
Identification of Novel Biomarkers of Spinal Muscular Atrophy and Therapeutic Response by Proteomic and Metabolomic Profiling of Human Biological Fluid Samples
 , ,
, , 
Abstract
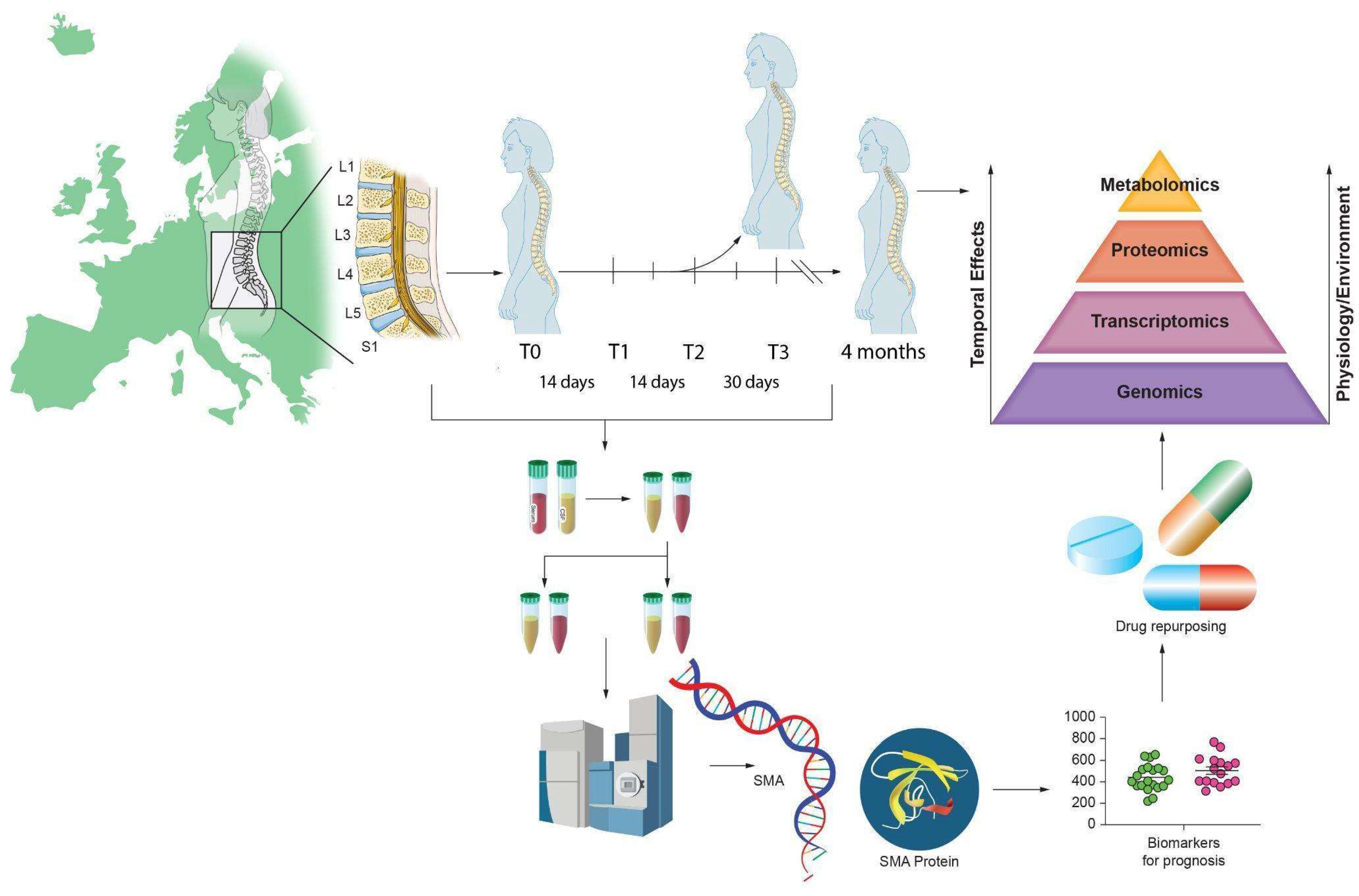
1. Introduction
2. Blood Proteomic Studies of SMA
2.1. Identification of Proteins, Metabolites, and Transcripts in Body Fluids of Untreated Patients: Biomarkers for SMA Study (BforSMA)
2.2. DPP4
2.3. OPN
2.4. TET
2.5. Fetuin-A
2.6. VTN
2.7. Multi-Analyte Panel Assay (SMA-MAP) of BforSMA
2.8. Cerebrospinal Fluid Proteomic Profiles in SMA Patients Treated with Nusinersen
CSF Proteomic by Mass Spectrometry
2.9. NPTX1
2.10. SEMA7A
2.11. CPE
2.12. COL6A1
2.13. CDH18
2.14. CSF Proteomic Profiles with 2D-PAGE
2.15. Apolipoproteins
2.16. TTR
2.17. Overall, Lipoproteins and TTR May Be Useful Biomarkers for Monitoring Patient Response and Disease Progression
Carbonyl Group Patterns
2.18. Combination of Untargeted and Targeted Proteomics of the CSF of SMA Subjects before and after Treatment
3. Neurometabolic Patterns in CSF before and after Treatment of SMA
3.1. 1H-NMR Metabolomes of Three Samples (Urine, Serum, and CSF)
3.2. NMR Spectroscopy: A Second Study
4. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lefebvre, S.; Bürglen, L.; Reboullet, S.; Clermont, O.; Burlet, P.; Viollet, L.; Benichou, B.; Cruaud, C.; Millasseau, P.; Zeviani, M.; et al. Identification and characterization of a spinal muscular atrophy-determining gene. Cell 1995, 80, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Mercuri, E.; Sumner, C.J.; Muntoni, F.; Darras, B.T.; Finkel, R.S. Spinal muscular atrophy. Nat. Rev. Dis. Prim. 2022, 8, 52. [Google Scholar] [CrossRef] [PubMed]
- Paushkin, S.; Gubitz, A.K.; Massenet, S.; Dreyfuss, G. The SMN complex, an assemblyosome of ribonucleoproteins. Curr. Opin. Cell Biol. 2002, 14, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Shpargel, K.B.; Matera, A.G. Gemin proteins are required for efficient assembly of Sm-class ribonucleoproteins. Proc. Natl. Acad. Sci. USA 2005, 102, 17372–17377. [Google Scholar] [CrossRef] [PubMed]
- Faravelli, I.; Nizzardo, M.; Comi, G.P.; Corti, S. Spinal muscular atrophy—Recent therapeutic advances for an old challenge. Nat. Rev. Neurol. 2015, 11, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Faravelli, I.; Riboldi, G.M.; Rinchetti, P.; Lotti, F. The SMN Complex at the Crossroad between RNA Metabolism and Neurodegeneration. Int. J. Mol. Sci. 2023, 24, 2247. [Google Scholar] [CrossRef]
- Wirth, B.; Karakaya, M.; Kye, M.J.; Mendoza-Ferreira, N. Twenty-Five Years of Spinal Muscular Atrophy Research: From Phenotype to Genotype to Therapy, and What Comes Next. Annu. Rev. Genom. Hum. Genet. 2020, 21, 231–261. [Google Scholar] [CrossRef]
- Ripolone, M.; Ronchi, D.; Violano, R.; Vallejo, D.; Fagiolari, G.; Barca, E.; Lucchini, V.; Colombo, I.; Villa, L.; Berardinelli, A.; et al. Impaired Muscle Mitochondrial Biogenesis and Myogenesis in Spinal Muscular Atrophy. JAMA Neurol. 2015, 72, 666–675. [Google Scholar] [CrossRef]
- Zilio, E.; Piano, V.; Wirth, B. Mitochondrial Dysfunction in Spinal Muscular Atrophy. Int. J. Mol. Sci. 2022, 23, 10878. [Google Scholar] [CrossRef]
- Lorson, C.L.; Hahnen, E.; Androphy, E.J.; Wirth, B. A single nucleotide in the SMN gene regulates splicing and is responsible for spinal muscular atrophy. Proc. Natl. Acad. Sci. USA 1999, 96, 6307–6311. [Google Scholar] [CrossRef]
- Lunn, M.R.; Wang, C.H. Spinal muscular atrophy. Lancet 2008, 371, 2120–2133. [Google Scholar] [CrossRef] [PubMed]
- Gama-Carvalho, M.; Garcia-Vaquero, M.L.; Pinto, F.R.; Besse, F.; Weis, J.; Voigt, A.; Schulz, J.B.; Rivas, J.D.L. Linking amyotrophic lateral sclerosis and spinal muscular atrophy through RNA-transcriptome homeostasis: A genomics perspective. J. Neurochem. 2017, 141, 12–30. [Google Scholar] [CrossRef]
- Finkel, R.S.; Chiriboga, C.A.; Vajsar, J.; Day, J.W.; Montes, J.; De Vivo, D.C.; Yamashita, M.; Rigo, F.; Hung, G.; Schneider, E.; et al. Treatment of infantile-onset spinal muscular atrophy with nusinersen: A phase 2, open-label, dose-escalation study. Lancet 2016, 388, 3017–3026. [Google Scholar] [CrossRef] [PubMed]
- Finkel, R.S.; Mercuri, E.; Darras, B.T.; Connolly, A.M.; Kuntz, N.L.; Kirschner, J.; Chiriboga, C.A.; Saito, K.; Servais, L.; Tizzano, E.; et al. Nusinersen versus Sham Control in Infantile-Onset Spinal Muscular Atrophy. N. Engl. J. Med. 2017, 377, 1723–1732. [Google Scholar] [CrossRef] [PubMed]
- Finkel, R.S.; Chiriboga, C.A.; Vajsar, J.; Day, J.W.; Montes, J.; De Vivo, D.C.; Bishop, K.M.; Foster, R.; Liu, Y.; Ramirez-Schrempp, D.; et al. Treatment of infantile-onset spinal muscular atrophy with nusinersen: Final report of a phase 2, open-label, multicentre, dose-escalation study. Lancet Child Adolesc. Health 2021, 5, 491–500. [Google Scholar] [CrossRef] [PubMed]
- Day, J.W.; Finkel, R.S.; Chiriboga, C.A.; Connolly, A.M.; Crawford, T.O.; Darras, B.T.; Iannaccone, S.T.; Kuntz, N.L.; Peña, L.D.; Shieh, P.B. Onasemnogene abeparvovec gene therapy for symptomatic infantile-onset spinal muscular atrophy in patients with two copies of SMN2 (STR1VE): An open-label, single-arm, multicentre, phase 3 trial. Lancet Neurol. 2021, 20, 284–293. [Google Scholar] [CrossRef]
- Darras, B.T.; Masson, R.; Mazurkiewicz-Bełdzińska, M.; Rose, K.; Xiong, H.; Zanoteli, E.; Baranello, G.; Bruno, C.; Vlodavets, D.; Wang, Y.; et al. Risdiplam-Treated Infants with Type 1 Spinal Muscular Atrophy versus Historical Controls. N. Engl. J. Med. 2021, 385, 427–435. [Google Scholar] [CrossRef] [PubMed]
- Masson, R.; Mazurkiewicz-Bełdzińska, M.; Rose, K.; Servais, L.; Xiong, H.; Zanoteli, E.; Baranello, G.; Bruno, C.; Day, J.W.; Deconinck, N.; et al. Safety and efficacy of risdiplam in patients with type 1 spinal muscular atrophy (FIREFISH part 2): Secondary analyses from an open-label trial. Lancet Neurol. 2022, 21, 1110–1119. [Google Scholar] [CrossRef] [PubMed]
- Bennett, C.F.; Baker, B.F.; Pham, N.; Swayze, E.; Geary, R.S. Pharmacology of antisense drugs. Annu. Rev. Pharmacol. Toxicol. 2017, 57, 81–105. [Google Scholar] [CrossRef]
- Wurster, C.D.; Ludolph, A.C. Antisense oligonucleotides in neurological disorders. Ther. Adv. Neurol. Disord. 2018, 11, 1756286418776932. [Google Scholar] [CrossRef]
- Pera, M.C.; Coratti, G.; Bovis, F.; Pane, M.; Pasternak, A.; Montes, J.; Sansone, V.A.; Young, S.D.; Duong, T.; Messina, S.; et al. Nusinersen in pediatric and adult patients with type III spinal muscular atrophy. Ann. Clin. Transl. Neurol. 2021, 8, 1622–1634. [Google Scholar] [CrossRef] [PubMed]
- Pino, M.G.; Rich, K.A.; Kolb, S.J. Update on Biomarkers in Spinal Muscular Atrophy. Biomark. Insights 2021, 16, 11772719211035643. [Google Scholar] [CrossRef] [PubMed]
- Gavriilaki, M.; Moschou, M.; Papaliagkas, V.; Notas, K.; Chatzikyriakou, E.; Zafeiridou, G.; Papagiannopoulos, S.; Arnaoutoglou, M.; Kimiskidis, V.K. Biomarkers of disease progression in adolescents and adults with 5q spinal muscular atrophy: A systematic review and meta-analysis. Neuromuscul. Disord. 2022, 32, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Dangouloff, T.; Burghes, A.; Tizzano, E.F.; Servais, L.; Bertini, E.; Boemer, F.; Hiligsmann, M.; Mueller-Felber, W.; Tiziano, D.; Young, P. 244th ENMC international workshop: Newborn screening in spinal muscular atrophy 10–12 May 2019, Hoofdorp, The Netherlands. Neuromuscul. Disord. 2020, 30, 93–103. [Google Scholar] [CrossRef]
- Bonanno, S.; Marcuzzo, S.; Malacarne, C.; Giagnorio, E.; Masson, R.; Zanin, R.; Arnoldi, M.T.; Andreetta, F.; Simoncini, O.; Venerando, A.; et al. Circulating MyomiRs as Potential Biomarkers to Monitor Response to Nusinersen in Pediatric SMA Patients. Biomedicines 2020, 8, 21. [Google Scholar] [CrossRef]
- Bonanno, S.; Cavalcante, P.; Salvi, E.; Giagnorio, E.; Malacarne, C.; Cattaneo, M.; Andreetta, F.; Venerando, A.; Pensato, V.; Gellera, C.; et al. Identification of a cytokine profile in serum and cerebrospinal fluid of pediatric and adult spinal muscular atrophy patients and its modulation upon nusinersen treatment. Front. Cell. Neurosci. 2022, 16, 982760. [Google Scholar] [CrossRef]
- Schumacher-Schuh, A.; Bieger, A.; Borelli, W.V.; Portley, M.K.; Awad, P.S.; Bandres-Ciga, S. Advances in Proteomic and Metabolomic Profiling of Neurodegenerative Diseases. Front. Neurol. 2022, 12, 2545. [Google Scholar] [CrossRef]
- Finkel, R.S.; Crawford, T.O.; Swoboda, K.J.; Kaufmann, P.; Juhasz, P.; Li, X.; Guo, Y.; Li, R.H.; Trachtenberg, F.; Forrest, S.J.; et al. Candidate Proteins, Metabolites and Transcripts in the Biomarkers for Spinal Muscular Atrophy (BforSMA) Clinical Study. PLoS ONE 2012, 7, e35462. [Google Scholar] [CrossRef]
- Kessler, T.; Latzer, P.; Schmid, D.; Warnken, U.; Saffari, A.; Ziegler, A.; Kollmer, J.; Möhlenbruch, M.; Ulfert, C.; Herweh, C.; et al. Cerebrospinal fluid proteomic profiling in nusinersen-treated patients with spinal muscular atrophy. J. Neurochem. 2020, 153, 650–661. [Google Scholar] [CrossRef]
- Bianchi, L.; Sframeli, M.; Vantaggiato, L.; Vita, G.L.; Ciranni, A.; Polito, F.; Oteri, R.; Gitto, E.; Di Giuseppe, F.; Angelucci, S.; et al. Nusinersen Modulates Proteomics Profiles of Cerebrospinal Fluid in Spinal Muscular Atrophy Type 1 Patients. Int. J. Mol. Sci. 2021, 22, 4329. [Google Scholar] [CrossRef]
- Schorling, D.C.; Kölbel, H.; Hentschel, A.; Pechmann, A.; Meyer, N.; Wirth, B.; Rombo, R.; Sickmann, A.; Kirschner, J.; Schara-Schmidt, U.; et al. Cathepsin D as biomarker in cerebrospinal fluid of nusinersen-treated patients with spinal muscular atrophy. Eur. J. Neurol. 2022, 29, 2084–2096. [Google Scholar] [CrossRef] [PubMed]
- Deutsch, L.; Osredkar, D.; Plavec, J.; Stres, B. Spinal Muscular Atrophy after Nusinersen Therapy: Improved Physiology in Pediatric Patients with No Significant Change in Urine, Serum, and Liquor 1H-NMR Metabolomes in Comparison to an Age-Matched, Healthy Cohort. Metabolites 2021, 11, 206. [Google Scholar] [CrossRef] [PubMed]
- Errico, F.; Marino, C.; Grimaldi, M.; Nuzzo, T.; Bassareo, V.; Valsecchi, V.; Panicucci, C.; Di Schiavi, E.; Mazza, T.; Bruno, C.; et al. Nusinersen Induces Disease-Severity-Specific Neurometabolic Effects in Spinal Muscular Atrophy. Biomolecules 2022, 12, 1431. [Google Scholar] [CrossRef] [PubMed]
- Ward, K.; Mulder, E.; Frings-Meuthen, P.; O’Gorman, D.J.; Cooper, D. Fetuin-A as a potential biomarker of metabolic variability following 60 days of bed rest. Front. Physiol. 2020, 11, 573581. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Wang, X.; Xue, X. Therapeutic Perspectives of CD26 Inhibitors in Imune-Mediated Diseases. Molecules 2022, 27, 4498. [Google Scholar] [CrossRef] [PubMed]
- Stalmach, A.; Boehm, I.; Fernandes, M.; Rutter, A.; Skipworth, R.J.E.; Husi, H. Gene Ontology (GO)-Driven Inference of Candidate Proteomic Markers Associated with Muscle Atrophy Conditions. Molecules 2022, 27, 5514. [Google Scholar] [CrossRef] [PubMed]
- Takada, S.; Masaki, Y.; Kinugawa, S.; Matsumoto, J.; Furihata, T.; Mizushima, W.; Kadoguchi, T.; Fukushima, A.; Homma, T.; Takahashi, M.; et al. Dipeptidyl peptidase-4 inhibitor improved exercise capacity and mitochondrial biogenesis in mice with heart failure via activation of glucagon-like peptide-1 receptor signalling. Cardiovasc. Res. 2016, 111, 338–347. [Google Scholar] [CrossRef] [PubMed]
- Ishii, S.; Nagai, Y.; Kato, H.; Fukuda, H.; Tanaka, Y. Effect of the Dipeptidyl Peptidase-4 Inhibitor Sitagliptin on Muscle Mass and the Muscle/Fat Ratio in Patients with Type 2 Diabetes. J. Clin. Med. Res. 2020, 12, 122–126. [Google Scholar] [CrossRef]
- Kalaitzoglou, E.; Fowlkes, J.L.; Popescu, I.; Thrailkill, K.M. Diabetes pharmacotherapy and effects on the musculoskeletal system. Diabetes/Metabolism Res. Rev. 2019, 35, e3100. [Google Scholar] [CrossRef]
- Angelopoulou, E.; Piperi, C. DPP-4 inhibitors: A promising therapeutic approach against Alzheimer’s disease. Ann. Transl. Med. 2018, 6, 255. [Google Scholar] [CrossRef]
- Bastos, A.C.S.D.F.; Gomes, A.V.P.; Silva, G.R.; Emerenciano, M.; Ferreira, L.B.; Gimba, E.R.P. The Intracellular and Secreted Sides of Osteopontin and Their Putative Physiopathological Roles. Int. J. Mol. Sci. 2023, 24, 2942. [Google Scholar] [CrossRef] [PubMed]
- Bello, L.; Pegoraro, E. The “Usual Suspects”: Genes for Inflammation, Fibrosis, Regeneration, and Muscle Strength Modify Duchenne Muscular Dystrophy. J. Clin. Med. 2019, 8, 649. [Google Scholar] [CrossRef] [PubMed]
- Cappellano, G.; Vecchio, D.; Magistrelli, L.; Clemente, N.; Raineri, D.; Mazzucca, C.B.; Virgilio, E.; Dianzani, U.; Chiocchetti, A.; Comi, C. The Yin-Yang of osteopontin in nervous system diseases: Damage versus repair. Neural Regen. Res. 2021, 16, 1131–1137. [Google Scholar] [CrossRef]
- Wewer, U.M.; Iba, K.; Durkin, M.E.; Nielsen, F.C.; Loechel, F.; Gilpin, B.J.; Kuang, W.; Engvall, E.; Albrechtsen, R. Tetranectin Is a Novel Marker for Myogenesis during Embryonic Development, Muscle Regeneration, and Muscle Cell Differentiationin Vitro. Dev. Biol. 1998, 200, 247–259. [Google Scholar] [CrossRef] [PubMed]
- McDonald, K.; Glezeva, N.; Collier, P.; O’reilly, J.; O’connell, E.; Tea, I.; Russell-Hallinan, A.; Tonry, C.; Pennington, S.; Gallagher, J.; et al. Tetranectin, a potential novel diagnostic biomarker of heart failure, is expressed within the myocardium and associates with cardiac fibrosis. Sci. Rep. 2020, 10, 7507. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Wang, E.; Hu, R.; Sun, Y.; Zhang, L.; Jiang, J.; Zhang, Y.; Jiang, H. Tetranectin gene deletion induces Parkinson’s disease by enhancing neuronal apoptosis. Biochem. Biophys. Res. Commun. 2015, 468, 400–407. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, S.; Mondal, S.A.; Kumar, M.; Dutta, D. Proinflammatory and Antiinflammatory Attributes of Fetu Iν-A: A Novel Hepatokine Modulating Cardiovascular and Glycemic Outcomes in Metabolic Syndrome. Endocr. Pr. 2014, 20, 1345–1351. [Google Scholar] [CrossRef]
- Ruzha, Y.; Ni, J.; Quan, Z.; Li, H.; Qing, H. Role of Vitronectin and Its Receptors in Neuronal Function and Neurodegenerative Diseases. Int. J. Mol. Sci. 2022, 23, 12387. [Google Scholar] [CrossRef]
- Kobayashi, D.T.; Shi, J.; Stephen, L.; Ballard, K.L.; Dewey, R.; Mapes, J.; Chung, B.; McCarthy, K.; Swoboda, K.J.; Crawford, T.O.; et al. SMA-MAP: A Plasma Protein Panel for Spinal Muscular Atrophy. PLoS ONE 2013, 8, e60113. [Google Scholar] [CrossRef]
- Arnold, W.D.; Duque, S.; Iyer, C.C.; Zaworski, P.; McGovern, V.L.; Taylor, S.J.; von Herrmann, K.M.; Kobayashi, D.T.; Chen, K.S.; Kolb, S.J.; et al. Normalization of Patient-Identified Plasma Biomarkers in SMNΔ7 Mice following Postnatal SMN Restoration. PLoS ONE 2016, 11, e0167077. [Google Scholar] [CrossRef]
- Kolb, S.J.; Coffey, C.S.; Yankey, J.W.; Krosschell, K.; Arnold, W.D.; Rutkove, S.B.; Swoboda, K.J.; Reyna, S.P.; Sakonju, A.; Darras, B.T.; et al. Baseline results of the Neuro NEXT spinal muscular atrophy infant biomarker study. Ann. Clin. Transl. Neurol. 2016, 3, 132–145. [Google Scholar] [CrossRef] [PubMed]
- Antipova, D.; Bandopadhyay, R. Expression of DJ-1 in Neurodegenerative Disorders, DJ-1/PARK7 Protein: Parkinson’s Disease. Cancer Oxidative Stress-Induc. Dis. 2017, 1037, 25–43. [Google Scholar] [CrossRef]
- Panicker, N.; Ge, P.; Dawson, V.L.; Dawson, T.M. The cell biology of Parkinson’s disease. J. Cell Biol. 2021, 220, e202012095. [Google Scholar] [CrossRef] [PubMed]
- Hong, Z.; Shi, M.; Chung, K.A.; Quinn, J.F.; Peskind, E.R.; Galasko, D.; Jankovic, J.; Zabetian, C.P.; Leverenz, J.B.; Baird, G. DJ-1 and α-synuclein in human cerebrospinal fluid as biomarkers of Parkinson’s disease. Brain 2010, 133, 713–726. [Google Scholar] [CrossRef] [PubMed]
- Schaukowitch, K.; Reese, A.L.; Kim, S.-K.; Kilaru, G.; Joo, J.-Y.; Kavalali, E.T.; Kim, T.-K. An Intrinsic Transcriptional Program Underlying Synaptic Scaling during Activity Suppression. Cell Rep. 2017, 18, 1512–1526. [Google Scholar] [CrossRef]
- Sogorb-Esteve, A.; Nilsson, J.; Swift, I.J.; Heller, C.; Bocchetta, M.; Russell, L.L.; Peakman, G.; Convery, R.S.; van Swieten, J.C.; Seelaar, H.; et al. Differential impairment of cerebrospinal fluid synaptic biomarkers in the genetic forms of frontotemporal dementia. Alzheimer’s Res. Ther. 2022, 14, 118. [Google Scholar] [CrossRef]
- Pasterkamp, R.J.; Kolk, S.M.; Hellemons, A.J.; Kolodkin, A.L. Expression patterns of semaphorin7A and plexinC1during rat neural development suggest roles in axon guidance and neuronal migration. BMC Dev. Biol. 2007, 7, 98. [Google Scholar] [CrossRef]
- Cantó, E.; Tintoré, M.; Villar, L.M.; Borrás, E.; Álvarez-Cermeño, J.C.; Chiva, C.; Sabidó, E.; Rovira, A.; Montalban, X.; Comabella, M. Validation of semaphorin 7A and ala-β-his-dipeptidase as biomarkers associated with the conversion from clinically isolated syndrome to multiple sclerosis. J. Neuroinflammation 2014, 11, 181. [Google Scholar] [CrossRef]
- Arbeille, E.; Reynaud, F.; Sanyas, I.; Bozon, M.; Kindbeiter, K.; Causeret, F.; Pierani, A.; Falk, J.; Moret, F.; Castellani, V. Cerebrospinal fluid-derived Semaphorin3B orients neuroepithelial cell divisions in the apicobasal axis. Nat. Commun. 2015, 6, 6366. [Google Scholar] [CrossRef]
- Gerstmann, K.; Kindbeiter, K.; Telley, L.; Bozon, M.; Reynaud, F.; Théoulle, E.; Charoy, C.; Jabaudon, D.; Moret, F.; Castellani, V. A balance of noncanonical Semaphorin signaling from the cerebrospinal fluid regulates apical cell dynamics during corticogenesis. Sci. Adv. 2022, 8, eabo4552. [Google Scholar] [CrossRef]
- Mastrantonio, R.; You, H.; Tamagnone, L. Semaphorins as emerging clinical biomarkers and therapeutic targets in cancer. Theranostics 2021, 11, 3262–3277. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Yang, X.; Loh, Y.P. Neurotrophic, gene regulation, and cognitive functions of carboxypeptidase E-neurotrophic factor-α1 and its variants. Front. Neurosci. 2019, 13, 243. [Google Scholar] [CrossRef] [PubMed]
- Bushby, K.M.; Collins, J.; Hicks, D. Collagen type VI myopathies. In Progress in Heritable Soft Connective Tissue Diseases; Springer: Berlin/Heidelberg, Germany, 2014; pp. 185–199. [Google Scholar] [CrossRef]
- Redies, C.; Hertel, N.; Hübner, C.A. Cadherins and neuropsychiatric disorders. Brain Res. 2012, 1470, 130–144. [Google Scholar] [CrossRef]
- Güzel, C.; Berg, C.B.V.D.; Koopman, S.; Van Krugten, R.J.; Stoop, M.; Stingl, C.; Duvekot, J.J.; Luider, T.M. Cerebrospinal Fluid of Preeclamptic and Normotensive Pregnant Women Compared to Nonpregnant Women Analyzed with Mass Spectrometry. ACS Omega 2020, 5, 32256–32266. [Google Scholar] [CrossRef] [PubMed]
- Fonteh, A.N.; Harrington, R.J.; Huhmer, A.F.; Biringer, R.G.; Riggins, J.N.; Harrington, M.G. Identification of Disease Markers in Human Cerebrospinal Fluid Using Lipidomic and Proteomic Methods. Dis. Markers 2006, 22, 39–64. [Google Scholar] [CrossRef]
- Spagnuolo, M.S.; Donizetti, A.; Iannotta, L.; Aliperti, V.; Cupidi, C.; Bruni, A.C.; Cigliano, L. Brain-derived neurotrophic factor modulates cholesterol homeostasis and Apolipoprotein E synthesis in human cell models of astrocytes and neurons. J. Cell. Physiol. 2018, 233, 6925–6943. [Google Scholar] [CrossRef]
- Ahmed, S.; Pande, A.H.; Sharma, S.S. Therapeutic potential of ApoE-mimetic peptides in CNS disorders: Current perspective. Exp. Neurol. 2022, 353, 114051. [Google Scholar] [CrossRef]
- Filou, S.; Lhomme, M.; Karavia, E.A.; Kalogeropoulou, C.; Theodoropoulos, V.; Zvintzou, E.; Sakellaropoulos, G.C.; Petropoulou, P.-I.; Constantinou, C.; Kontush, A.; et al. Distinct Roles of Apolipoproteins A1 and E in the Modulation of High-Density Lipoprotein Composition and Function. Biochemistry 2016, 55, 3752–3762. [Google Scholar] [CrossRef]
- Johansson, P.; Almqvist, E.G.; Bjerke, M.; Wallin, A.; Johansson, J.-O.; Andreasson, U.; Blennow, K.; Zetterberg, H.; Svensson, J. Reduced Cerebrospinal Fluid Concentration of Apolipoprotein A-I in Patients with Alzheimer’s Disease. J. Alzheimer’s Dis. 2017, 59, 1017–1026. [Google Scholar] [CrossRef]
- Liu, Y.; Song, J.-H.; Xu, W.; Hou, X.-H.; Li, J.-Q.; Yu, J.-T.; Tan, L.; Chi, S.; Initiative, A.D.N. The Associations of Cerebrospinal Fluid ApoE and Biomarkers of Alzheimer’s Disease: Exploring Interactions with Sex. Front. Neurosci. 2021, 15, 633576. [Google Scholar] [CrossRef]
- Yin, Y.; Wang, Z. ApoE and neurodegenerative diseases in aging. In Aging and Aging-Related Diseases: Mechanisms and Interventions; Springer: Berlin/Heidelberg, Germany, 2018; pp. 77–92. [Google Scholar] [CrossRef]
- Yang, A.; Kantor, B.; Chiba-Falek, O. APOE: The New Frontier in the Development of a Therapeutic Target towards Precision Medicine in Late-Onset Alzheimer’s. Int. J. Mol. Sci. 2021, 22, 1244. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Khan, S.; Rahman, S.; Singh, L.R. The Extracellular Protein, Transthyretin Is an Oxidative Stress Biomarker. Front. Physiol. 2019, 10, 5. [Google Scholar] [CrossRef] [PubMed]
- Alshehri, B.; Pagnin, M.; Lee, J.Y.; Petratos, S.; Richardson, S.J. The Role of Transthyretin in Oligodendrocyte Development. Sci. Rep. 2020, 10, 4189. [Google Scholar] [CrossRef]
- Abati, E.; Citterio, G.; Bresolin, N.; Comi, G.P.; Corti, S. Glial cells involvement in spinal muscular atrophy: Could SMA be a neuroinflammatory disease? Neurobiol. Dis. 2020, 140, 104870. [Google Scholar] [CrossRef] [PubMed]
- Liz, M.A.; Coelho, T.; Bellotti, V.; Fernandez-Arias, M.I.; Mallaina, P.; Obici, L. A Narrative Review of the Role of Transthyretin in Health and Disease. Neurol. Ther. 2020, 9, 395–402. [Google Scholar] [CrossRef]
- Gião, T.; Saavedra, J.; Cotrina, E.; Quintana, J.; Llop, J.; Arsequell, G.; Cardoso, I. Undiscovered Roles for Transthyretin: From a Transporter Protein to a New Therapeutic Target for Alzheimer’s Disease. Int. J. Mol. Sci. 2020, 21, 2075. [Google Scholar] [CrossRef]
- Vidoni, C.; Follo, C.; Savino, M.; Melone, M.A.B.; Isidoro, C. The Role of Cathepsin D in the Pathogenesis of Human Neurodegenerative Disorders. Med. Res. Rev. 2016, 36, 845–870. [Google Scholar] [CrossRef]
- Bunk, J.; Huarcaya, S.P.; Drobny, A.; Dobert, J.P.; Walther, L.; Rose-John, S.; Arnold, P.; Zunke, F. Cathepsin D variants associated with neurodegenerative diseases show dysregulated functionality and modified α-synuclein degradation properties. Front. Cell Dev. Biol. 2021, 9, 581805. [Google Scholar] [CrossRef]
- Johnson, C.H.; Ivanisevic, J.; Siuzdak, G. Metabolomics: Beyond biomarkers and towards mechanisms. Nat. Rev. Mol. Cell Biol. 2016, 17, 451–459. [Google Scholar] [CrossRef]
- Moco, S. Studying Metabolism by NMR-Based Metabolomics. Front. Mol. Biosci. 2022, 9, 882487. [Google Scholar] [CrossRef]
- Moon, J.S.; Lee, J.E.; Yoon, J.S. Variation in Serum Creatinine Level Is Correlated to Risk of Type 2 Diabetes. Endocrinol. Metab. 2013, 28, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Freigang, M.; Wurster, C.D.; Hagenacker, T.; Stolte, B.; Weiler, M.; Kamm, C.; Schreiber-Katz, O.; Osmanovic, A.; Petri, S.; Kowski, A.; et al. Serum creatine kinase and creatinine in adult spinal muscular atrophy under nusinersen treatment. Ann. Clin. Transl. Neurol. 2021, 8, 1049–1063. [Google Scholar] [CrossRef] [PubMed]
- Alves, C.R.; Zhang, R.; Johnstone, A.J.; Garner, R.; Nwe, P.H.; Siranosian, J.J.; Swoboda, K.J. Serum creatinine is a biomarker of progressive denervation in spinal muscular atrophy. Neurology 2019, 94, e921–e931. [Google Scholar] [CrossRef] [PubMed]
- Faravelli, I.; Corti, S. Spinal muscular atrophy—Challenges in the therapeutic era. Nat. Rev. Neurol. 2020, 16, 655–656. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Author, Year | Patient Cohorts | Treatment | Samples | Methods | Major Findings | Molecular Pathways |
|---|---|---|---|---|---|---|
| Finkel et al., 2012 [28] | 108 children with SMA (type 1–3, age 2–12 years old) and 22 controls of similar age | No treatment | Plasma | Proteomics: multidimensional liquid chromatography and eight-plex iTRAQ labels with mass spectrometry. Metabolomics: QStar Elite quadrupole time-of-flight instrument. | Dipeptidyl Peptidase IV osteopontin, and tetranectin positively correlate with MHFMS; Fetuin-A and Vitronectin negatively correlate with MHFMS. | Dipeptidyl peptidase IV: glucose/insulin metabolism, immune response. Osteopontin: bone metabolism, immune response. Tetranectin: bone metabolism. Fetuin-A |
| Kessler et al., 2020 [29] | SMA type 3 (n = 9); SMA type 2 (n = 1) and age- and gender-matched controls (n = 10) | Baseline and after nusinersen treatment | CSF | Proteomics: Thermo Scientific QExactive HF-X mass spectrometer | Increased PARK7/DJ-1 in the CSF; two gene expression clusters in SMA patients based on age and two gene expression clusters in SMA patients based on treatment response. No significant single protein shift after therapy. | PARK7/DJ-1: DJ-1 acts as a chaperone and sensor under oxidative stress, protecting neurons by inhibiting α-synuclein aggregation. |
| Bianchi et al., 2021 [30] | SMA type 1 infants (n = 10) compared with age-matched controls (n = 10) | Baseline and after nusinersen treatment | CSF | Proteomics: SDS-PAGE followed by MALDI-TOF MS analysis | Increased levels of APOA1, APOE, and TTR after treatment. | APOA1: the primary apolipoprotein found in high-density lipoproteins (HDLs). APOE: protein linked to neurological and neurodegenerative diseases, with lipid transport, gene polymorphisms, and expression regulation being key factors. TTR: transport protein carrying thyroid hormone and vitamin A-bound retinol, with potential roles in neuroprotection and neurite outgrowth, mutations cause amyloidosis. |
| Schorling et al., 2022 [31] | SMA types 1, 2, and 3 (n = 3 mass spectrometry, validation n = 31) | Baseline and after nusinersen treatment | CSF | Mass spectroscopy | Cathepsin D decreased in SMA patients aged ≥2 months at the start of treatment but was only significant in patients who demonstrated a positive motor response. | Cathepsin D is a type of aspartyl protease prominently found in the central nervous system and skeletal and cardiac muscle. Its primary function is to catalyze the degradation of proteins within lysosomes. |
| Deutsch et al., 2021 [32] | SMA pediatric (type 1–3; n = 25) and matching controls (n = 25 serum, CSF) or n = 125 (urine]) | Baseline and after nusinersen treatment | Urine, serum, and CSF | Metabolomics: 600 MHz 1H-NMR spectrometer | No metabolome shift after therapy. Creatinine urinary level reduced in SMA patients. | |
| Errico et al., 2022 [33] | SMA pediatric patients (all three types, n = 27); no control group | Baseline and after nusinersen treatment | CSF | Metabolomics: 600 MHz 1H-NMR spectrometer | Effects after nusinersen treatment: increased metabolic rate of glucose (SMA type 1), increased ketone levels (SMA type 2), increased amino acid metabolism (SMA type 3). |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meneri, M.; Abati, E.; Gagliardi, D.; Faravelli, I.; