
Anisotropy and Frequency Dependence of Signal Propagation in the Cerebellar Circuit Revealed by High-Density Multielectrode Array Recordings

Abstract
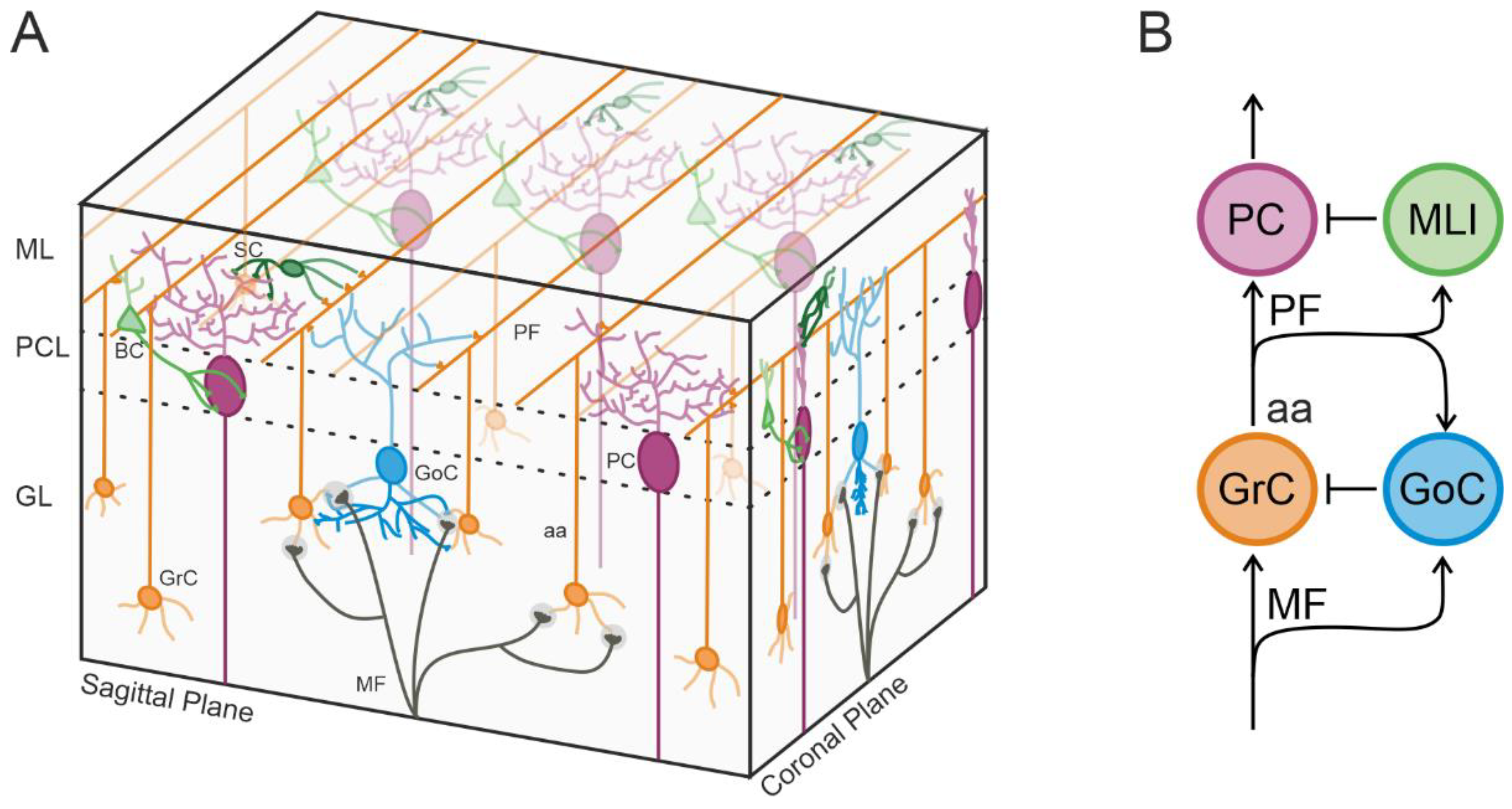
1. Introduction
2. Materials and Methods
2.1. Slice Preparation and Maintenance
