
Determination of the Minimum Sample Amount for Capillary Electrophoresis-Fourier Transform Mass Spectrometry (CE-FTMS)-Based Metabolomics of Colorectal Cancer Biopsies

Abstract
1. Introduction
2. Materials and Methods
2.1. Specimen Collection and Pretreatment
2.2. Metabolite Extraction
2.3. Metabolome Analysis
2.4. Statistical Analysis
3. Results
3.1. Patient Characteristics
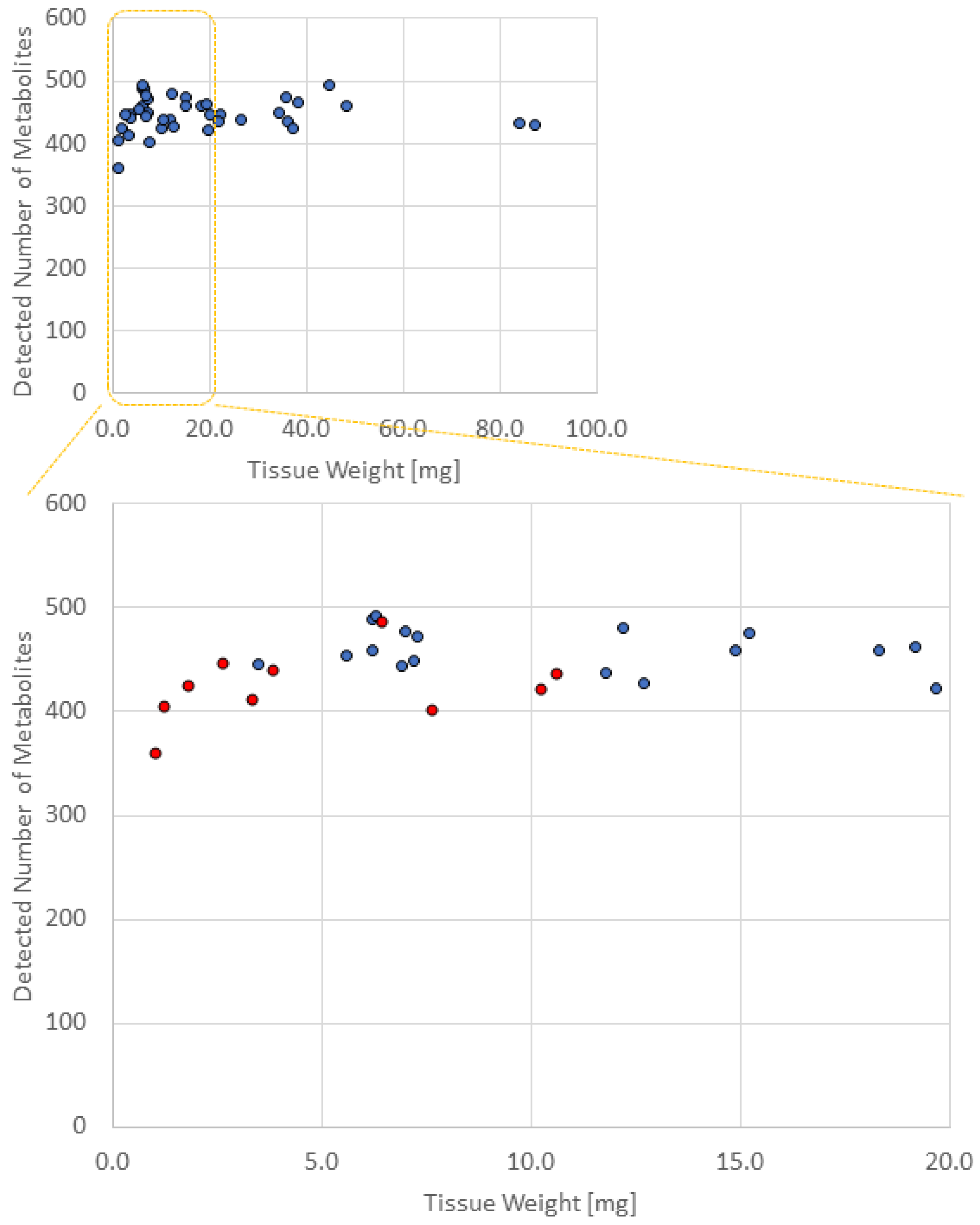
3.2. Comparison of the Number of Metabolites Detected in Different Numbers of Biopsied Samples
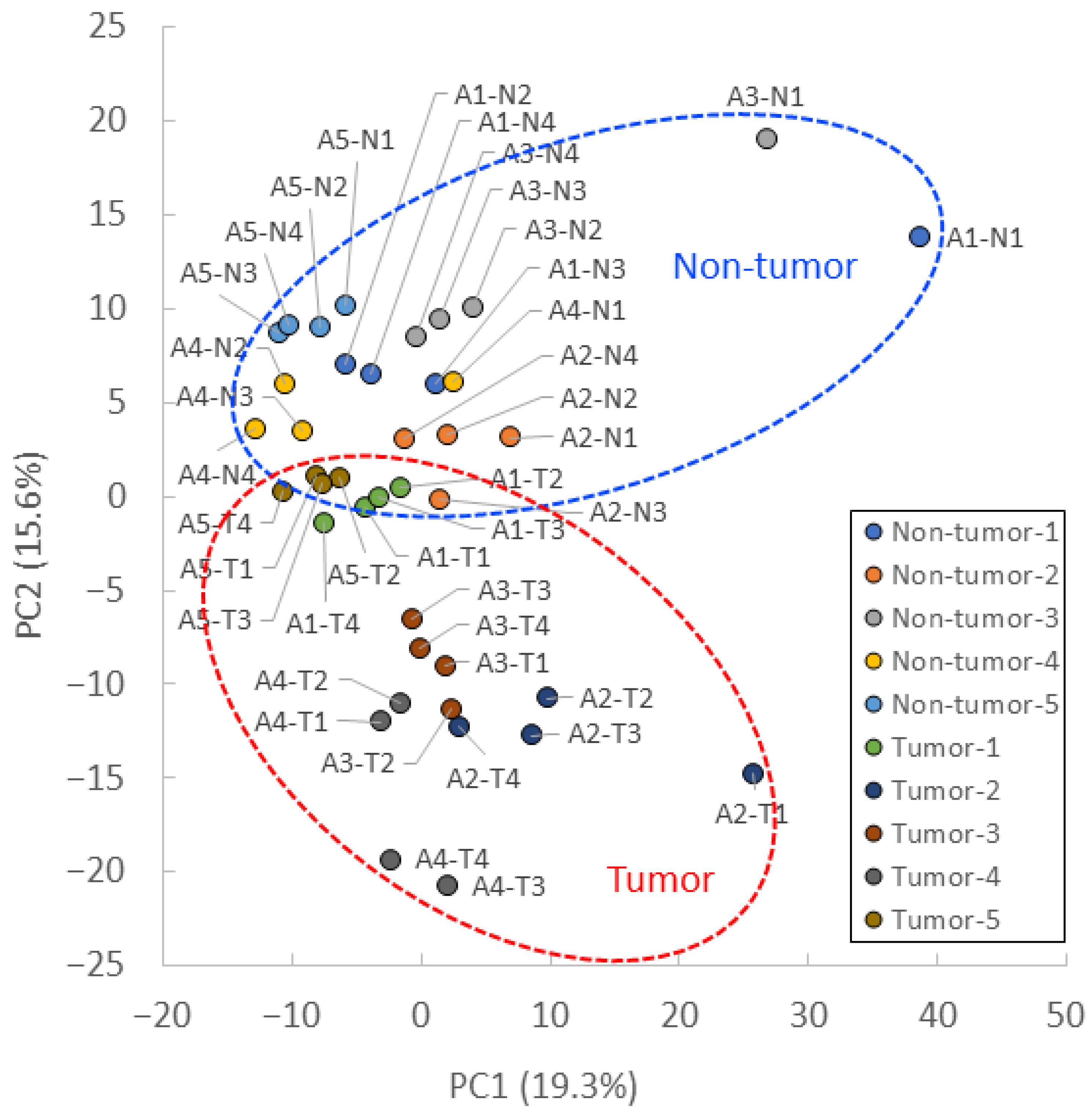
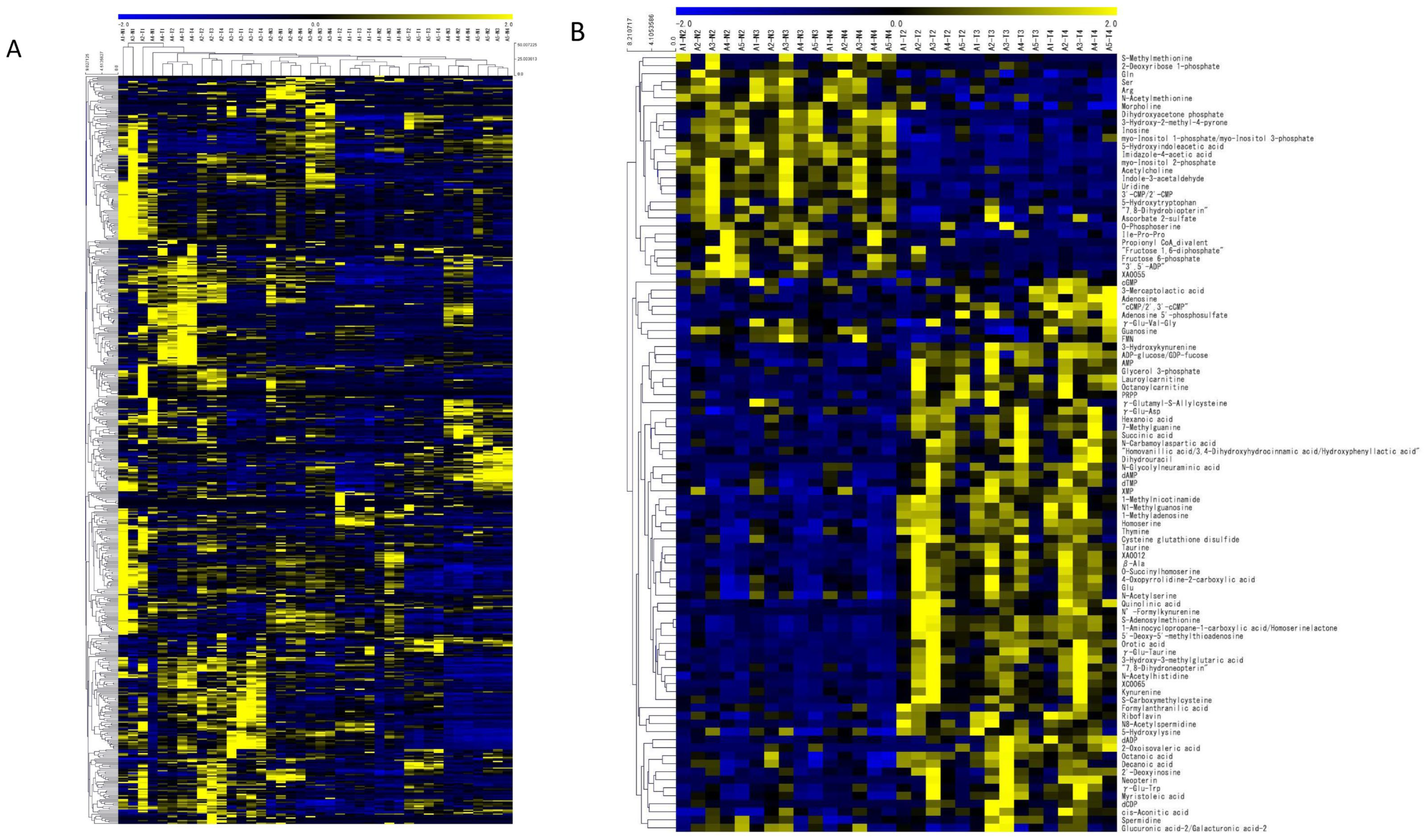
3.3. PCA and Heat Maps
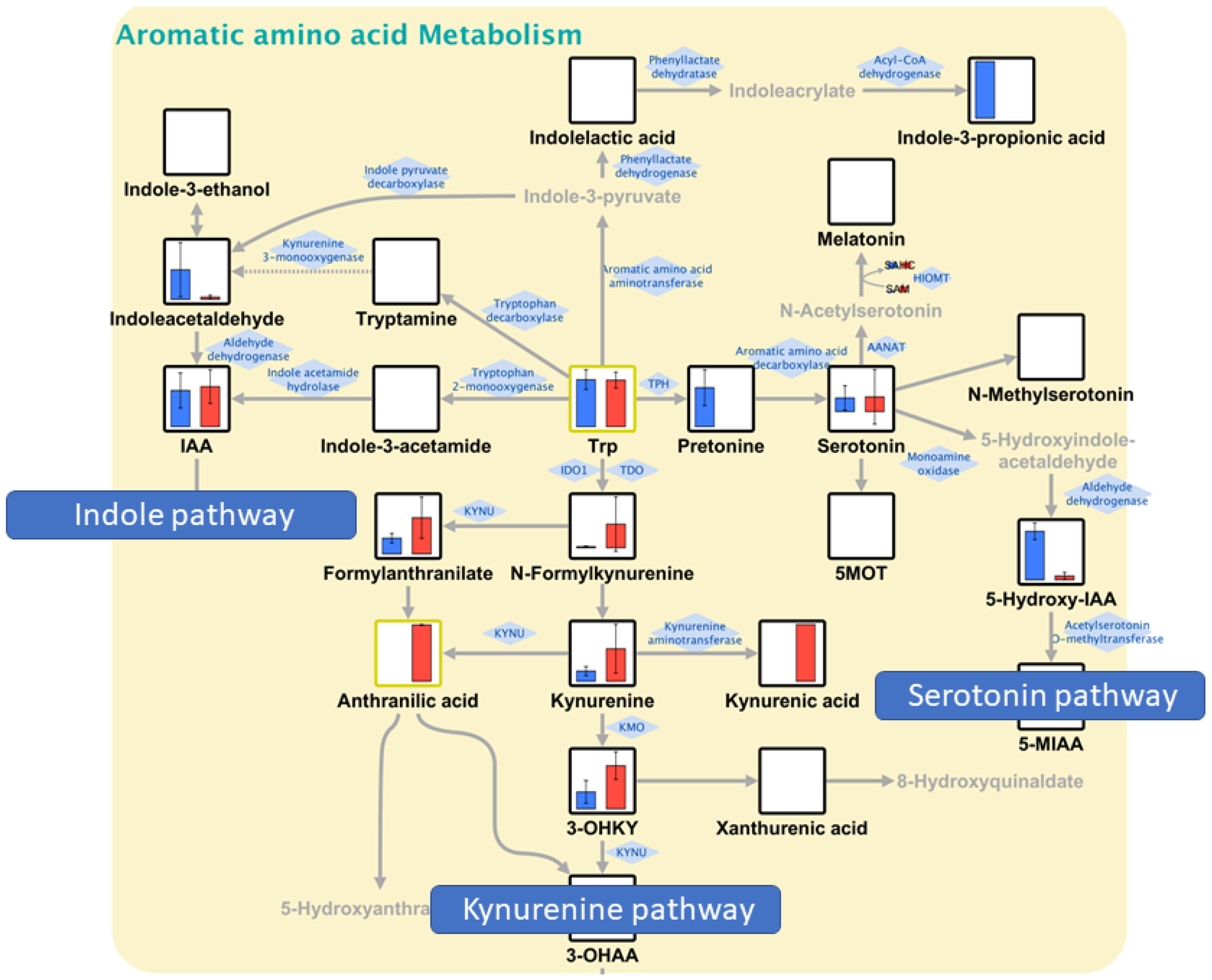
3.4. Pathway Maps and QMSEA
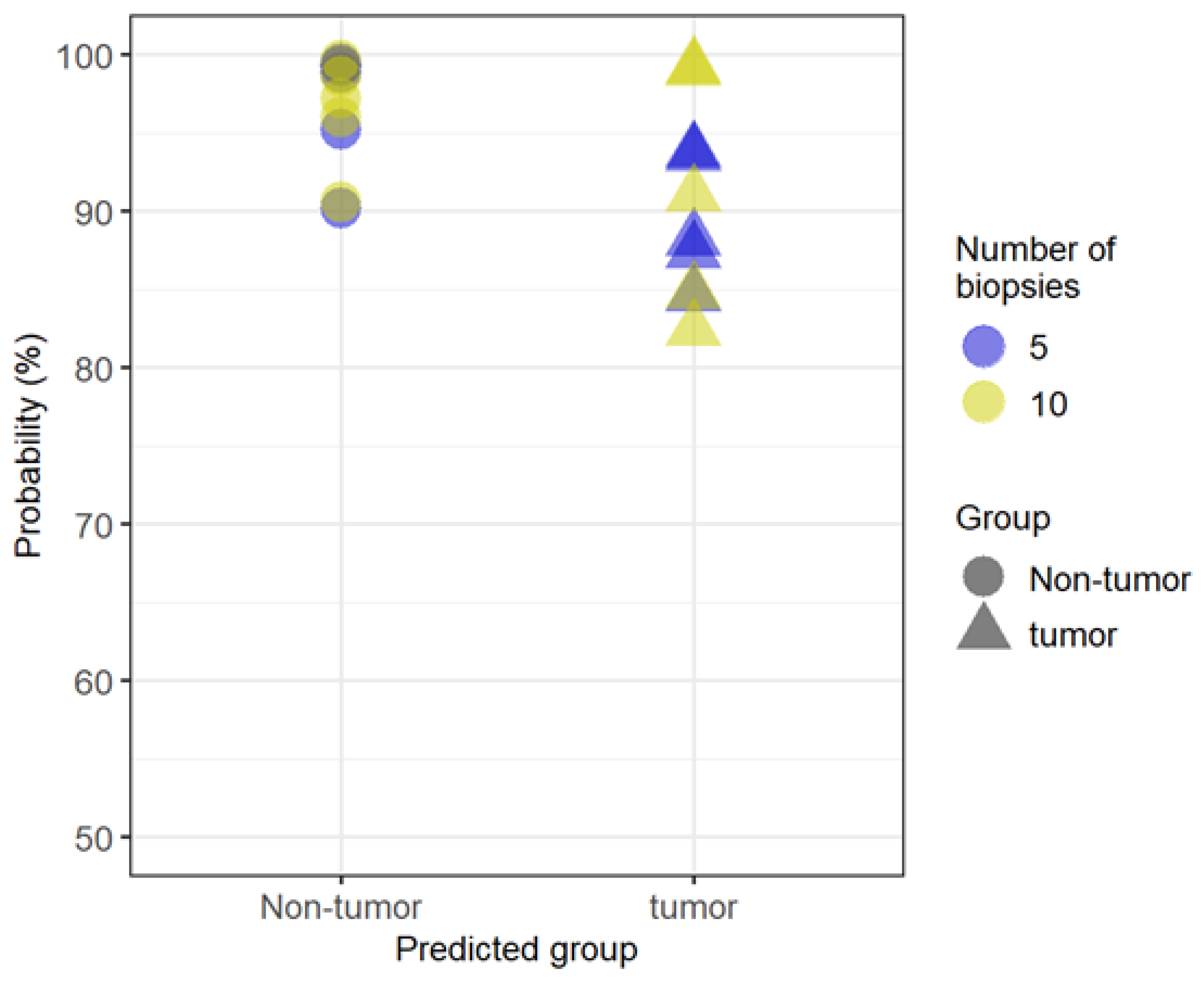
3.5. Random Forest
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, N.; Lu, B.; Luo, C.; Cai, J.; Lu, M.; Zhang, Y.; Chen, H.; Dai, M. Incidence, mortality, survival, risk factor and screening of colorectal cancer: A comparison among China, Europe, and northern America. Cancer Lett. 2021, 522, 255–268. [Google Scholar] [CrossRef] [PubMed]
- Heidelberger, C.; Chaudhuri, N.K.; Danneberg, P.; Mooren, D.; Griesbach, L.; Duschinsky, R.; Schnitzer, R.J.; Pleven, E.; Scheiner, J. Fluorinated pyrimidines, a new class of tumour-inhibitory compounds. Nature 1957, 179, 663–666. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, D.; Pyrhönen, S.; James, R.D.; Punt, C.J.; Hickish, T.F.; Heikkila, R.; Johannesen, T.B.; Starkhammar, H.; Topham, C.A.; Awad, L.; et al. Randomised trial of irinotecan plus supportive care versus supportive care alone after fluorouracil failure for patients with metastatic colorectal cancer. Lancet 1998, 352, 1413–1418. [Google Scholar] [CrossRef] [PubMed]
- Rougier, P.; Van Cutsem, E.; Bajetta, E.; Niederle, N.; Possinger, K.; Labianca, R.; Navarro, M.; Morant, R.; Bleiberg, H.; Wils, J.; et al. Randomised trial of irinotecan versus fluorouracil by continuous infusion after fluorouracil failure in patients with metastatic colorectal cancer. Lancet 1998, 352, 1407–1412. [Google Scholar] [CrossRef] [PubMed]
