
Kinin B1 Receptor Antagonism Prevents Acute Kidney Injury to Chronic Kidney Disease Transition in Renal Ischemia-Reperfusion by Increasing the M2 Macrophages Population in C57BL6J Mice

 ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Ischemia-Reperfusion
2.3. R715 Treatment
2.4. Blood Sampling and Tissue Collection
2.5. Renal Function
2.6. Real-Time PCR
2.7. Tubular Injury and Renal Fibrosis Analysis
2.8. Statistical Analysis
3. Results
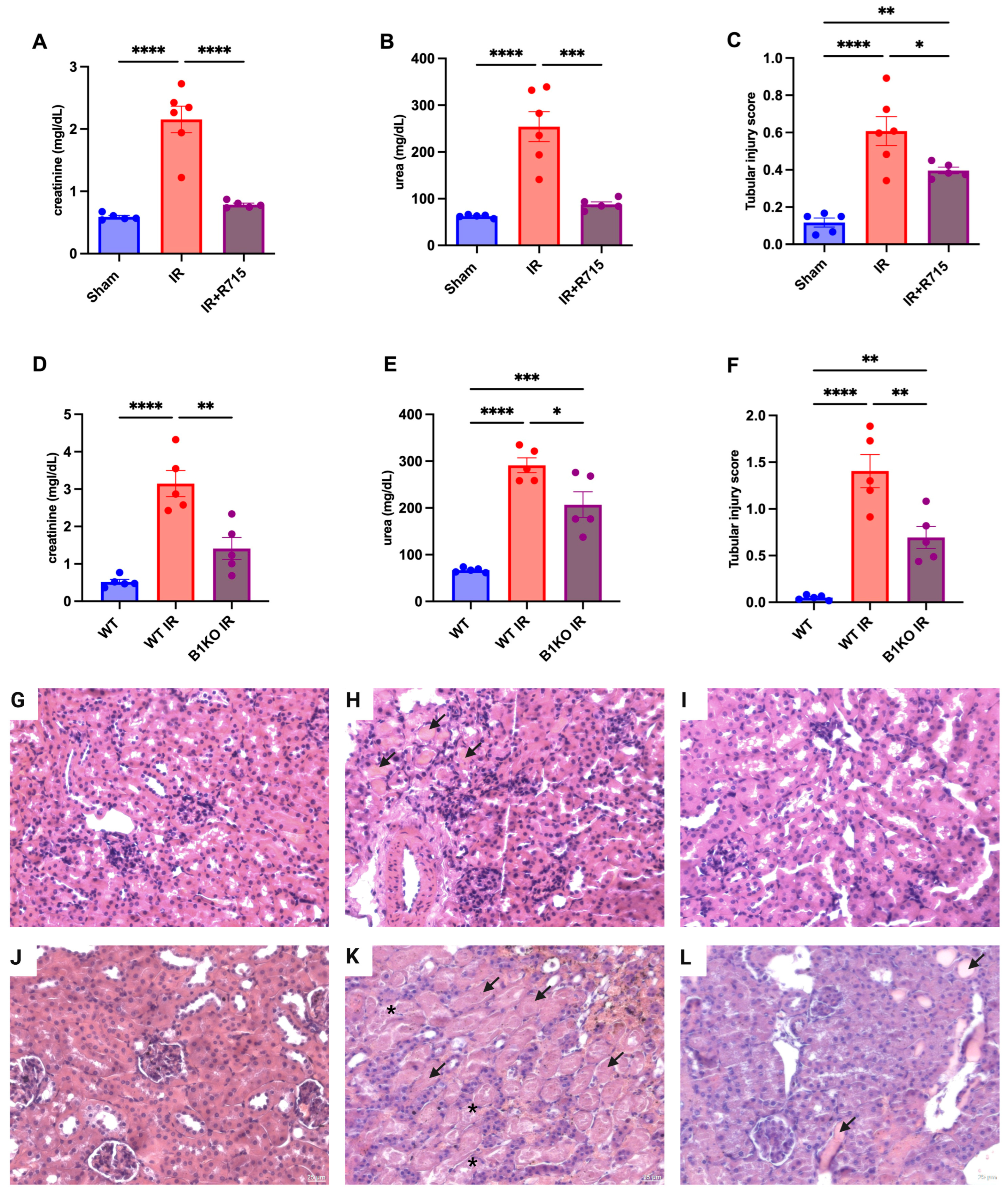
3.1. B1R Absence and B1R Antagonism (R715) Preserves Kidney Function after Acute Kidney Injury
3.2. B1R Absence and B1R Antagonism Preserve Kidney Function in Chronic Disease Kidney Model
3.3. B1R Antagonism Ameliorates Resolution Kidney Inflammatory Process through Macrophage M2 Polarization Increase and Macrophage M1 Decrease
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- GBD Chronic Kidney Disease Collaboration. Global, regional, and national burden of chronic kidney disease, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2020, 395, 709–733. [Google Scholar] [CrossRef] [PubMed]
- Klein, J.; Gonzalez, J.; Decramer, S.; Bandin, F.; Neau, E.; Salant, D.J.; Heeringa, P.; Pesquero, J.B.; Schanstra, J.P.; Bascands, J.L. Blockade of the kinin B1 receptor ameloriates glomerulonephritis. J. Am. Soc. Nephrol. 2010, 21, 1157–1164. [Google Scholar] [CrossRef] [PubMed]
- Marceau, F.; Hess, J.F.; Bachvarov, D.R. The B1 receptors for kinins. Pharmacol. Rev. 1998, 50, 357–386. [Google Scholar] [PubMed]
- Araujo, R.C.; Kettritz, R.; Fichtner, I.; Paiva, A.C.; Pesquero, J.B.; Bader, M. Altered neutrophil homeostasis in kinin B1 receptor-deficient mice. Biol. Chem. 2001, 382, 91–95. [Google Scholar] [CrossRef]
- Pesquero, J.B.; Araujo, R.C.; Heppenstall, P.A.; Stucky, C.L.; Silva, J.A., Jr.; Walther, T.; Oliveira, S.M.; Pesquero, J.L.; Paiva, A.C.; Calixto, J.B.; et al. Hypoalgesia and altered inflammatory responses in mice lacking kinin B1 receptors. Proc. Natl. Acad. Sci. USA 2000, 97, 8140–8145. [Google Scholar] [CrossRef]
- Schanstra, J.P.; Bataille, E.; Marin Castano, M.E.; Barascud, Y.; Hirtz, C.; Pesquero, J.B.; Pecher, C.; Gauthier, F.; Girolami, J.P.; Bascands, J.L. The B1-agonist [des-Arg10]-kallidin activates transcription factor NF-kappaB and induces homologous upregulation of the bradykinin B1-receptor in cultured human lung fibroblasts. J. Clin. Investig. 1998, 101, 2080–2091. [Google Scholar] [CrossRef][Green Version]
- Kahn, R.; Mossberg, M.; Stahl, A.L.; Johansson, K.; Lopatko Lindman, I.; Heijl, C.; Segelmark, M.; Morgelin, M.; Leeb-Lundberg, L.M.; Karpman, D. Microvesicle transfer of kinin B1-receptors is a novel inflammatory mechanism in vasculitis. Kidney Int. 2017, 91, 96–105. [Google Scholar] [CrossRef]
- Wang, P.H.; Cenedeze, M.A.; Campanholle, G.; Malheiros, D.M.; Torres, H.A.; Pesquero, J.B.; Pacheco-Silva, A.; Camara, N.O. Deletion of bradykinin B1 receptor reduces renal fibrosis. Int. Immunopharmacol. 2009, 9, 653–657. [Google Scholar] [CrossRef]
- Estrela, G.R.; Wasinski, F.; Almeida, D.C.; Amano, M.T.; Castoldi, A.; Dias, C.C.; Malheiros, D.M.; Almeida, S.S.; Paredes-Gamero, E.J.; Pesquero, J.B.; et al. Kinin B1 receptor deficiency attenuates cisplatin-induced acute kidney injury by modulating immune cell migration. J. Mol. Med. 2014, 92, 399–409. [Google Scholar] [CrossRef]
- Estrela, G.R.; Wasinski, F.; Felizardo, R.J.F.; Souza, L.L.; Camara, N.O.S.; Bader, M.; Araujo, R.C. MATE-1 modulation by kinin B1 receptor enhances cisplatin efflux from renal cells. Mol. Cell Biochem. 2017, 428, 101–108. [Google Scholar] [CrossRef]
- Sun, P.P.; Zhou, X.J.; Su, J.Q.; Wang, C.; Yu, X.J.; Su, T.; Liu, G.; Wang, S.X.; Nie, J.; Yang, L. Urine macrophages reflect kidney macrophage content during acute tubular interstitial and glomerular injury. Clin. Immunol. 2019, 205, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Lu, X.; Ren, J.; Privratsky, J.R.; Yang, B.; Rudemiller, N.P.; Zhang, J.; Griffiths, R.; Jain, M.K.; Nedospasov, S.A.; et al. KLF4 in Macrophages Attenuates TNFalpha-Mediated Kidney Injury and Fibrosis. J. Am. Soc. Nephrol. 2019, 30, 1925–1938. [Google Scholar] [CrossRef] [PubMed]
- Barrera-Chimal, J.; Estrela, G.R.; Lechner, S.M.; Giraud, S.; El Moghrabi, S.; Kaaki, S.; Kolkhof, P.; Hauet, T.; Jaisser, F. The myeloid mineralocorticoid receptor controls inflammatory and fibrotic responses after renal injury via macrophage interleukin-4 receptor signaling. Kidney Int. 2018, 93, 1344–1355. [Google Scholar] [CrossRef]
- Zhang, M.Z.; Yao, B.; Yang, S.; Jiang, L.; Wang, S.; Fan, X.; Yin, H.; Wong, K.; Miyazawa, T.; Chen, J.; et al. CSF-1 signaling mediates recovery from acute kidney injury. J. Clin. Investig. 2012, 122, 4519–4532. [Google Scholar] [CrossRef]
- Ranganathan, P.V.; Jayakumar, C.; Ramesh, G. Netrin-1-treated macrophages protect the kidney against ischemia-reperfusion injury and suppress inflammation by inducing M2 polarization. Am. J. Physiol. Renal Physiol. 2013, 304, F948–F957. [Google Scholar] [CrossRef]
- Xie, X.; Yang, X.; Wu, J.; Ma, J.; Wei, W.; Fei, X.; Wang, M. Trib1 Contributes to Recovery From Ischemia/Reperfusion-Induced Acute Kidney Injury by Regulating the Polarization of Renal Macrophages. Front. Immunol. 2020, 11, 473. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Zhang, Q.; Wen, J.; Chen, T.; He, L.; Wang, Y.; Yin, J.; Wu, R.; Xue, R.; Li, S.; et al. Ischemic Duration and Frequency Determines AKI-to-CKD Progression Monitored by Dynamic Changes of Tubular Biomarkers in IRI Mice. Front. Physiol. 2019, 10, 153. [Google Scholar] [CrossRef]
- Estrela, G.R.; Freitas-Lima, L.C.; Budu, A.; Arruda, A.C.; Perilhao, M.S.; Fock, R.A.; Barrera-Chimal, J.; Araujo, R.C. Chronic Kidney Disease Induced by Cisplatin, Folic Acid and Renal Ischemia Reperfusion Induces Anemia and Promotes GATA-2 Activation in Mice. Biomedicines 2021, 9, 769. [Google Scholar] [CrossRef]
- Li, C.; Ding, X.Y.; Xiang, D.M.; Xu, J.; Huang, X.L.; Hou, F.F.; Zhou, Q.G. Enhanced M1 and Impaired M2 Macrophage Polarization and Reduced Mitochondrial Biogenesis via Inhibition of AMP Kinase in Chronic Kidney Disease. Cell Physiol. Biochem. 2015, 36, 358–372. [Google Scholar] [CrossRef]
- Lv, L.L.; Tang, P.M.; Li, C.J.; You, Y.K.; Li, J.; Huang, X.R.; Ni, J.; Feng, M.; Liu, B.C.; Lan, H.Y. The pattern recognition receptor, Mincle, is essential for maintaining the M1 macrophage phenotype in acute renal inflammation. Kidney Int. 2017, 91, 587–602. [Google Scholar] [CrossRef]
- Lee, S.; Huen, S.; Nishio, H.; Nishio, S.; Lee, H.K.; Choi, B.S.; Ruhrberg, C.; Cantley, L.G. Distinct macrophage phenotypes contribute to kidney injury and repair. J. Am. Soc. Nephrol. 2011, 22, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Morii, T.; Fujita, H.; Narita, T.; Shimotomai, T.; Fujishima, H.; Yoshioka, N.; Imai, H.; Kakei, M.; Ito, S. Association of monocyte chemoattractant protein-1 with renal tubular damage in diabetic nephropathy. J. Diabetes Complicat. 2003, 17, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Doi, K.; Maeda-Mamiya, R.; Negishi, K.; Portilla, D.; Sugaya, T.; Fujita, T.; Noiri, E. Urinary L-type fatty acid-binding protein can reflect renal tubulointerstitial injury. Am. J. Pathol. 2009, 174, 1203–1211. [Google Scholar] [CrossRef]
- Zhang, H.; Sun, S.C. NF-kappaB in inflammation and renal diseases. Cell Biosci. 2015, 5, 63. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.G.; Kim, S.C.; Ko, Y.S.; Lee, H.Y.; Jo, S.K.; Cho, W. The Role of M2 Macrophages in the Progression of Chronic Kidney Disease following Acute Kidney Injury. PLoS ONE 2015, 10, e0143961. [Google Scholar] [CrossRef]
- Welch, J.S.; Escoubet-Lozach, L.; Sykes, D.B.; Liddiard, K.; Greaves, D.R.; Glass, C.K. TH2 cytokines and allergic challenge induce Ym1 expression in macrophages by a STAT6-dependent mechanism. J. Biol. Chem. 2002, 277, 42821–42829. [Google Scholar] [CrossRef]
- Cavalcante, P.A.M.; Alenina, N.; Budu, A.; Freitas-Lima, L.C.; Alves-Silva, T.; Agudelo, J.S.H.; Qadri, F.; Camara, N.O.S.; Bader, M.; Araujo, R.C. Nephropathy in Hypertensive Animals Is Linked to M2 Macrophages and Increased Expression of the YM1/Chi3l3 Protein. Mediat. Inflamm. 2019, 2019, 9086758. [Google Scholar] [CrossRef]
- Lu, J.; Cao, Q.; Zheng, D.; Sun, Y.; Wang, C.; Yu, X.; Wang, Y.; Lee, V.W.; Zheng, G.; Tan, T.K.; et al. Discrete functions of M2a and M2c macrophage subsets determine their relative efficacy in treating chronic kidney disease. Kidney Int. 2013, 84, 745–755. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Y.P.; Zheng, G.; Lee, V.W.; Ouyang, L.; Chang, D.H.; Mahajan, D.; Coombs, J.; Wang, Y.M.; Alexander, S.I.; et al. Ex vivo programmed macrophages ameliorate experimental chronic inflammatory renal disease. Kidney Int. 2007, 72, 290–299. [Google Scholar] [CrossRef]
- Feng, Y.; Ren, J.; Gui, Y.; Wei, W.; Shu, B.; Lu, Q.; Xue, X.; Sun, X.; He, W.; Yang, J.; et al. Wnt/beta-Catenin-Promoted Macrophage Alternative Activation Contributes to Kidney Fibrosis. J. Am. Soc. Nephrol. 2018, 29, 182–193. [Google Scholar] [CrossRef]
- Liu, B.; Jiang, J.; Liang, H.; Xiao, P.; Lai, X.; Nie, J.; Yu, W.; Gao, Y.; Wen, S. Natural killer T cell/IL-4 signaling promotes bone marrow-derived fibroblast activation and M2 macrophage-to-myofibroblast transition in renal fibrosis. Int. Immunopharmacol. 2021, 98, 107907. [Google Scholar] [CrossRef] [PubMed]
- Pan, B.; Liu, G.; Jiang, Z.; Zheng, D. Regulation of renal fibrosis by macrophage polarization. Cell Physiol. Biochem. 2015, 35, 1062–1069. [Google Scholar] [CrossRef] [PubMed]
- Yurdagul, A., Jr.; Subramanian, M.; Wang, X.; Crown, S.B.; Ilkayeva, O.R.; Darville, L.; Kolluru, G.K.; Rymond, C.C.; Gerlach, B.D.; Zheng, Z.; et al. Macrophage Metabolism of Apoptotic Cell-Derived Arginine Promotes Continual Efferocytosis and Resolution of Injury. Cell Metab. 2020, 31, 518–533.e510. [Google Scholar] [CrossRef] [PubMed]
- Pesce, J.T.; Ramalingam, T.R.; Mentink-Kane, M.M.; Wilson, M.S.; El Kasmi, K.C.; Smith, A.M.; Thompson, R.W.; Cheever, A.W.; Murray, P.J.; Wynn, T.A. Arginase-1-expressing macrophages suppress Th2 cytokine-driven inflammation and fibrosis. PLoS Pathog. 2009, 5, e1000371. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Gene | Forward 5′-3′ | Reverse 5′-3′ |
|---|---|---|
| 18S | CGC CGC TAG AGG TGA AAT TC | TCT TGG CAA ATG CTT TCG C |
| β-actin | CTG GCC TCA CTG TCC ACC TT | CGG ACT CAT CGT ACT CCT GCT T |
| TNF-α | GCC TCT TCT CAT TCC TGC TTG | CTG ATG AGA GGG AGG CCA TT |
| IL-1b | AGGAGAACCAAGCAACGACA | CGTTTTTCCATCTTCTTCTTTG |
| IL-4R | CAC AGT GCA CGA AAG CTG AA | ATG GGC ACA AGC TGT GGT AG |
| YM1 | CCC CTG GAC ATG GAT GAC TT | AGC TCC TCT CAA TAA GGG CC |
| IL6 | TAGTCCTTCCTACCCCAATTTCC | TTGGTCCTTAGCCACTCCTCC |
| ARG | CGC-CTT-TCT-CAA-AAG-GAC-AG | CCA-GCT-CTT-CAT-TGG-CTT-TC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Estrela, G.R.; Santos, R.B.; Budu, A.; de Arruda, A.C.; Barrera-Chimal, J.; Araújo, R.C. Kinin B1 Receptor Antagonism Prevents Acute Kidney Injury to Chronic Kidney Disease Transition in Renal Ischemia-Reperfusion by Increasing the M2 Macrophages Population in C57BL6J Mice. Biomedicines 2023, 11, 2194. https://doi.org/10.3390/biomedicines11082194
Estrela GR, Santos RB, Budu A, de Arruda AC, Barrera-Chimal J, Araújo RC. Kinin B1 Receptor Antagonism Prevents Acute Kidney Injury to Chronic Kidney Disease Transition in Renal Ischemia-Reperfusion by Increasing the M2 Macrophages Population in C57BL6J Mice. Biomedicines. 2023; 11(8):2194. https://doi.org/10.3390/biomedicines11082194
Chicago/Turabian StyleEstrela, Gabriel Rufino, Raisa Brito Santos, Alexandre Budu, Adriano Cleis de Arruda, Jonatan Barrera-Chimal, and Ronaldo Carvalho Araújo. 2023. "Kinin B1 Receptor Antagonism Prevents Acute Kidney Injury to Chronic Kidney Disease Transition in Renal Ischemia-Reperfusion by Increasing the M2 Macrophages Population in C57BL6J Mice" Biomedicines 11, no. 8: 2194. https://doi.org/10.3390/biomedicines11082194
APA StyleEstrela, G. R., Santos, R. B., Budu, A., de Arruda, A. C., Barrera-Chimal, J., & Araújo, R. C. (2023). Kinin B1 Receptor Antagonism Prevents Acute Kidney Injury to Chronic Kidney Disease Transition in Renal Ischemia-Reperfusion by Increasing the M2 Macrophages Population in C57BL6J Mice. Biomedicines, 11(8), 2194. https://doi.org/10.3390/biomedicines11082194