2.2. High-Resolution Electrophysiological Recordings
2.3. Data Analysis
2.3.1. Local Field Potentials
2.3.2. Purkinje Cell Firing
3. Results

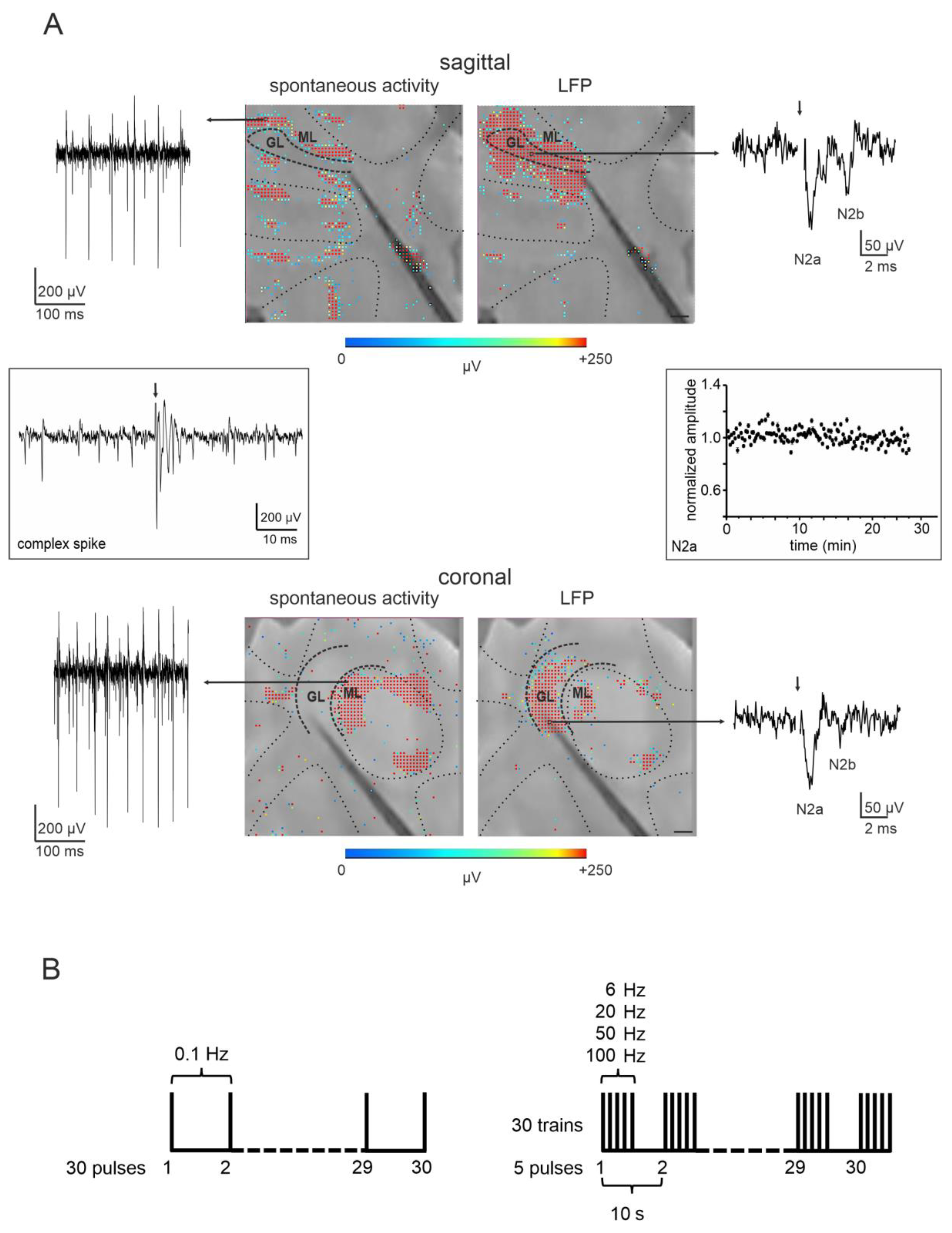
3.1. Characterization of Cerebellar Cortical Activity with HD-MEA


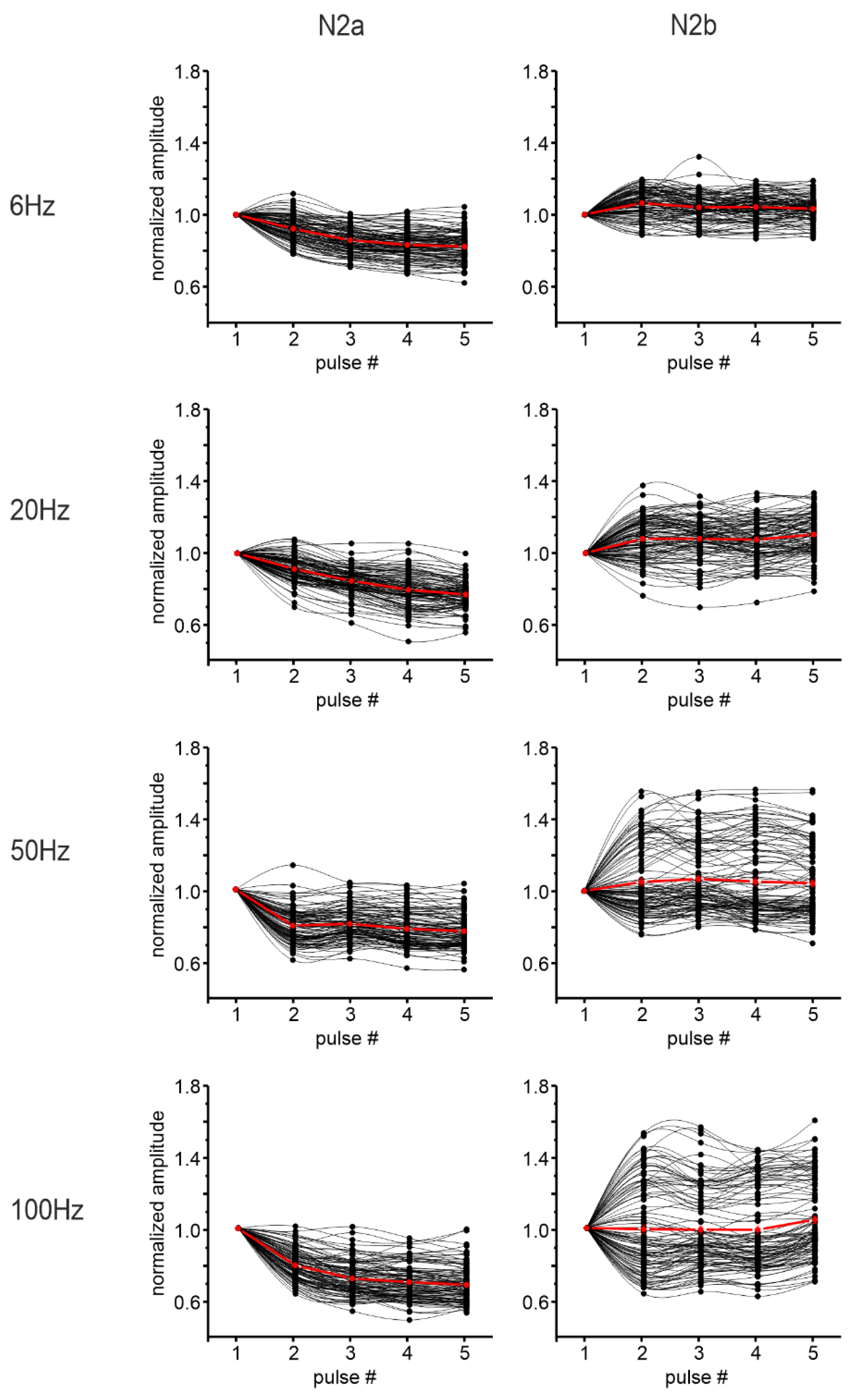
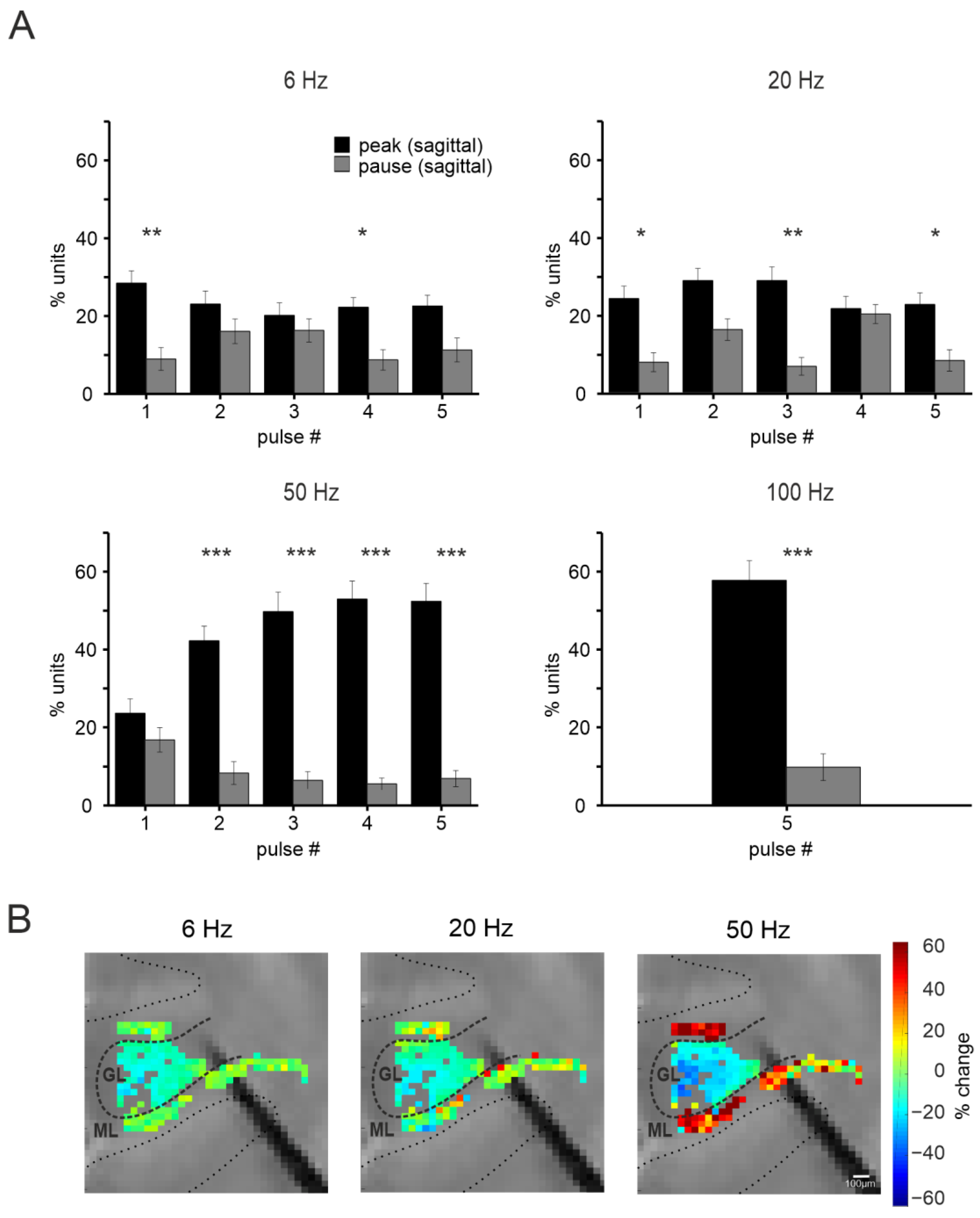
3.2. Short-Term Plasticity on the Sagittal Plane
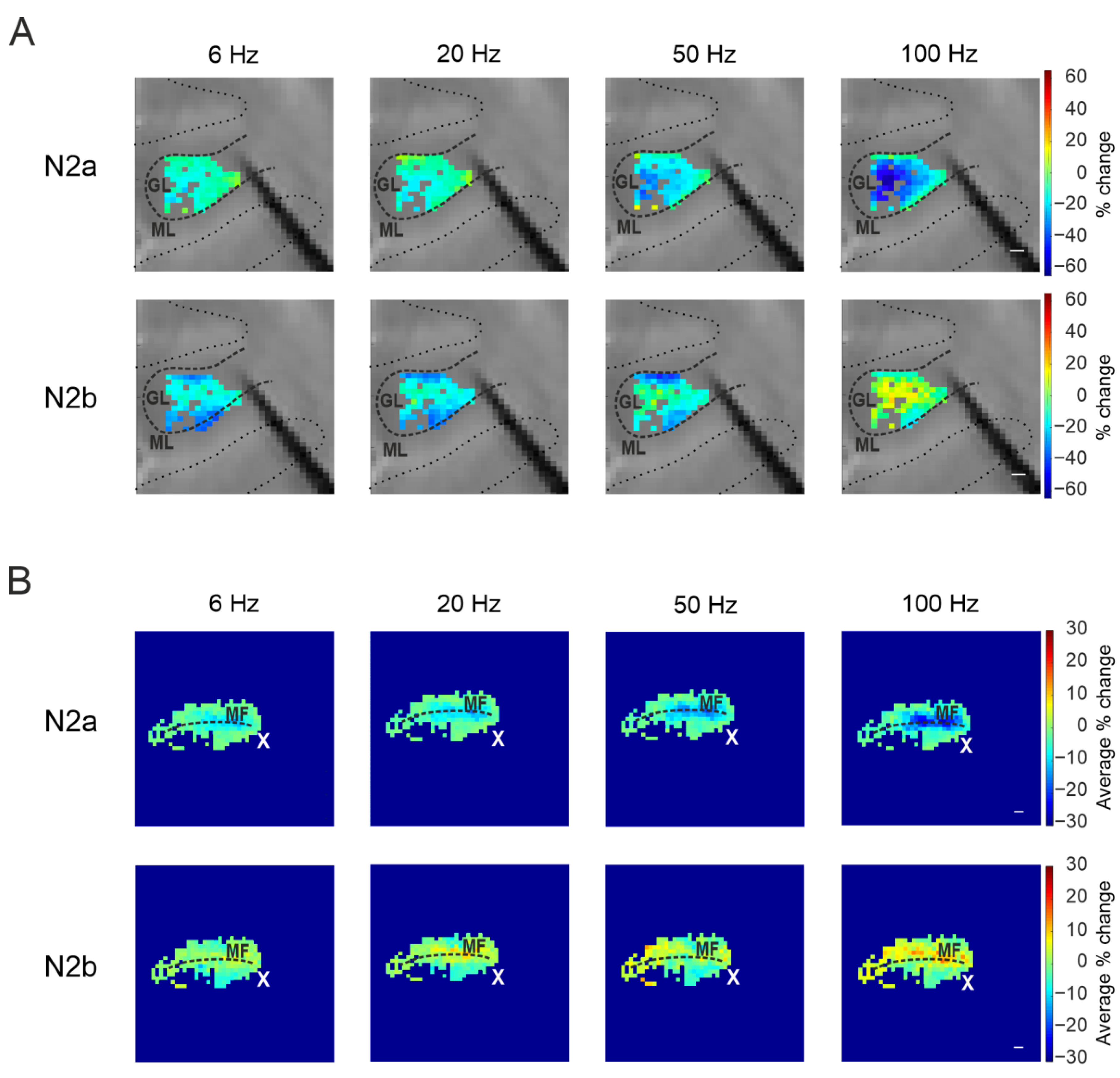
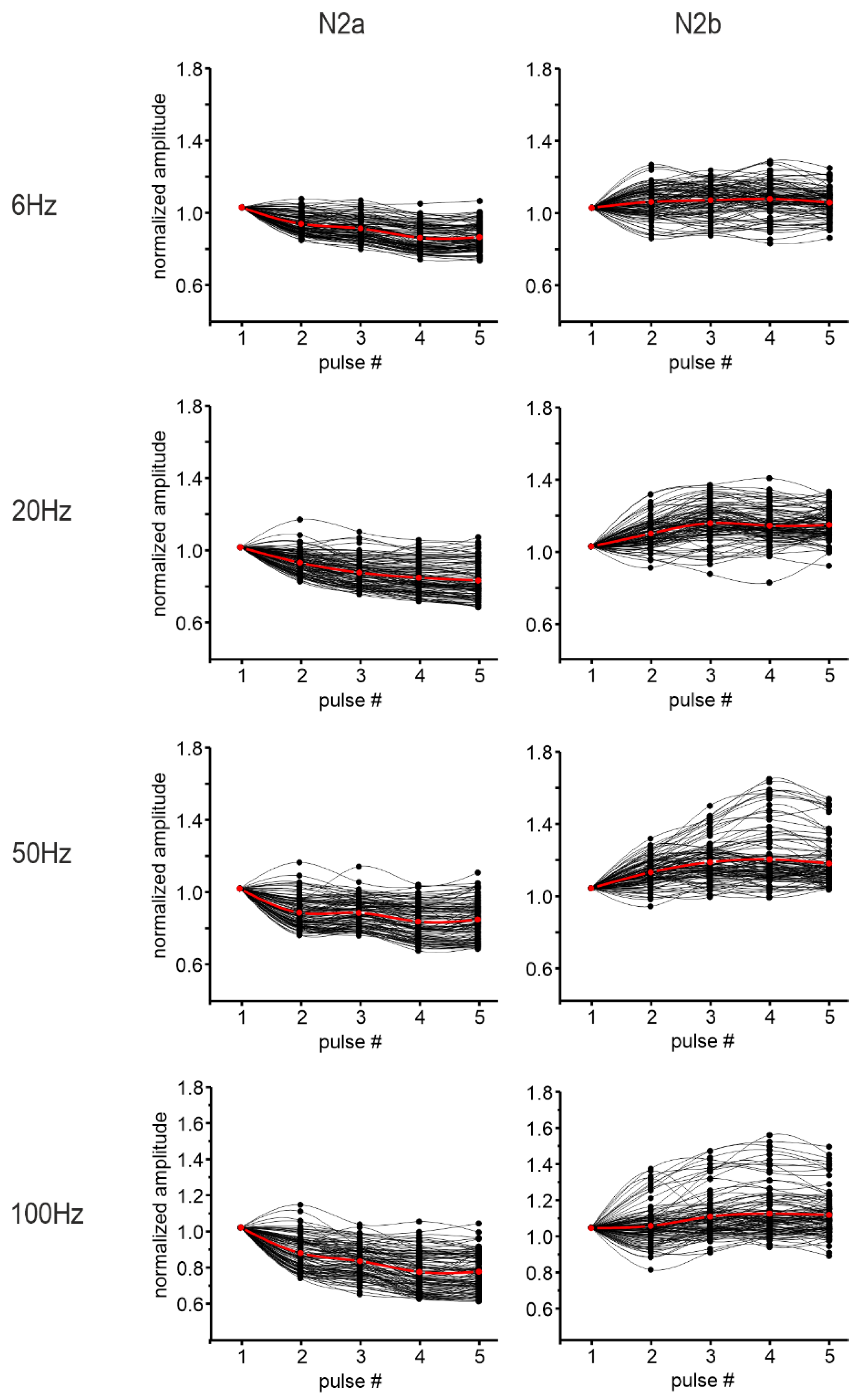
3.2.1. Granular Layer Responses at Different Input Frequencies
3.2.2. The Spatial Organization of Short-Term Plasticity in the Granular Layer
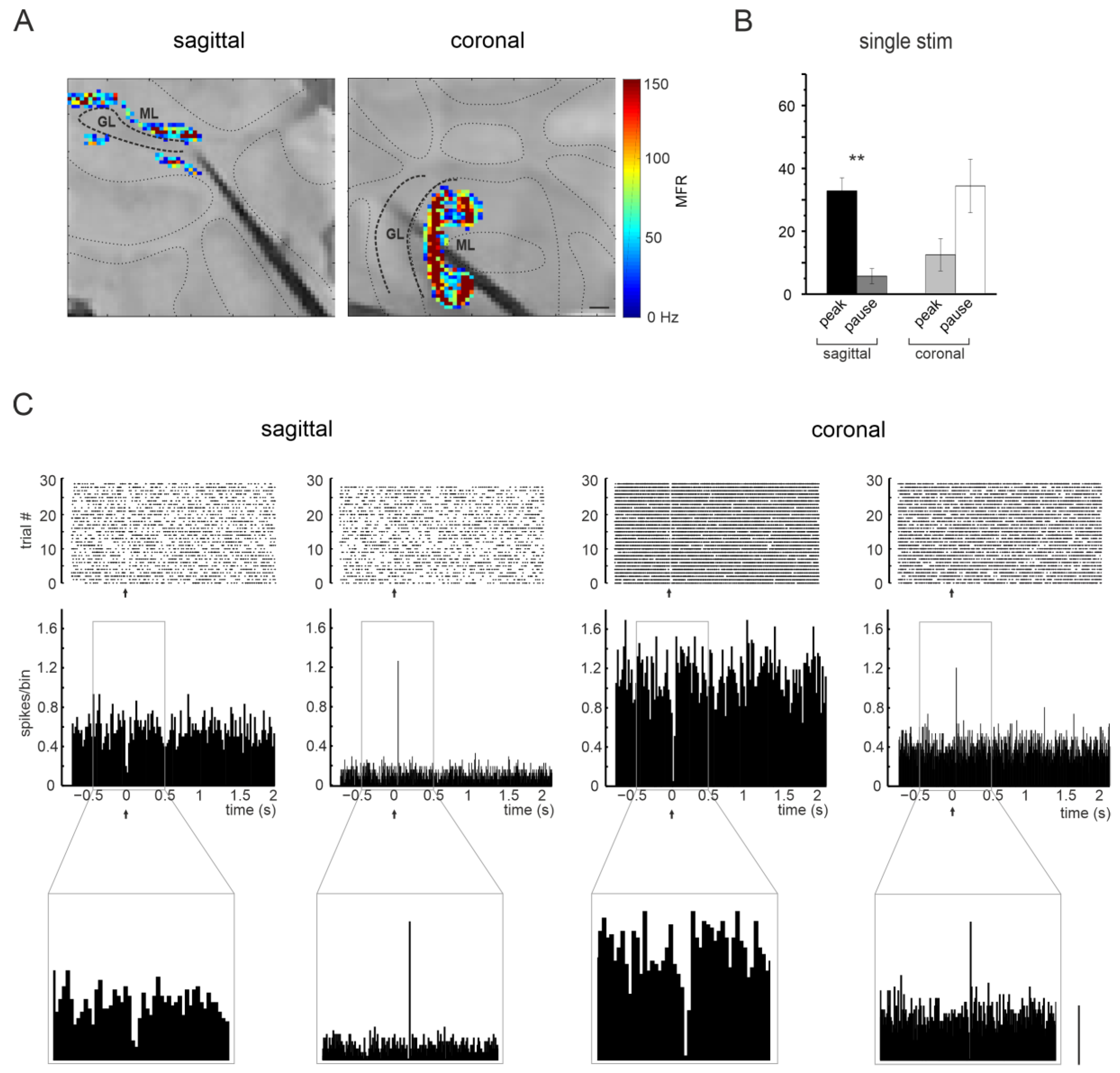
3.2.3. Purkinje Cells Responses at Different Input Frequencies

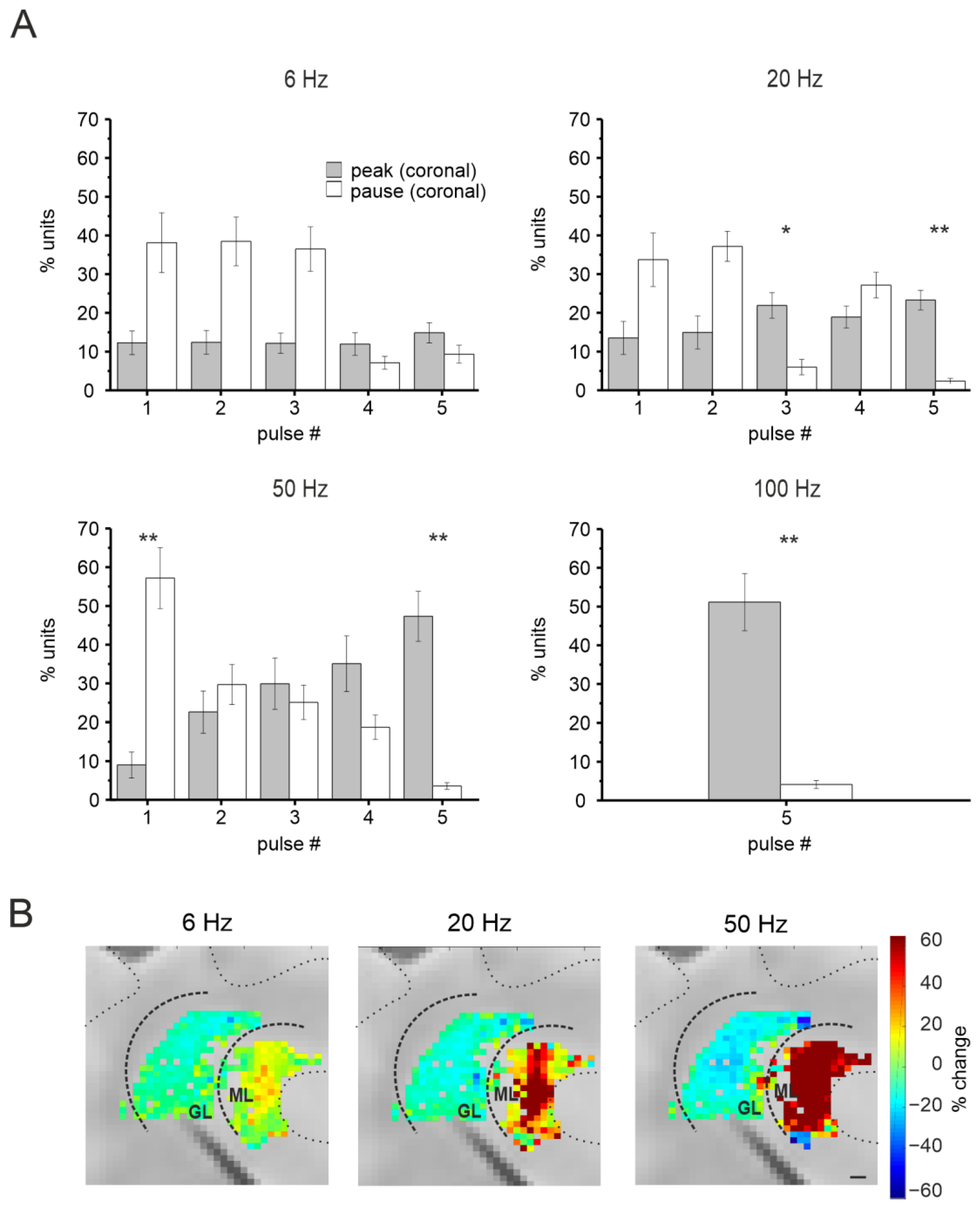
3.3. Short-Term Plasticity on the Coronal Plane
