg, K.V.; Joosten, S.A.; van Meijgaarden, K.; Geluk, A.; Ottenhoff, T.H.M. Human anti-inflammatory macrophages induced Foxp3+ GITR+ CD25+ regulatory T cells, which suppress via membrane-bound TGF beta-1. J. Immunol. 2008, 181, 2220–2226. [Google Scholar] [CrossRef]
- Li, C.; Shintani, S.; Terakado, N.; Nakashiro, K.; Hamakawa, H. Infiltration of tumor-associated macrophages in human oral squamous cell carcinoma. Oncol. Rep. 2002, 9, 1219–1223. [Google Scholar] [CrossRef]
- Takanami, I.; Takeuchi, K.; Kodaira, S. Tumor-associated macrophage infiltration in pulmonary adenocarcinoma: Association with angiogenesis and poor prognosis. Oncology 1999, 57, 138–142. [Google Scholar] [CrossRef]
- Ge, G.; Wang, Y.; Xu, Y.; Pu, W.; Tan, Y.; Liu, P.; Ding, H.; Lu, Y.M.; Wang, J.; Liu, W.; et al. Induced skin aging by blue-light irradiation in human skin fibroblasts via TGF-β, JNK and EGFR pathways. J. Dermatol. Sci. 2023, 28. [Google Scholar] [CrossRef]
- Vignais, P.V. The superoxide-generating NADPH oxidase. Cell. Mol. Life Sci. CMLS 2002, 59, 1428–1459. [Google Scholar] [CrossRef]
- Miyata, K.; Tamura, M.; Sumimoto, H. Molecular mechanism for production of reactive oxygen species by neutrophils. Inflamm. Regen. 2023, 25, 113–117. [Google Scholar]
- Epelman, S.; Lavine, K.J.; Randolph, G.J. Origin and functions of tissue macrophages. Immunity 2014, 41, 21–35. [Google Scholar] [CrossRef] [PubMed]
- Billings, E.A.; Lee, C.S.; Owen, K.A.; D’Souza, R.S.; Ravichandran, K.S.; Casanova, J.E. The adhesion GRCR BAI1 mediates macrophage ROS production and microbicidal activity against Gram-negative bacteria. Sci. Signal. 2016, 9, ra14. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hiramoto, K.; Kubo, S.; Tsuji, K.; Sugiyama, D.; Hamano, H. Induction of Skin Cancer by Long-Term Blue Light Irradiation. Biomedicines 2023, 11, 2321. https://doi.org/10.3390/biomedicines11082321
Hiramoto K, Kubo S, Tsuji K, Sugiyama D, Hamano H. Induction of Skin Cancer by Long-Term Blue Light Irradiation. Biomedicines. 2023; 11(8):2321. https://doi.org/10.3390/biomedicines11082321
Chicago/Turabian StyleHiramoto, Keiichi, Sayaka Kubo, Keiko Tsuji, Daijiro Sugiyama, and Hideo Hamano. 2023. "Induction of Skin Cancer by Long-Term Blue Light Irradiation" Biomedicines 11, no. 8: 2321. https://doi.org/10.3390/biomedicines11082321
APA StyleHiramoto, K., Kubo, S., Tsuji, K., Sugiyama, D., & Hamano, H. (2023). Induction of Skin Cancer by Long-Term Blue Light Irradiation. Biomedicines, 11(8), 2321. https://doi.org/10.3390/biomedicines11082321