- Douillard, J.Y.; Cunningham, D.; Roth, A.D.; Navarro, M.; James, R.D.; Karasek, P.; Jandik, P.; Iveson, T.; Carmichael, J.; Alakl, M.; et al. Irinotecan combined with fluorouracil compared with fluorouracil alone as first-line treatment for metastatic colorectal cancer: A multicentre randomised trial. Lancet 2000, 355, 1041–1047. [Google Scholar] [CrossRef] [PubMed]
- Saltz, L.B.; Cox, J.V.; Blanke, C.; Rosen, L.S.; Fehrenbacher, L.; Moore, M.J.; Maroun, J.A.; Ackland, S.P.; Locker, P.K.; Pirotta, N.; et al. Irinotecan plus fluorouracil and leucovorin for metastatic colorectal cancer. N. Engl. J. Med. 2000, 343, 905–914. [Google Scholar] [CrossRef]
- Hurwitz, H.; Fehrenbacher, L.; Novotny, W.; Cartwright, T.; Hainsworth, J.; Heim, W.; Berlin, J.; Baron, A.; Griffing, S.; Holmgren, E.; et al. Bevacizumab plus irinotecan, fluorouracil, and leucovorin for metastatic colorectal cancer. N. Engl. J. Med. 2004, 350, 2335–2342. [Google Scholar] [CrossRef]
- Fuchs, C.S.; Marshall, J.; Mitchell, E.; Wierzbicki, R.; Ganju, V.; Jeffery, M.; Schulz, J.; Richards, D.; Soufi-Mahjoubi, R.; Wang, B.; et al. Randomized, controlled trial of irinotecan plus infusional, bolus, or oral fluoropyrimidines in first-line treatment of metastatic colorectal cancer: Results from the BICC-C Study. J. Clin. Oncol. 2007, 25, 4779–4786. [Google Scholar] [CrossRef]
- Saltz, L.B.; Clarke, S.; Diaz-Rubio, E.; Scheithauer, W.; Figer, A.; Wong, R.; Koski, S.; Lichinitser, M.; Yang, T.-S.; Rivera, F.; et al. Bevacizumab in combination with oxaliplatin-based chemotherapy as first-line therapy in metastatic colorectal cancer: A randomized phase III study. J. Clin. Oncol. 2008, 26, 2013–2019. [Google Scholar] [CrossRef]
- Van Cutsem, E.; Köhne, C.-H.; Hitre, E.; Zaluski, J.; Chien, C.-R.C.; Makhson, A.; D’Haens, G.; Pintér, T.; Lim, R.; Bodoky, G.; et al. Cetuximab and chemotherapy as initial treatment for metastatic colorectal cancer. N. Engl. J. Med. 2009, 360, 1408–1417. [Google Scholar] [CrossRef]
- Douillard, J.-Y.; Siena, S.; Cassidy, J.; Tabernero, J.; Burkes, R.; Barugel, M.; Humblet, Y.; Bodoky, G.; Cunningham, D.; Jassem, J.; et al. Randomized, phase III trial of panitumumab with infusional fluorouracil, leucovorin, and oxaliplatin (FOLFOX4) versus FOLFOX4 alone as first-line treatment in patients with previously untreated metastatic colorectal cancer: The PRIME study. J. Clin. Oncol. 2010, 28, 4697–4705. [Google Scholar] [CrossRef]
- Tabernero, J.; Yoshino, T.; Cohn, A.L.; Obermannova, R.; Bodoky, G.; Garcia-Carbonero, R.; Ciuleanu, T.-E.; Portnoy, D.C.; Van Cutsem, E.; Grothey, A.; et al. Ramucirumab versus placebo in combination with second-line FOLFIRI in patients with metastatic colorectal carcinoma that progressed during or after first-line therapy with bevacizumab, oxaliplatin, and a fluoropyrimidine (RAISE): A randomised, double-blind, multicentre, phase 3 study. Lancet Oncol. 2015, 16, 499–508. [Google Scholar] [CrossRef]
- Zimmermann, D.; Hartmann, M.; Moyer, M.P.; Nolte, J.; Baumbach, J.I. Determination of volatile products of human colon cell line metabolism by GC/MS analysis. Metabolomics 2007, 3, 13–17. [Google Scholar] [CrossRef]
- Yusof, H.M.; Ab-Rahim, S.; Ngah, W.Z.W.; Nathan, S.; Jamal, A.R.A.; Mazlan, M. Metabolomic characterization of colorectal cancer cell lines highlighting stage-specific alterations during cancer progression. BioImpacts 2021, 11, 147–156. [Google Scholar] [CrossRef]
- Sasaki, K.; Sagawa, H.; Suzuki, M.; Yamamoto, H.; Tomita, M.; Soga, T.; Ohashi, Y. Metabolomics platform with capillary electrophoresis coupled with high-resolution mass spectrometry for plasma analysis. Anal. Chem. 2019, 91, 1295–1301. [Google Scholar] [CrossRef]
- Sugimoto, M.; Wong, D.T.; Hirayama, A.; Soga, T.; Tomita, M. Capillary electrophoresis mass spectrometry-based saliva metabolomics identified oral, breast and pancreatic cancer-specific profiles. Metabolomics 2010, 6, 78–95. [Google Scholar] [CrossRef]
- Yamamoto, H.; Fujimori, T.; Sato, H.; Ishikawa, G.; Kami, K.; Ohashi, Y. Statistical hypothesis testing of factor loading in principal component analysis and its application to metabolite set enrichment analysis. BMC Bioinform. 2014, 15, 51. [Google Scholar] [CrossRef]
- Junker, B.H.; Klukas, C.; Schreiber, F. VANTED: A system for advanced data analysis and visualization in the context of biological networks. BMC Bioinform. 2006, 7, 109. [Google Scholar] [CrossRef]
- Saeed, A.I.; Bhagabati, N.K.; Braisted, J.C.; Liang, W.; Sharov, V.; Howe, E.A.; Li, J.; Thiagarajan, M.; White, J.A.; Quackenbush, J. TM4 microarray software suite. Methods Enzymol. 2006, 411, 134–193. [Google Scholar] [CrossRef]
- Pang, Z.; Chong, J.; Zhou, G.; de Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.-É.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the gap between raw spectra and functional insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef]
- Xia, J.; Wishart, D.S. Web-based inference of biological patterns, functions and pathways from metabolomic data using MetaboAnalyst. Nat. Protoc. 2011, 6, 743–760. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Santhanam, S.; Alvarado, D.M.; Ciorba, M.A. Therapeutic targeting of inflammation and tryptophan metabolism in colon and gastrointestinal cancer. Transl. Res. 2016, 167, 67–79. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, M.; Greathouse, K.L. Targeting dietary and microbial tryptophan-indole metabolism as therapeutic approaches to colon cancer. Nutrients 2021, 13, 1189. [Google Scholar] [CrossRef]
- Qiu, Y.; Cai, G.; Zhou, B.; Li, D.; Zhao, A.; Xie, G.; Li, H.; Cai, S.; Xie, D.; Huang, C.; et al. A distinct metabolic signature of human colorectal cancer with prognostic potential. Clin. Cancer Res. 2014, 20, 2136–2146. [Google Scholar] [CrossRef]
- Zhu, G.; Wang, Y.; Wang, W.; Shang, F.; Pei, B.; Zhao, Y.; Kong, D.; Fan, Z. Untargeted GC-MS-based metabolomics for early detection of colorectal cancer. Front. Oncol. 2021, 11, 729512. [Google Scholar] [CrossRef]
- Reitzer, L.; Wice, B.; Kennell, D. Evidence that glutamine, not sugar, is the major energy source for cultured HeLa cells. J. Biol. Chem. 1979, 254, 2669–2676. [Google Scholar] [CrossRef]
- Miyo, M.; Konno, M.; Nishida, N.; Sueda, T.; Noguchi, K.; Matsui, H.; Colvin, H.; Kawamoto, K.; Koseki, J.; Haraguchi, N.; et al. Metabolic adaptation to nutritional stress in human colorectal cancer. Sci. Rep. 2016, 6, 38415. [Google Scholar] [CrossRef]
- Huang, F.; Zhang, Q.; Ma, H.; Lv, Q.; Zhang, T. Expression of glutaminase is upregulated in colorectal cancer and of clinical significance. Int. J. Clin. Exp. Pathol. 2014, 7, 1093–1100. [Google Scholar] [PubMed]
- Camici, M.; Tozzi, M.G.; Allegrini, S.; Del Corso, A.; Sanfilippo, O.; Daidone, M.G.; De Marco, C.; Ipata, P.L. Purine salvage enzyme activities in normal and neoplastic human tissues. Cancer Biochem. Biophys. 1990, 11, 201–209. [Google Scholar] [PubMed]
- Melnyk, S.; Hakkak, R. Metabolic Status of lean and obese Zucker rats based on untargeted and targeted metabolomics analysis of serum. Biomedicines 2022, 10, 153. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient | A1 | A2 | A3 | A4 | A5 |
|---|---|---|---|---|---|
| Age (years) | 56 | 73 | 59 | 78 | 57 |
| Gender | M | M | M | M | M |
| BMI (kg/m2) | 25.5 | 25.5 | 30.7 | 21.3 | 23.3 |
| Blood sugar (mg/dL) | 108 | 104 | 131 | 114 | 99 |
| AST (IU/L) | 30 | 19 | 10 | 35 | 25 |
| ALT (IU/L) | 27 | 9 | 7 | 20 | 27 |
| γ-GTP (IU/L) | 178 | 21 | 19 | 134 | 59 |
| Cr (mg/dL) | 0.7 | 0.96 | 0.76 | 1.05 | 0.89 |
| eGFR (mL/min/1.73 m2) | 90.7 | 59.4 | 81.7 | 52.7 | 69.4 |
| Tumor location | rectum | rectum | sigmoid colon | rectum | rectum |
| Tumor size (mm) | 41 × 42 | 20 × 20 | 47 × 35 | 29 × 28 | 40 × 35 |
| Histological type | tub2 | tub1 > tub2 | tub1 > pap | tub1 > tub2 | tub1 > tub2 |
| Depth of tumor invasion | T2 | T1 | T4 | T4 | T3 |
| Lymphatic invasion | ly1 | ly0 | ly0 | ly0 | ly0 |
| Venous invasion | v1 | v1 | v1a | v1a | v1b |
| Lymph node metastasis | N0 | N0 | N0 | N0 | N0 |
| Sample Name | Number of Biopsies | Amount (mg) | Group Name | Detected Number of Metabolites |
|---|---|---|---|---|
| A1-N1 | 1 | 1.2 | Nontumor-1 | 405 |
| A1-N2 | 2 | 7.2 | 449 | |
| A1-N3 | 5 | 12.7 | 428 | |
| A1-N4 | 10 | 26.6 | 437 | |
| A2-N1 | 1 | 3.8 | Nontumor-2 | 441 |
| A2-N2 | 2 | 7.3 | 472 | |
| A2-N3 | 5 | 12.2 | 480 | |
| A2-N4 | 10 | 34.2 | 449 | |
| A3-N1 | 1 | 1.0 | Nontumor-3 | 360 |
| A3-N2 | 2 | 5.6 | 454 | |
| A3-N3 | 5 | 14.9 | 459 | |
| A3-N4 | 10 | 36.3 | 436 | |
| A4-N1 | 1 | 2.6 | Nontumor-4 | 447 |
| A4-N2 | 2 | 7.0 | 477 | |
| A4-N3 | 5 | 19.2 | 463 | |
| A4-N4 | 10 | 38.3 | 467 | |
| A5-N1 | 1 | 3.3 | Nontumor-5 | 412 |
| A5-N2 | 2 | 6.9 | 444 | |
| A5-N3 | 5 | 21.7 | 434 | |
| A5-N4 | 10 | 37.4 | 424 | |
| A1-T1 | 1 | 10.2 | Tumor-1 | 423 |
| A1-T2 | 2 | 6.2 | 459 | |
| A1-T3 | 5 | 19.7 | 422 | |
| A1-T4 | 10 | 87.2 | 431 | |
| A2-T1 | 1 | 1.8 | Tumor-2 | 425 |
| A2-T2 | 2 | 3.5 | 446 | |
| A2-T3 | 5 | 6.2 | 489 | |
| A2-T4 | 10 | 35.7 | 473 | |
| A3-T1 | 1 | 7.6 | Tumor-3 | 403 |
| A3-T2 | 2 | 11.8 | 437 | |
| A3-T3 | 5 | 22.4 | 445 | |
| A3-T4 | 10 | 48.4 | 459 | |
| A4-T1 | 1 | 6.4 | Tumor-4 | 488 |
| A4-T2 | 2 | 6.3 | 493 | |
| A4-T3 | 5 | 15.2 | 475 | |
| A4-T4 | 10 | 44.9 | 494 | |
| A5-T1 | 1 | 10.6 | Tumor-5 | 438 |
| A5-T2 | 2 | 20.1 | 446 | |
| A5-T3 | 5 | 18.3 | 459 | |
| A5-T4 | 10 | 84.0 | 433 |
| (1) N2 vs. T2 (Two Pieces of Biopsied Samples) | Raw p | (2) N3 vs. T3 (Five Pieces of Biopsied Samples) | Raw p | (3) N4 vs. T4 (Ten Pieces of Biopsied Samples) | Raw p | Statistical Significance |
|---|---|---|---|---|---|---|
| Taurine and hypotaurine metabolism | 1.47 × 10−3 | Nitrogen metabolism | 1.24 × 10−3 | Cysteine and methionine metabolism | 9.28 × 10−5 | (1)–(3) |
| Tryptophan metabolism | 2.57 × 10−3 | Tryptophan metabolism | 2.43 × 10−3 | Purine metabolism | 9.48 × 10−4 | (1)–(3) |
| Cysteine and methionine metabolism | 4.04 × 10−3 | Pyrimidine metabolism | 3.83 × 10−3 | Taurine and hypotaurine metabolism | 1.19 × 10−3 | (1)–(3) |
| Histidine metabolism | 5.44 × 10−3 | Glyoxylate and dicarboxylate metabolism | 7.01 × 10−3 | Glycerophospholipid metabolism | 1.82 × 10−3 | (1)–(3) |
| Folate biosynthesis | 7.52 × 10−3 | Purine metabolism | 7.16 × 10−3 | Pyrimidine metabolism | 2.97 × 10−3 | (1)–(3) |
| Nitrogen metabolism | 8.20 × 10−3 | D-glutamine and D-glutamate metabolism | 8.87 × 10−3 | Nitrogen metabolism | 3.71 × 10−3 | (1)–(3) |
| Pyrimidine metabolism | 1.29 × 10−2 | Fructose and mannose metabolism | 1.06 × 10−2 | Tryptophan metabolism | 3.79 × 10−3 | (1)–(3) |
| Riboflavin metabolism | 1.30 × 10−2 | Propanoate metabolism | 1.49 × 10−2 | D-glutamine and D-glutamate metabolism | 9.56 × 10−3 | (1)–(3) |
| D-glutamine and D-glutamate metabolism | 1.46 × 10−2 | Cysteine and methionine metabolism | 2.04 × 10−2 | Sulfur metabolism | 1.37 × 10−2 | (3) only |
| Purine metabolism | 1.95 × 10−2 | Inositol phosphate metabolism | 2.17 × 10−2 | Glycerolipid metabolism | 1.42 × 10−2 | (3) only |
| Propanoate metabolism | 1.98 × 10−2 | Beta-alanine metabolism | 2.27 × 10−2 | Glyoxylate and dicarboxylate metabolism | 1.48 × 10−2 | (2), (3) only |
| Glycerophospholipid metabolism | 2.15 × 10−2 | Amino sugar and nucleotide sugar metabolism | 2.35 × 10−2 | Histidine metabolism | 2.22 × 10−2 | (1), (3) only |
| Fructose and mannose metabolism | 2.21 × 10−2 | Taurine and hypotaurine metabolism | 2.46 × 10−2 | Fructose and mannose metabolism | 2.42 × 10−2 | (1)–(3) |
| Butanoate metabolism | 2.25 × 10−2 | Starch and sucrose metabolism | 3.36 × 10−2 | Folate biosynthesis | 2.72 × 10−2 | (1), (3) only |
| Pantothenate and CoA biosynthesis | 2.91 × 10−2 | Alanine, aspartate, and glutamate metabolism | 3.50 × 10−2 | Primary bile acid biosynthesis | 2.93 × 10−2 | (3) only |
| Arginine and proline metabolism | 2.94 × 10−2 | Neomycin, kanamycin, and gentamicin biosynthesis | 3.70 × 10−2 | Propanoate metabolism | 3.38 × 10−2 | (1)–(3) |
| Porphyrin and chlorophyll metabolism | 3.36 × 10−2 | Fatty acid biosynthesis | 3.70 × 10−2 | Sphingolipid metabolism | 3.45 × 10−2 | (3) only |
| Citrate cycle (TCA cycle) | 4.07 × 10−2 | Glycerophospholipid metabolism | 4.12 × 10−2 | Alanine, aspartate, and glutamate metabolism | 3.69 × 10−2 | (2), (3) only |
| One carbon pool by folate | 4.18 × 10−2 | Galactose metabolism | 4.91 × 10−2 | Nicotinate and nicotinamide metabolism | 4.50 × 10−2 | (3) only |
| Arginine and proline metabolism | 4.79 × 10−2 | (1), (3) only | ||||
| Amino sugar and nucleotide sugar metabolism | 4.80 × 10−2 | (2), (3) only |
| Metabolite | Importance |
|---|---|
| 5-Hydroxyindoleacetic acid | 100 |
| Indoleacetaldehyde | 69.60 |
| Formylanthranilate | 67.28 |
| XA0012 | 66.93 |
| 1-Methylnicotinamide | 62.38 |
| Taurine | 62.02 |
| Octanoylcarnitine | 58.29 |
| γ-Glu-Taurine | 55.30 |
| β-Ala | 49.10 |
| 5′-Deoxy-5′-methylthioadenosine | 42.98 |
| 1-Aminocyclopropane-1-carboxylic acid homoserine lactone | 42.30 |
| O-Succinylhomoserine | 42.13 |
| Myo-inositol 2-phosphate | 36.75 |
| Imidazole-4-acetic acid | 34.39 |
| Uridine | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sugishita, T.; Tokunaga, M.; Kami, K.; Terai, K.; Yamamoto, H.; Shinohara, H.; Kinugasa, Y. Determination of the Minimum Sample Amount for Capillary Electrophoresis-Fourier Transform Mass Spectrometry (CE-FTMS)-Based Metabolomics of Colorectal Cancer Biopsies. Biomedicines 2023, 11, 1706. https://doi.org/10.3390/biomedicines11061706
Sugishita T, Tokunaga M, Kami K, Terai K, Yamamoto H, Shinohara H, Kinugasa Y. Determination of the Minimum Sample Amount for Capillary Electrophoresis-Fourier Transform Mass Spectrometry (CE-FTMS)-Based Metabolomics of Colorectal Cancer Biopsies. Biomedicines. 2023; 11(6):1706. https://doi.org/10.3390/biomedicines11061706
Chicago/Turabian StyleSugishita, Tetsuo, Masanori Tokunaga, Kenjiro Kami, Kozue Terai, Hiroyuki Yamamoto, Hajime Shinohara, and Yusuke Kinugasa. 2023. "Determination of the Minimum Sample Amount for Capillary Electrophoresis-Fourier Transform Mass Spectrometry (CE-FTMS)-Based Metabolomics of Colorectal Cancer Biopsies" Biomedicines 11, no. 6: 1706. https://doi.org/10.3390/biomedicines11061706
APA StyleSugishita, T., Tokunaga, M., Kami, K., Terai, K., Yamamoto, H., Shinohara, H., & Kinugasa, Y. (2023). Determination of the Minimum Sample Amount for Capillary Electrophoresis-Fourier Transform Mass Spectrometry (CE-FTMS)-Based Metabolomics of Colorectal Cancer Biopsies. Biomedicines, 11(6), 1706. https://doi.org/10.3390/biomedicines11061706