
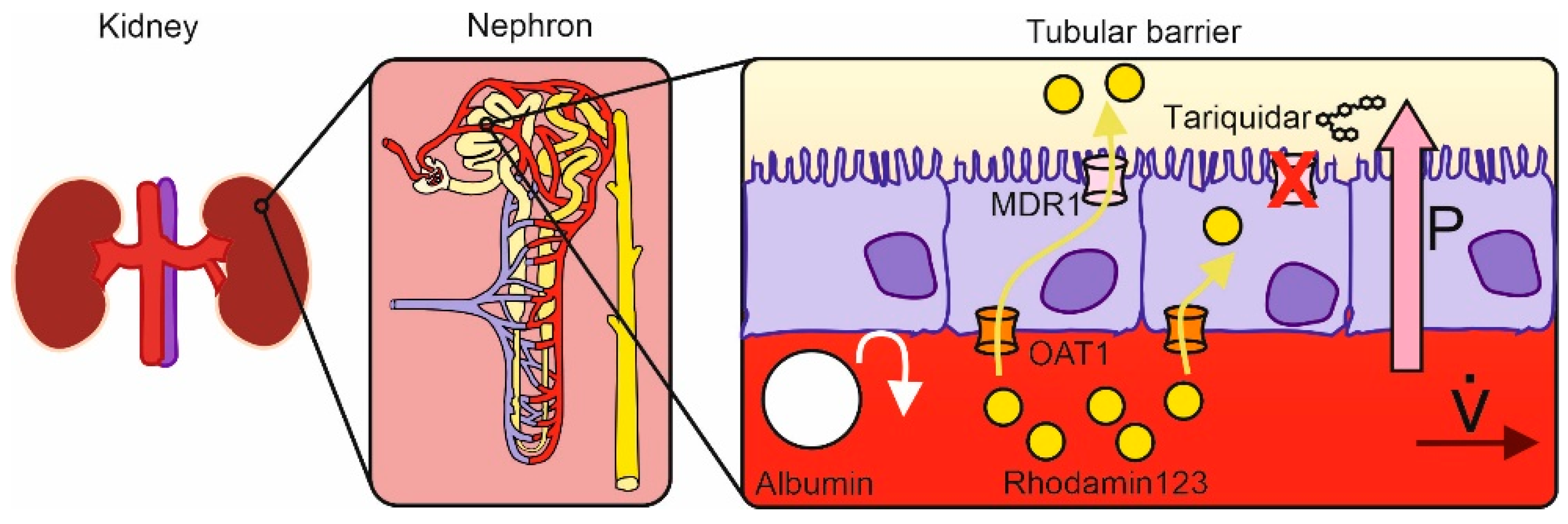
Microphysiological Conditions Do Not Affect MDR1-Mediated Transport of Rhodamine 123 above an Artificial Proximal Tubule

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture (Barrier Formation)
2.2. Transport Assay
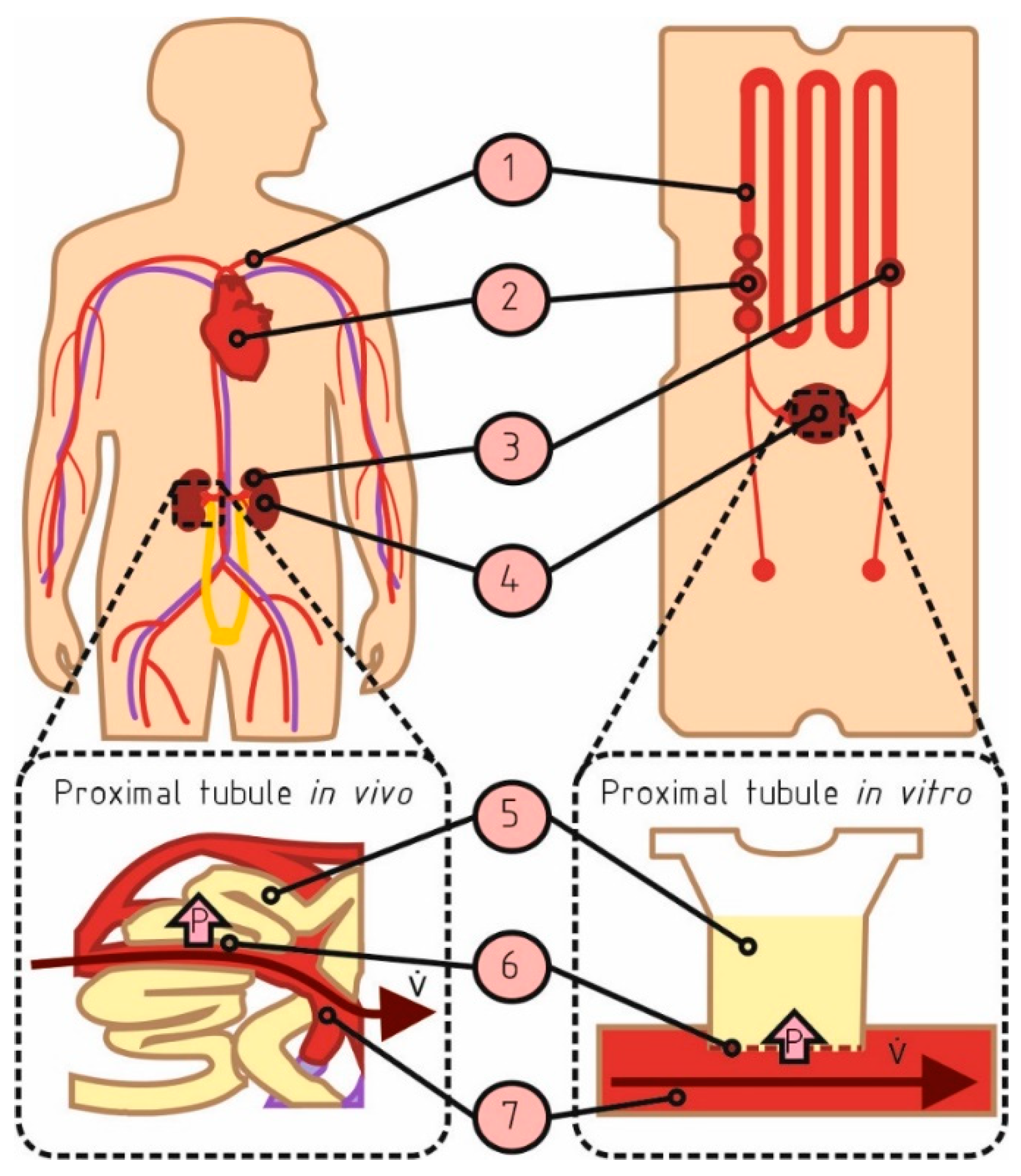
2.3. Adoption of the Microphysiological System
2.3.1. Microphysiological Flow
2.3.2. Microphysiological Pressure
- Pcap—hydrodynamic pressure of the peritubular capillary;
- Ptub—hydrodynamic pressure of the proximal tubular lumen;
- πcap—colloid osmotic pressure of the peritubular capillary;
- πbow—colloid osmotic pressure of proximal tubular lumen;
- ΔPint—pressure drop within the tubular interstitium.
2.4. Fluorescence Microscopy
2.5. Quantitative Measurement of MDR1 Expression
2.6. Statistical Analysis
3. Results
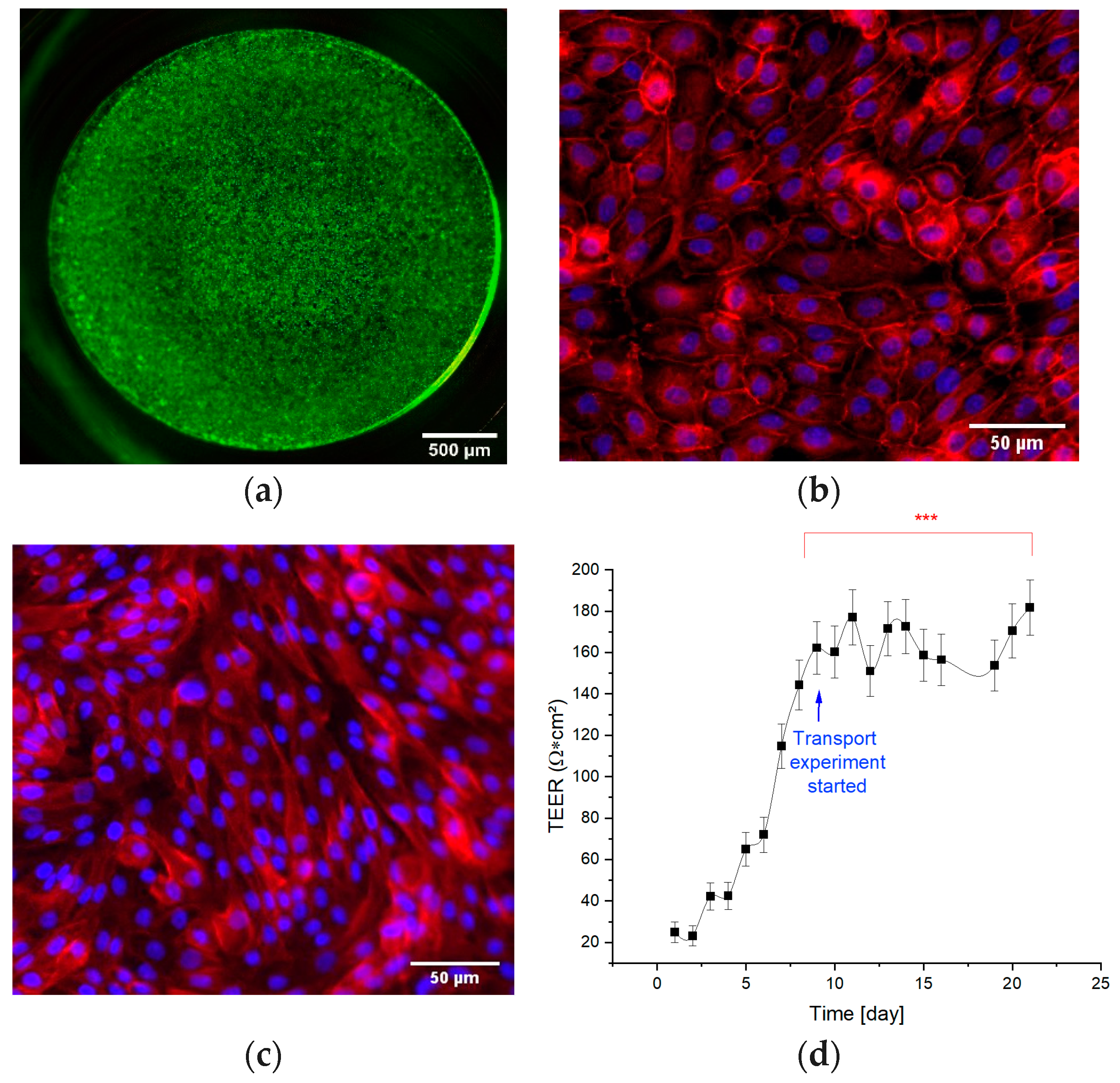
3.1. Barrier Integrity of the Tubular Barrier
3.2. Dynamic Cultivation
3.3. Time-Resolved Fluorescence Measurements of R123 Transport
3.4. Permeability of the Barrier
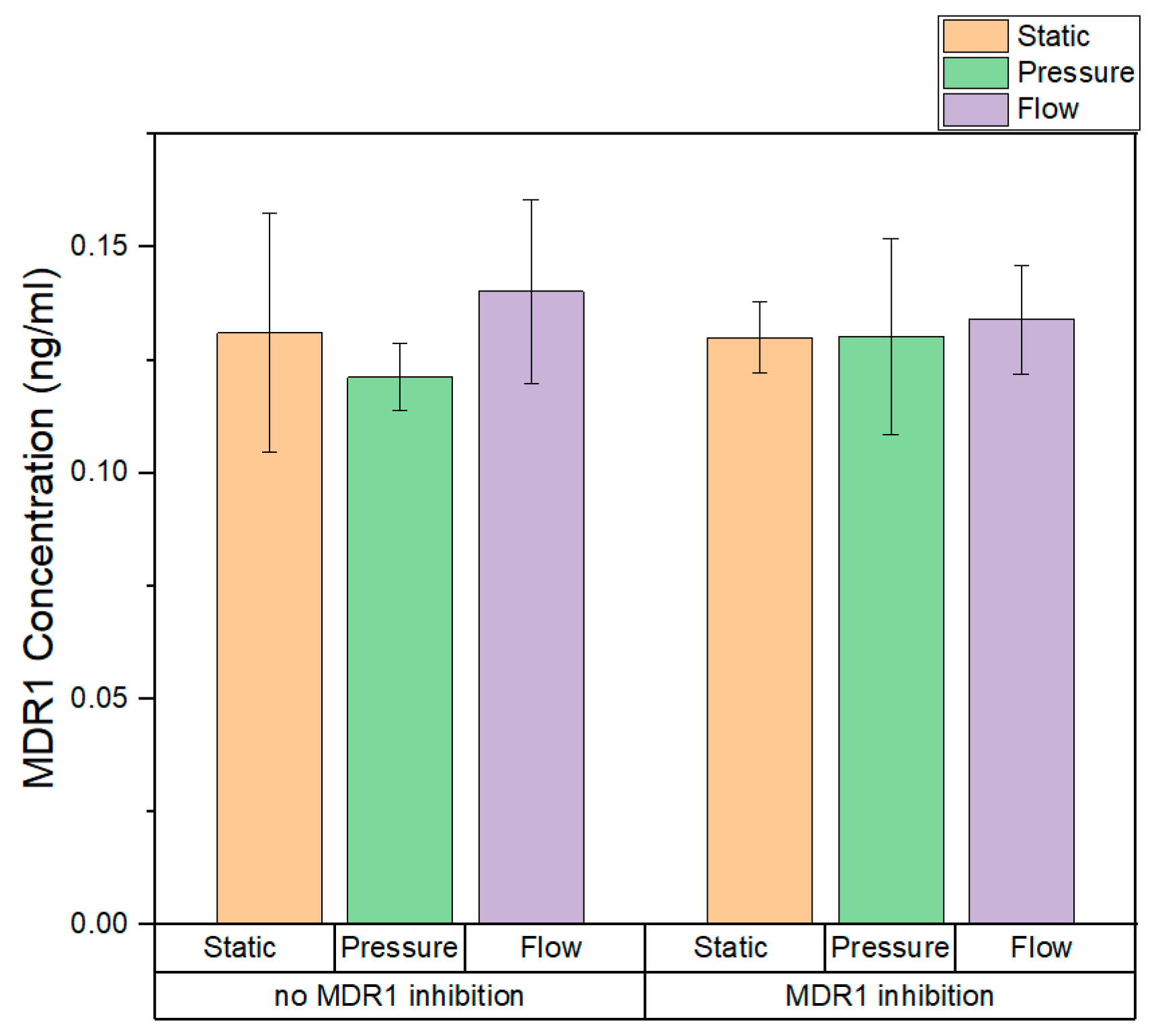
3.5. Quantitative Measurment of MDR1 Expression in Static and Dynamic Conditions
4. Discussion
4.1. Kinetic Characterization of Rhodamine Transport
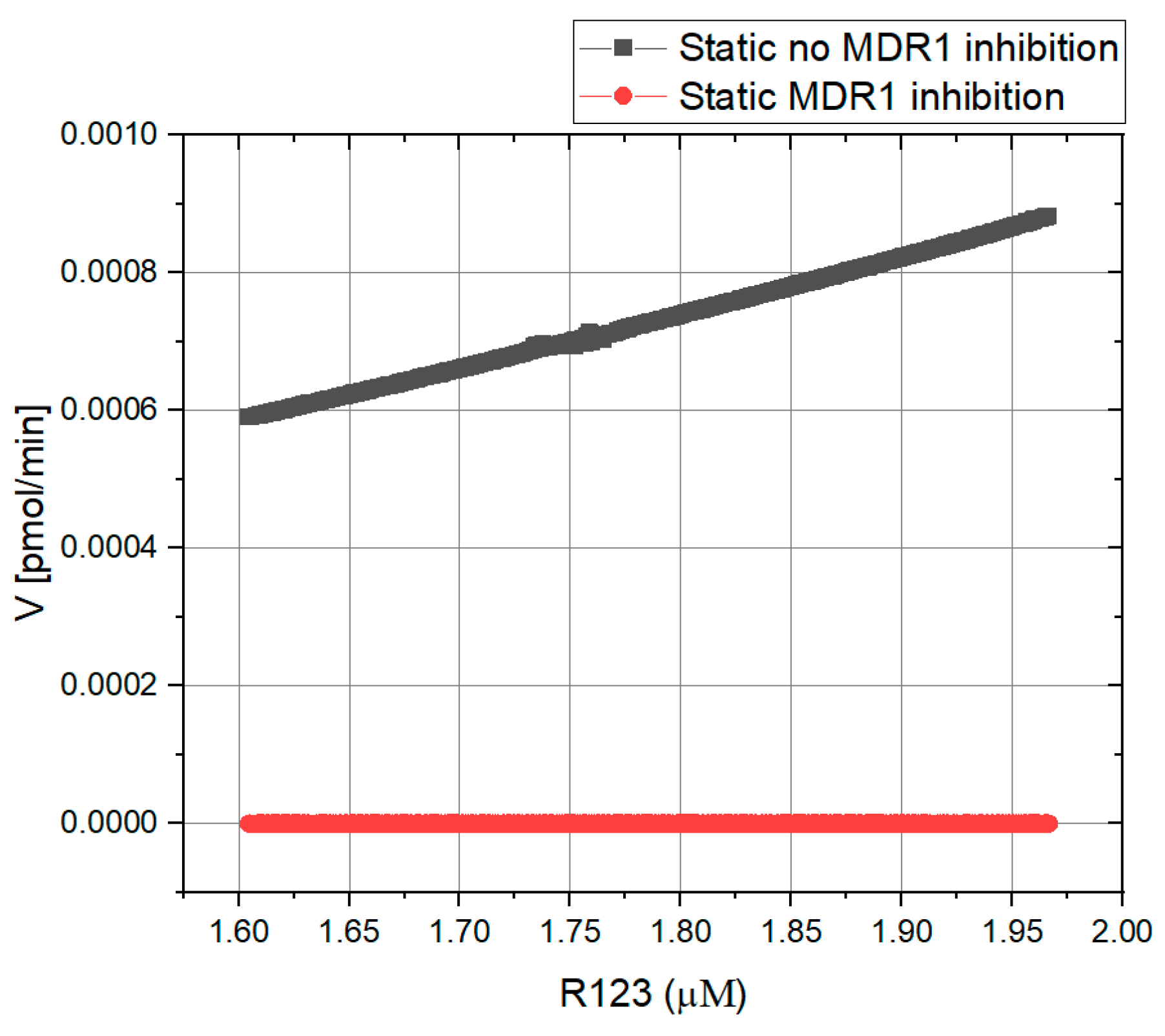
4.1.1. Kinetic Characterization of R123 Transport—Static Conditions
4.1.2. The Kinetic Characterization of R123 Transport—Dynamic Conditions
4.2. Apparent Permeability Coefficients
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jang, K.-J.; Mehr, A.P.; Hamilton, G.A.; McPartlin, L.A.; Chung, S.; Suh, K.-Y.; Ingber, D.E. Human kidney proximal tubule-on-a-chip for drug transport and nephrotoxicity assessment. Integr. Biol. 2013, 5, 1119–1129. [Google Scholar] [CrossRef] [PubMed]
- Segerer, K.; Wanner, C.; Steffel, J.; Luescher, T. Niere und Ableitende Harnwege; Springer: Berlin/Heidelberg, Germany, 2014; ISBN 978-3-642-28235-5. [Google Scholar]
- Kasper, D.L.; Fauci, A.S.; Longo, D.L. Harrisons Innere Medizin; 19. Auflage, deutsche Ausgabe; ABW Wissenschaftsverlag: Berlin, Germany; Thieme: Berlin, Germany; McGraw-Hill Education: New York, NY, USA, 2016; ISBN 3940615501. [Google Scholar]
- Morrissey, K.M.; Stocker, S.L.; Wittwer, M.B.; Xu, L.; Giacomini, K.M. Renal Transporters in Drug Development. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 503–529. [Google Scholar] [CrossRef] [PubMed]
- Krawczenko, A.; Bielawska-Pohl, A.; Wojtowicz, K.; Jura, R.; Paprocka, M.; Wojdat, E.; Kozłowska, U.; Klimczak, A.; Grillon, C.; Kieda, C.; et al. Expression and activity of multidrug resistance proteins in mature endothelial cells and their precursors: A challenging correlation. PLoS ONE 2017, 12, e0172371. [Google Scholar] [CrossRef]
- Nies, A.T.; Koepsell, H.; Damme, K.; Schwab, M. Organic Cation Transporters (OCTs, MATEs), In Vitro and In Vivo Evidence for the Importance in Drug Therapy. In Drug Transporters; Fromm, M.F., Kim, R.B., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 105–167. ISBN 978-3-642-14540-7. [Google Scholar]
- Fernandez, C.E.; Yen, R.W.; Perez, S.M.; Bedell, H.W.; Povsic, T.J.; Reichert, W.M.; Truskey, G.A. Human Vascular Microphysiological System for in vitro Drug Screening. Sci. Rep. 2016, 6, 21579. [Google Scholar] [CrossRef]
- Hering, Y.; Berthier, A.; Duez, H.; Lefebvre, P.; Deprez, B.; Gribbon, P.; Wolf, M.; Reinshagen, J.; Halley, F.; Hannemann, J.; et al. Development and implementation of a cell-based assay to discover agonists of the nuclear receptor REV-ERBα. J. Biol. Methods 2018, 5, 94. [Google Scholar] [CrossRef]
- Doke, S.K.; Dhawale, S.C. Alternatives to animal testing: A review. Saudi Pharm. J. 2015, 23, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Astashkina, A.; Mann, B.; Grainger, D.W. A critical evaluation of in vitro cell culture models for high-throughput drug screening and toxicity. Pharmacol. Ther. 2012, 134, 82–106. [Google Scholar] [CrossRef] [PubMed]
- Blom, K.; Nygren, P.; Alvarsson, J.; Larsson, R.; Andersson, C.R. Ex Vivo Assessment of Drug Activity in Patient Tumor Cells as a Basis for Tailored Cancer Therapy. J. Lab. Autom. 2016, 21, 178–187. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Takayama, S. Organ-on-a-chip and the kidney. Kidney Res. Clin. Pract. 2015, 34, 165–169. [Google Scholar] [CrossRef]
- Schophuizen, C.M.; de Napoli, I.E.; Jansen, J.; Teixeira, S.; Wilmer, M.J.; Hoenderop, J.G.; van den Heuvel, L.P.; Masereeuw, R.; Stamatialis, D. Development of a living membrane comprising a functional human renal proximal tubule cell monolayer on polyethersulfone polymeric membrane. Acta Biomater. 2015, 14, 22–32. [Google Scholar] [CrossRef]
- Sakolish, C.M.; Philip, B.; Mahler, G.J. A human proximal tubule-on-a-chip to study renal disease and toxicity. Biomicrofluidics 2019, 13, 14107. [Google Scholar] [CrossRef] [PubMed]
- Vormann, M.K.; Gijzen, L.; Hutter, S.; Boot, L.; Nicolas, A.; van den Heuvel, A.; Vriend, J.; Ng, C.P.; Nieskens, T.T.G.; van Duinen, V.; et al. Nephrotoxicity and Kidney Transport Assessment on 3D Perfused Proximal Tubules. AAPS J. 2018, 20, 90. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Gotoh, N.; Yan, Q.; Du, Z.; Weinstein, A.M.; Wang, T.; Weinbaum, S. Shear-induced reorganization of renal proximal tubule cell actin cytoskeleton and apical junctional complexes. Proc. Natl. Acad. Sci. USA 2008, 105, 11418–11423. [Google Scholar] [CrossRef]
- Forsythe, P.; Paterson, S. Ciclosporin 10 years on: Indications and efficacy. Vet. Rec. 2014, 174, 13–21. [Google Scholar] [CrossRef]
- Lin, K.; Gallay, P. Curing a viral infection by targeting the host: The example of cyclophilin inhibitors. Antivir. Res. 2013, 99, 68–77. [Google Scholar] [CrossRef]
- Hanino, M. Mechanism of Cyclosporin A-Induced Gingival Hyperplasia. Ph.D. Dissertation, Queen Mary University of London, London, UK, 2011. [Google Scholar]
- Fox, E.; Bates, S.E. Tariquidar (XR9576): A P-glycoprotein drug efflux pump inhibitor. Expert Rev. Anticancer Ther. 2007, 7, 447–459. [Google Scholar] [CrossRef] [PubMed]
- Wieser, M.; Stadler, G.; Jennings, P.; Streubel, B.; Pfaller, W.; Ambros, P.; Riedl, C.; Katinger, H.; Grillari, J.; Grillari-Voglauer, R. hTERT alone immortalizes epithelial cells of renal proximal tubules without changing their functional characteristics. Am. J. Physiol. Ren. Physiol. 2008, 295, F1365–F1375. [Google Scholar] [CrossRef]
- Forster, S.; Thumser, A.E.; Hood, S.R.; Plant, N. Characterization of Rhodamine-123 as a Tracer Dye for Use In In vitro Drug Transport Assays. PLoS ONE 2012, 7, e33253. [Google Scholar] [CrossRef]
- Schmieder, F.; Behrens, S.; Reustle, N.; Franke, N.; Sonntag, F.; Sradnick, J.; Hohenstein, B. A microphysiological system to investigate the pressure dependent filtration at an artificial glomerular kidney barrier. Curr. Dir. Biomed. Eng. 2019, 5, 389–391. [Google Scholar] [CrossRef]
- Weber, E.J.; Chapron, A.; Chapron, B.D.; Voellinger, J.L.; Lidberg, K.A.; Yeung, C.K.; Wang, Z.; Yamaura, Y.; Hailey, D.W.; Neumann, T.; et al. Development of a microphysiological model of human kidney proximal tubule function. Kidney Int. 2016, 90, 627–637. [Google Scholar] [CrossRef]
- Huppelsberg, J.; Walter, K. Kurzlehrbuch Physiologie, 4th ed.; Thieme: Stuttgart, Germany, 2013; ISBN 9783131364340. [Google Scholar]
- Abcam. Immunocytochemistry and Immunofluorescence Staining Protocol. Available online: https://www.abcam.com/protocols/immunocytochemistry-immunofluorescence-protocol (accessed on 5 March 2022).
- Mac Fhearraigh, S. ELISA Assays Principles, Protocols, Methods & Kits. Available online: https://www.assaygenie.com/elisa-assay-principles-methods-kits#Protocols (accessed on 12 January 2022).
- Wang, Y.; Hao, D.; Stein, W.D.; Yang, L. A kinetic study of Rhodamine123 pumping by P-glycoprotein. Biochim. Biophys. Acta 2006, 1758, 1671–1676. [Google Scholar] [CrossRef]
- Bentz, J.; Tran, T.T.; Polli, J.W.; Ayrton, A.; Ellens, H. The steady-state Michaelis-Menten analysis of P-glycoprotein mediated transport through a confluent cell monolayer cannot predict the correct Michaelis constant Km. Pharm. Res. 2005, 22, 1667–1677. [Google Scholar] [CrossRef]
- Aller, S.G.; Yu, J.; Ward, A.; Weng, Y.; Chittaboina, S.; Zhuo, R.; Harrell, P.M.; Trinh, Y.T.; Zhang, Q.; Urbatsch, I.L.; et al. Structure of P-Glycoprotein Reveals a Molecular Basis for Poly-Specific Drug Binding. Science 2009, 323, 1718–1722. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, A.B.; Ling, V. Positively cooperative sites for drug transport by P-glycoprotein with distinct drug specificities. Eur. J. Biochem. 1997, 250, 130–137. [Google Scholar] [CrossRef] [PubMed]
- González, M.L.; Vera, D.M.A.; Laiolo, J.; Joray, M.B.; Maccioni, M.; Palacios, S.M.; Molina, G.; Lanza, P.A.; Gancedo, S.; Rumjanek, V.; et al. Mechanism Underlying the Reversal of Drug Resistance in P-Glycoprotein-Expressing Leukemia Cells by Pinoresinol and the Study of a Derivative. Front. Pharmacol. 2017, 8, 205. [Google Scholar] [CrossRef] [PubMed]
- Zelman, A. Membrane Permeability. Biophys. J. 1972, 12, 414–419. [Google Scholar] [CrossRef]
- Jarzyńska, M.; Pietruszka, M. The application of the Kedem–Katchalsky equations to membrane transport of ethyl alcohol and glucose. Desalination 2011, 280, 14–19. [Google Scholar] [CrossRef]
- Petrak, K.; Goddard, P. Transport of macromolecules across the capillary walls. Adv. Drug Deliv. Rev. 1989, 3, 191–214. [Google Scholar] [CrossRef]
- Ginzburg, B.Z.; Katchalsky, A. The Frictional Coefficients of the Flows of Non-Electrolytes through Artificial Membranes. J. Gen. Physiol. 1963, 47, 403–418. [Google Scholar] [CrossRef]
- Michel, C.C.; Woodcock, T.E.; Curry, F.-R.E. Understanding and extending the Starling principle. Acta Anaesthesiol. Scand. 2020, 64, 1032–1037. [Google Scholar] [CrossRef] [PubMed]
- Palmer, B.F.; Alpern, R.J.; Seldin, D.W. Pathophysiology of Sodium Retention and Wastage. In Seldin and Giebisch’s The Kidney; Elsevier: Amsterdam, The Netherlands, 2013; pp. 1283–1317. ISBN 9780123814623. [Google Scholar]
- Jarzyńska, M. The application of practical Kedem-Katchalsky equations in membrane transport. Open Phys. 2006, 4, 429–438. [Google Scholar] [CrossRef]
- Chen, T.W.; Khanna, R.; Moore, H.; Twardowski, Z.J.; Nolph, K.D. Sieving and reflection coefficients for sodium salts and glucose during peritoneal dialysis in rats. J. Am. Soc. Nephrol. 1991, 2, 1092–1100. [Google Scholar] [CrossRef] [PubMed]
- Jarzyńska, M.; Pietruszka, M. Derivation of the formula for the filtration coefficient by application of Poiseuille’s law in membrane transport. Acta Soc. Bot. Pol. 2011, 78, 93–96. [Google Scholar] [CrossRef]
- Fortuna, A.; Alves, G.; Falcão, A.; Soares-da-Silva, P. Evaluation of the permeability and P-glycoprotein efflux of carbamazepine and several derivatives across mouse small intestine by the Ussing chamber technique. Epilepsia 2012, 53, 529–538. [Google Scholar] [CrossRef] [PubMed]
- Larson, B.; Banks, P.; Sherman, H.; Rothenberg, M. Automation of cell-based drug absorption assays in 96-well format using permeable support systems. J. Lab. Autom. 2012, 17, 222–232. [Google Scholar] [CrossRef]
- Ross, E.J.; Gordon, E.R.; Sothers, H.; Darji, R.; Baron, O.; Haithcock, D.; Prabhakarpandian, B.; Pant, K.; Myers, R.M.; Cooper, S.J.; et al. Three dimensional modeling of biologically relevant fluid shear stress in human renal tubule cells mimics in vivo transcriptional profiles. Sci. Rep. 2021, 11, 14053. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No MDR-1 Inhibition | MDR-1 Inhibition | |||
|---|---|---|---|---|
| Condition | Slope | R-Square | Slope | R-Square |
| Static | −0.008 ± 0.0001 | 0.99 | −0.009 ± 0.00002 | 0.98 |
| Pressure (0–2.5 h) | −0.24 ± 0.005 | 0.92 | −0.09 ± 0.0038 | 0.91 |
| Pressure (10–48 h) | −0.002 ± 0.0002 | 0.91 | −0.005 ± 0.00003 | 0.96 |
| Flow (0–2.5 h) | −0.17 ± 0.002 | 0.96 | −0.017 ± 0.0009 | 0.97 |
| Flow (20–48 h) | −0.006 ± 0.0001 | 0.97 | −0.008 ± 0.0001 | 0.99 |
| Papp (cm/s) × 10− 6 | ||
|---|---|---|
| Condition | No MDR1 Inhibition | MDR1 Inhibition |
| Static | 2.35 ± 0.003 | 2.65 ± 0.01 |
| Pressure (0−2.5 h) | 70.55 ± 1.47 | 26.45 ± 1.12 |
| Pressure (10−48 h) | 1.43 ± 0.10 | 1.83 ± 0.11 |
| Flow (0−2.5 h) | 49.97 ± 0.59 | 5.58 ± 0.06 |
| Flow (20−48 h) | 4.41 ± 0.06 | 3.22 ± 0.04 |
| Km (μM) | Vmax (pmol/min) | n | |
|---|---|---|---|
| From the literature | ≅17.5 [4,22,28,31] | 0.0525 [22] | 2 [16,22,28] |
| Obtained values (present study) | 17.47 ± 8 | 0.069 ± 0.05 | 2 |
| Km (μM) | V’max (pmol/min) | n | |
|---|---|---|---|
| From the literature | ≅17.5 [4,22,28,31] | 2.18 × 10−7 (from Equation (3)) [32] | 2 [22,28] |
| Obtained values (present study) | 11.06 ± 0.25 | 1.25 × 10−7 ± 3 × 10−9 (from curve fitting) | 2 |
| Conditions of Passive Transport of R123 | Symbols | Calculated from Equation | Measured |
|---|---|---|---|
| Pressure | 0.0102 | 0.027 | |
| Flow | 0.026 | 0.064 |
| Conditions of Total Transport Rate of R123 | Symbol | Measured |
|---|---|---|
| Static | 0.068 | |
| Pressure | 0.069 | |
| Flow | 0.112 |
| Conditions of Active Transport Rate of R123 | Symbol | Measured |
|---|---|---|
| Static | 0.038 | |
| Pressure | 0.042 | |
| Flow | 0.048 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Namazian Jam, N.; Gottlöber, F.; Hempel, M.; Dzekhtsiarova, Y.; Behrens, S.; Sonntag, F.; Sradnick, J.; Hugo, C.; Schmieder, F. Microphysiological Conditions Do Not Affect MDR1-Mediated Transport of Rhodamine 123 above an Artificial Proximal Tubule. Biomedicines 2023, 11, 2045. https://doi.org/10.3390/biomedicines11072045
Namazian Jam N, Gottlöber F, Hempel M, Dzekhtsiarova Y, Behrens S, Sonntag F, Sradnick J, Hugo C, Schmieder F. Microphysiological Conditions Do Not Affect MDR1-Mediated Transport of Rhodamine 123 above an Artificial Proximal Tubule. Biomedicines. 2023; 11(7):2045. https://doi.org/10.3390/biomedicines11072045
Chicago/Turabian StyleNamazian Jam, Negin, Felix Gottlöber, Melanie Hempel, Yuliya Dzekhtsiarova, Stephan Behrens, Frank Sonntag, Jan Sradnick, Christian Hugo, and Florian Schmieder. 2023. "Microphysiological Conditions Do Not Affect MDR1-Mediated Transport of Rhodamine 123 above an Artificial Proximal Tubule" Biomedicines 11, no. 7: 2045. https://doi.org/10.3390/biomedicines11072045
APA StyleNamazian Jam, N., Gottlöber, F., Hempel, M., Dzekhtsiarova, Y., Behrens, S., Sonntag, F., Sradnick, J., Hugo, C., & Schmieder, F. (2023). Microphysiological Conditions Do Not Affect MDR1-Mediated Transport of Rhodamine 123 above an Artificial Proximal Tubule. Biomedicines, 11(7), 2045. https://doi.org/10.3390/biomedicines11072045