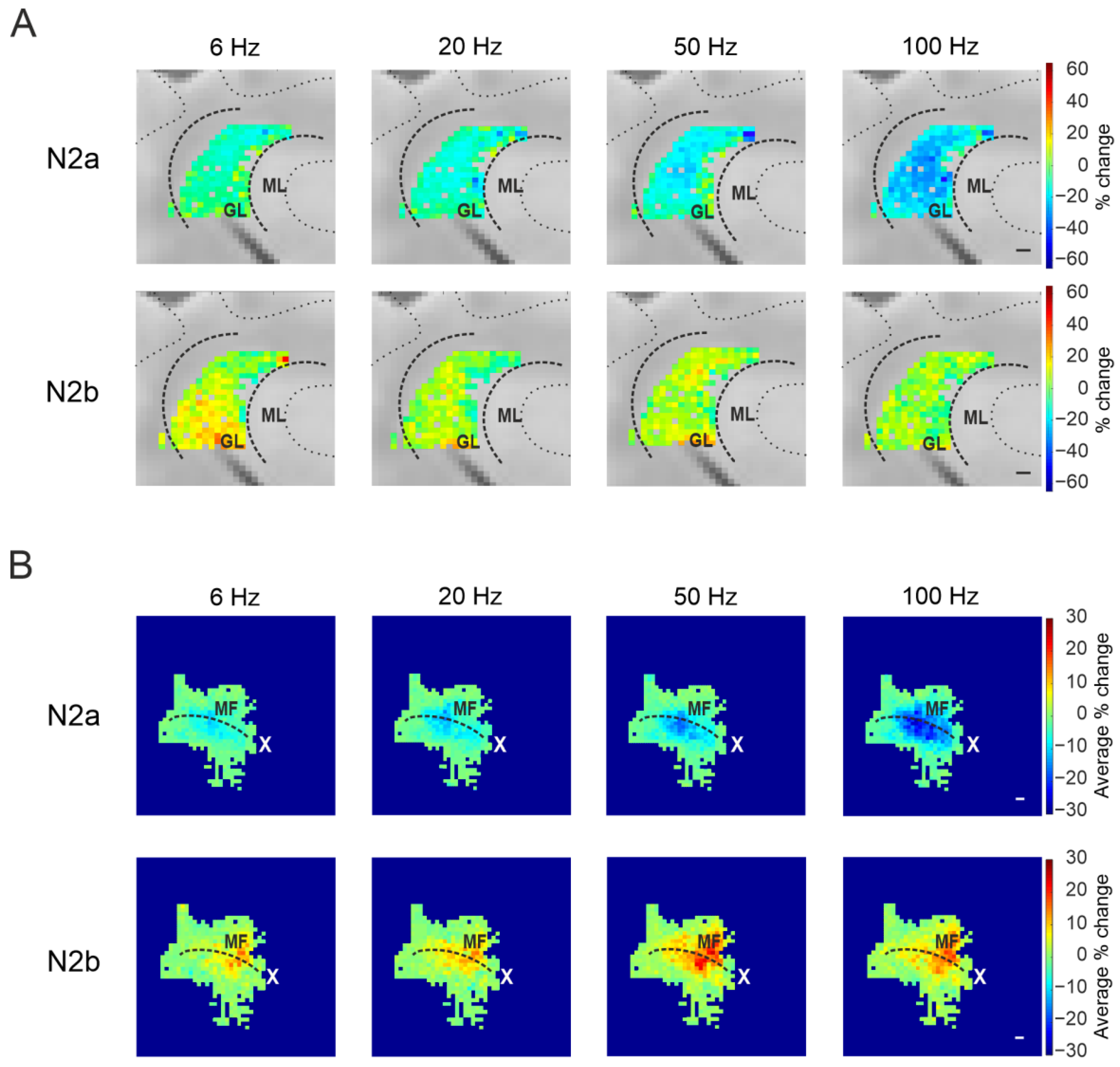
3.3.1. Granular Layer Responses at Different Input Frequencies
3.3.2. The Spatial Organization of Short-Term Plasticity in the Granular Layer

3.3.3. Purkinje Cells Responses at Different Input Frequencies

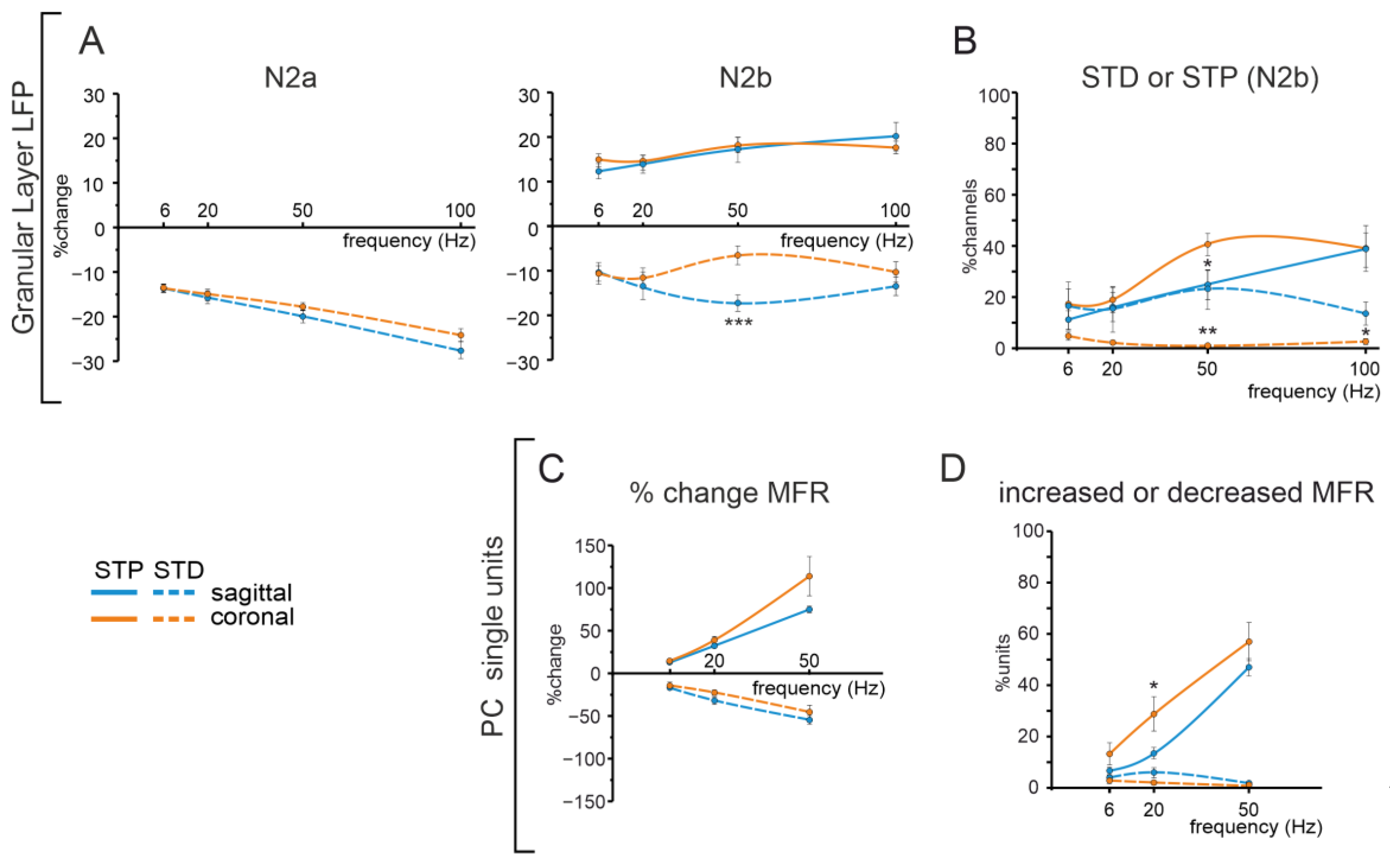
3.4. Comparison of Cerebellar Network Responses in the Sagittal and Coronal Planes
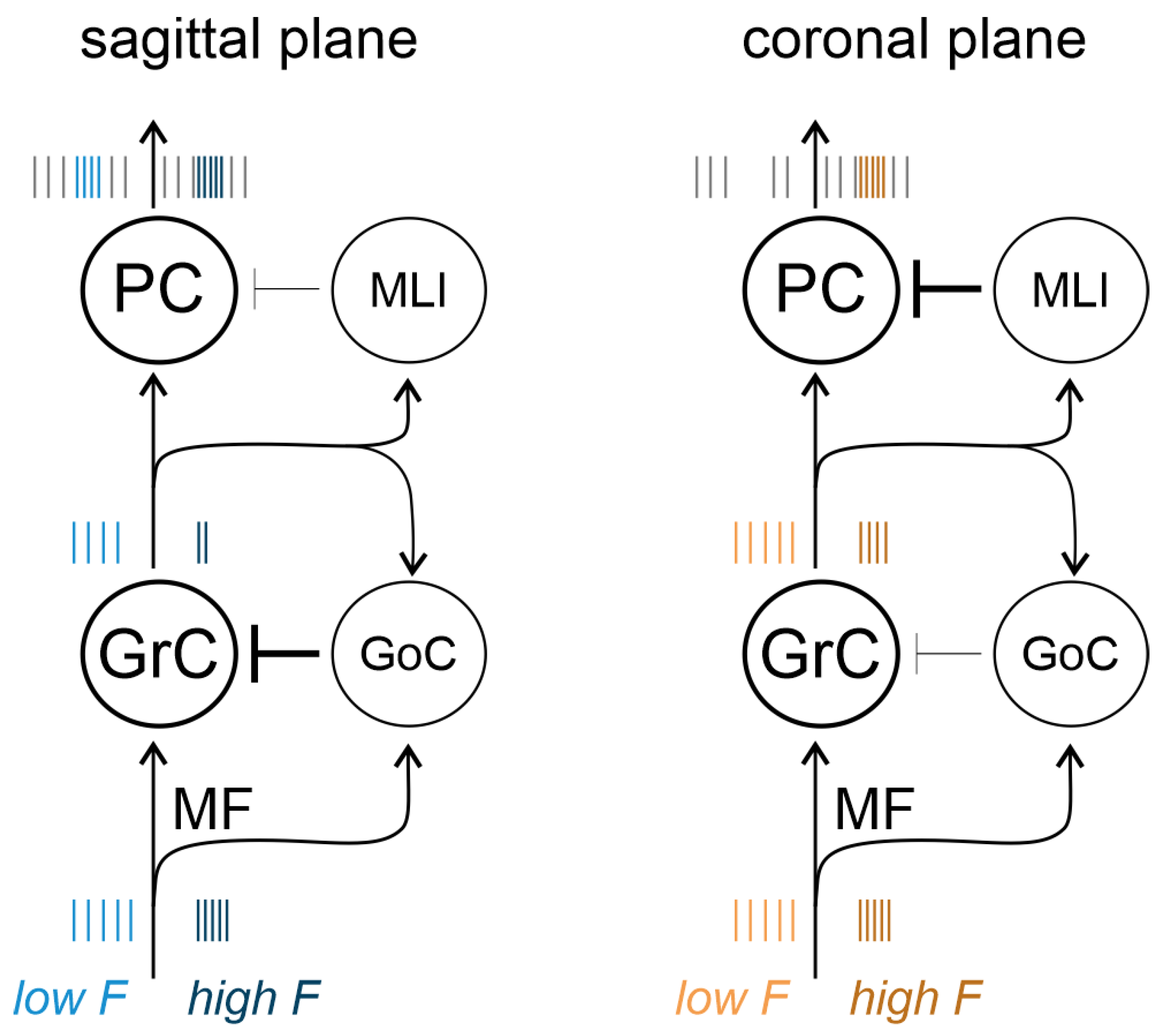
4. Discussion
4.1. Considerations on HD-MEA Recordings
4.2. Characterization of Spontaneous and Evoked Activity in Granular and PC Layers
 PC pathway).
PC pathway).4.3. Frequency-Dependent Responses in the Granular and PC Layers
5. Summary and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- D’Angelo, E. Chapter 6—Physiology of the cerebellum. In The Cerebellum: From Embryology to Diagnostic Investigations; Manto, M., Thierry AGM Huisman, Eds.; Elsevier: Amsterdam, The Netherlands, 2018; Volume 154, pp. 85–108. ISBN 0072-9752. [Google Scholar]
- Mapelli, L.; Solinas, S.; D’Angelo, E. Integration and regulation of glomerular inhibition in the cerebellar granular layer circuit. Front. Cell. Neurosci. 2014, 8, 55. [Google Scholar] [CrossRef] [PubMed]
- Prestori, F.; Mapelli, L.; D’Angelo, E. Diverse neuron properties and complex network dynamics in the cerebellar cortical inhibitory circuit. Front. Mol. Neurosci. 2019, 12, 267. [Google Scholar] [CrossRef]
- D’Angelo, E. The critical role of golgi cells in regulating spatio-temporal integration and plasticity at the cerebellum input stage. Front. Neurosci. 2009, 2, 35–46. [Google Scholar] [CrossRef]
- Tabuchi, S.; Gilmer, J.I.; Purba, K.; Person, A.L. Pathway-specific drive of cerebellar golgi cells reveals integrative rules of cortical inhibition. J. Neurosci. 2019, 39, 1169–1181. [Google Scholar] [CrossRef] [PubMed]
- Rizza, M.F.; Locatelli, F.; Masoli, S.; Sánchez-Ponce, D.; Muñoz, A.; Prestori, F.; D’Angelo, E. Stellate cell computational modeling predicts signal filtering in the molecular layer circuit of cerebellum. Sci. Rep. 2021, 11, 3873. [Google Scholar] [CrossRef] [PubMed]
- Rieubland, S.; Roth, A.; Häusser, M. Structured connectivity in cerebellar inhibitory networks. Neuron 2014, 81, 913–929. [Google Scholar] [CrossRef]
- Ango, F.; Wu, C.; Van Der Want, J.J.; Wu, P.; Schachner, M.; Huang, Z.J. Bergmann glia and the recognition molecule CHL1 organize GABAergic axons and direct innervation of purkinje cell dendrites. PLoS Biol. 2008, 6, 739–756. [Google Scholar] [CrossRef]
- D’Angelo, E.; Casali, S. Seeking a unified framework for cerebellar function and dysfunction: From circuit operations to cognition. Front. Neural Circuits 2012, 6, 116. [Google Scholar] [CrossRef] [PubMed]
- D’Angelo, E. The cerebellum gets social. Science 2019, 363, 229. [Google Scholar] [CrossRef]
- Palesi, F.; Tournier, J.D.; Calamante, F.; Muhlert, N.; Castellazzi, G.; Chard, D.; D’Angelo, E.; Wheeler-Kingshott, C.A.M. Contralateral cerebello-thalamo-cortical pathways with prominent involvement of associative areas in humans in vivo. Brain Struct. Funct. 2015, 220, 3369–3384. [Google Scholar] [CrossRef]
- Casiraghi, L.; Alahmadi, A.A.S.; Monteverdi, A.; Palesi, F.; Castellazzi, G.; Savini, G.; Friston, K.; Wheeler-Kingshott, C.A.M.G.; D’Angelo, E. I see your effort: Force-related BOLD effects in an extended action execution-observation network involving the cerebellum. Cereb. Cortex 2019, 29, 1351–1368. [Google Scholar] [CrossRef]
- Mapelli, L.; Soda, T.; D’Angelo, E.; Prestori, F. The cerebellar involvement in autism spectrum disorders: From the social brain to mouse models. Int. J. Mol. Sci. 2022, 23, 3894. [Google Scholar] [CrossRef] [PubMed]
- Schmahmann, J.D.; Guell, X.; Stoodley, C.J.; Halko, M.A. The theory and neuroscience of cerebellar cognition. Annu. Rev. Neurosci. 2019, 42, 337–364. [Google Scholar] [CrossRef] [PubMed]
- D’Angelo, E.; Koekkoek, S.K.E.; Lombardo, P.; Solinas, S.; Ros, E.; Garrido, J.; Schonewille, M.; De Zeeuw, C.I. Timing in the cerebellum: Oscillations and resonance in the granular layer. Neuroscience 2009, 162, 805–815. [Google Scholar] [CrossRef]
- Solinas, S.; Nieus, T.; D’Angelo, E. A realistic large-scale model of the cerebellum granular layer predicts circuit spatio-temporal filtering properties. Front. Cell. Neurosci. 2010, 4, 12. [Google Scholar] [CrossRef]
- Mapelli, J.; Gandolfi, D.; D’Angelo, E. High-pass filtering and dynamic gain regulation enhance vertical bursts transmission along the mossy fiber pathway of cerebellum. Front. Cell. Neurosci. 2010, 4, 14. [Google Scholar] [CrossRef] [PubMed]
- Gagliano, G.; Monteverdi, A.; Casali, S.; Laforenza, U.; Wheeler-Kingshott, C.A.M.G.; D’angelo, E.; Mapelli, L. Non-linear frequency dependence of neurovascular coupling in the cerebellar cortex implies vasodilation–Vasoconstriction competition. Cells 2022, 11, 1047. [Google Scholar] [CrossRef]
- Salmasi, M.; Loebel, A.; Glasauer, S.; Stemmler, M. Short-term synaptic depression can increase the rate of information transfer at a release site. PLoS Comput. Biol. 2019, 15, e1006666. [Google Scholar] [CrossRef]
- Grangeray-Vilmint, A.; Valera, A.M.; Kumar, A.; Isope, P. Short-term plasticity combines with excitation–Inhibition balance to expand cerebellar purkinje cell dynamic range. J. Neurosci. 2018, 38, 5153–5167. [Google Scholar] [CrossRef]
- Tognolina, M.; Monteverdi, A.; D’Angelo, E. Discovering microcircuit secrets with multi-spot imaging and electrophysiological recordings: The example of cerebellar network dynamics. Front. Cell. Neurosci. 2022, 16, 805670. [Google Scholar] [CrossRef]
- Rothman, J.S.; Cathala, L.; Steuber, V.; Silver, R.A. Synaptic depression enables neuronal gain control. Nature 2009, 457, 1015–1018. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; An, L.; Yuan, Y.; Pei, Q.; Wang, Q.; Liu, J.K. Modulation of the dynamics of cerebellar purkinje cells through the interaction of excitatory and inhibitory feedforward pathways. PLoS Comput. Biol. 2021, 17, e1008670. [Google Scholar] [CrossRef] [PubMed]
- Maffei, A.; Prestori, F.; Rossi, P.; Taglietti, V.; D’Angelo, E. Presynaptic current changes at the mossy fiber-granule cell synapse of cerebellum during LTP. J. Neurophysiol. 2002, 88, 627–638. [Google Scholar] [CrossRef]
- Mapelli, J.; D’Angelo, E. The spatial organization of long-term synaptic plasticity at the input stage of cerebellum. J. Neurosci. 2007, 27, 1285–1296. [Google Scholar] [CrossRef]
- Nieus, T.; D’Andrea, V.; Amin, H.; Di Marco, S.; Safaai, H.; Maccione, A.; Berdondini, L.; Panzeri, S. State-dependent representation of stimulus-evoked activity in high-density recordings of neural cultures. Sci. Rep. 2018, 8, 5578. [Google Scholar] [CrossRef] [PubMed]
- Hawkes, R. Purkinje cell stripes and long-term depression at the parallel fiber-purkinje cell synapse. Front. Syst. Neurosci. 2014, 8, 41. [Google Scholar] [CrossRef]
- Diwakar, S.; Lombardo, P.; Solinas, S.; Naldi, G.; D’Angelo, E. Local field potential modeling predicts dense activation in cerebellar granule cells clusters under LTP and LTD control. PLoS ONE 2011, 6, e21928. [Google Scholar] [CrossRef] [PubMed]
- D’Angelo, E.; Solinas, S.; Mapelli, J.; Gandolfi, D.; Mapelli, L.; Prestori, F. The cerebellar golgi cell and spatiotemporal organization of granular layer activity. Front. Neural Circuits 2013, 7, 93. [Google Scholar] [CrossRef]
- Korbo, L.; Andersen, B.B.; Ladefoged, O.; Møller, A. Total numbers of various cell types in rat cerebellar cortex estimated using an unbiased stereological method. Brain Res. 1993, 609, 262–268. [Google Scholar] [CrossRef]
- Masoli, S.; D’Angelo, E. Synaptic activation of a detailed purkinje cell model predicts voltage-dependent control of burst-pause responses in active dendrites. Front. Cell. Neurosci. 2017, 11, 278. [Google Scholar] [CrossRef]
- Cerminara, N.L.; Lang, E.J.; Sillitoe, R.V.; Apps, R. Redefining the cerebellar cortex as an assembly of non-uniform purkinje cell microcircuits. Nat. Rev. Neurosci. 2015, 16, 79–93. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Lin, Z.; Voges, K.; Ju, C.; Gao, Z.; Bosman, L.W.J.; Ruigrok, T.J.; Hoebeek, F.E.; De Zeeuw, C.I.; Schonewille, M. Cerebellar modules operate at different frequencies. Elife 2014, 2014, e02536. [Google Scholar] [CrossRef] [PubMed]
- Sultan, F. Distribution of mossy fibre rosettes in the cerebellum of cat and mice: Evidence for a parasagittal organization at the single fibre level. Eur. J. Neurosci. 2001, 13, 2123–2130. [Google Scholar] [CrossRef]
- Walter, J.T.; Dizon, M.J.; Khodakhah, K. The functional equivalence of ascending and parallel fiber inputs in cerebellar computation. J. Neurosci. 2009, 29, 8462–8473. [Google Scholar] [CrossRef] [PubMed]
- Sola, E.; Prestori, F.; Rossi, P.; Taglietti, V.; D’Angelo, E. Increased neurotransmitter release during long-term potentiation at mossy fibre-granule cell synapses in rat cerebellum. J. Physiol. 2004, 557, 843–861. [Google Scholar] [CrossRef]
- Nieus, T.R.; Mapelli, L.; D’Angelo, E. Regulation of output spike patterns by phasic inhibition in cerebellar granule cells. Front. Cell. Neurosci. 2014, 8, 246. [Google Scholar] [CrossRef]
- D’Errico, A.; Prestori, F.; D’Angelo, E. Differential induction of bidirectional long-term changes in neurotransmitter release by frequency-coded patterns at the cerebellar input. J. Physiol. 2009, 587, 5843–5857. [Google Scholar] [CrossRef]
- Gao, Z.; Van Beugen, B.J.; De Zeeuw, C.I. Distributed synergistic plasticity and cerebellar learning. Nat. Rev. Neurosci. 2012, 13, 619–635. [Google Scholar] [CrossRef] [PubMed]
- Wall, M.J. Short-term synaptic plasticity during development of rat mossy fibre to granule cell synapses. Eur. J. Neurosci. 2005, 21, 2149–2158. [Google Scholar] [CrossRef]
- DiGregorio, D.A.; Rothman, J.S.; Nielsen, T.A.; Silver, R.A. Desensitization properties of AMPA receptors at the cerebellar mossy fiber-granule cell synapse. J. Neurosci. 2007, 27, 8344–8357. [Google Scholar] [CrossRef]
- Mapelli, L.; Rossi, P.; Nieus, T.; D’Angelo, E. Tonic activation of GABAB receptors reduces release probability at inhibitory connections in the cerebellar glomerulus. J. Neurophysiol. 2009, 101, 3089–3099. [Google Scholar] [CrossRef] [PubMed]
- Carter, A.G.; Regehr, W.G. Prolonged synaptic currents and glutamate spillover at the parallel fiber to stellate cell synapse. J. Neurosci. 2000, 20, 4423–4434. [Google Scholar] [CrossRef] [PubMed]
- Bao, J.; Reim, K.; Sakaba, T. Target-dependent feedforward inhibition mediated by short-term synaptic plasticity in the cerebellum. J. Neurosci. 2010, 30, 8171–8179. [Google Scholar] [CrossRef] [PubMed]
- Dorgans, K.; Demai, V.; Bailly, Y.; Poulain, B.; Isope, P.; Doussau, F. Short-term plasticity at cerebellar granule cell to molecular layer interneuron synapses expands information processing. Elife 2019, 8, e41586. [Google Scholar] [CrossRef] [PubMed]
- Szapiro, G.; Barbour, B. Multiple climbing fibers signal to molecular layer interneurons exclusively via glutamate spillover. Nat. Neurosci. 2007, 10, 735–742. [Google Scholar] [CrossRef] [PubMed]
- De Schepper, R.; Geminiani, A.; Masoli, S.; Rizza, M.F.; Antonietti, A.; Casellato, C.; D’Angelo, E. Model simulations unveil the structure-function-dynamics relationship of the cerebellar cortical microcircuit. Commun. Biol. 2022, 5, 1240. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SAGITTAL | CORONAL | ||
|---|---|---|---|
| n = 9 | n = 9 | tot | |
| ch granular layer | 804 | 1649 | 2453 |
| ch Purkinje cells | 1859 | 1809 | 3668 |
| detected units | 858 | 1095 | 1953 |
| A | ||||
| 6 Hz | 20 Hz | 50 Hz | 100 Hz | |
| N2a | −13.60 ± 0.99 | −15.58 ± 1.45 | −19.87 ± 1.41 | −27.37 ± 1.92 |
| N2b↑ | 12.33 ± 1.68 | 13.89 ± 2.01 | 17.13 ± 2.83 | 20.09 ± 3.18 |
| N2b↓ | −10.59 ± 2.42 | −13.36 ± 3.08 | −17.34 ± 1.85 | −13.48 ± 2.10 |
| B | ||||
| 6 Hz | 20 Hz | 50 Hz | 100 Hz | |
| %ch N2a | 75.82 ± 4.63 | 72.34 ± 6.48 | 83.92 ± 3.85 | 88.82 ± 3.03 |
| %ch N2b ↑ | 11.83 ± 3.61 | 16.94 ± 5.74 | 25.74 ± 5.88 | 40.09 ± 9.05 |
| %ch N2b ↓ | 16.57 ± 9.41 | 15.15 ± 8.86 | 22.81 ± 7.61 | 13.56 ± 4.49 |
| A | ||||
| 6 Hz | 20 Hz | 50 Hz | 100 Hz | |
| N2a | −13.44 ± 0.84 | −14.92 ± 1.26 | −17.56 ± 0.89 | −23.93 ± 1.46 |
| N2b + | 14.80 ± 1.44 | 14.26 ± 1.65 | 18.23 ± 1.69 | 17.62 ± 1.41 |
| N2b − | −10.63 ± 1.71 | −11.55 ± 2.19 | −6.53 ± 2.12 | −10.28 ± 2.30 |
| B | ||||
| 6 Hz | 20 Hz | 50 Hz | 100 Hz | |
| %ch N2a | 53.96 ± 7.18 | 55.47 ± 6.85 | 61.59 ± 6.73 | 78.94 ± 5.37 |
| %ch N2b ↑ | 18.01 ± 6.02 | 19.77 ± 4.94 | 41.60 ± 4.43 | 39.41 ± 6.78 |
| %ch N2b ↓ | 4.75 ± 1.60 | 2.23 ± 0.79 | 0.74 ± 0.29 | 2.60 ± 1.12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monteverdi, A.; Di Domenico, D.; D’Angelo, E.; Mapelli, L. Anisotropy and Frequency Dependence of Signal Propagation in the Cerebellar Circuit Revealed by High-Density Multielectrode Array Recordings. Biomedicines 2023, 11, 1475. https://doi.org/10.3390/biomedicines11051475
Monteverdi A, Di Domenico D, D’Angelo E, Mapelli L. Anisotropy and Frequency Dependence of Signal Propagation in the Cerebellar Circuit Revealed by High-Density Multielectrode Array Recordings. Biomedicines. 2023; 11(5):1475. https://doi.org/10.3390/biomedicines11051475
Chicago/Turabian StyleMonteverdi, Anita, Danila Di Domenico, Egidio D’Angelo, and Lisa Mapelli. 2023. "Anisotropy and Frequency Dependence of Signal Propagation in the Cerebellar Circuit Revealed by High-Density Multielectrode Array Recordings" Biomedicines 11, no. 5: 1475. https://doi.org/10.3390/biomedicines11051475
APA StyleMonteverdi, A., Di Domenico, D., D’Angelo, E., & Mapelli, L. (2023). Anisotropy and Frequency Dependence of Signal Propagation in the Cerebellar Circuit Revealed by High-Density Multielectrode Array Recordings. Biomedicines, 11(5), 1475. https://doi.org/10.3390/biomedicines11051475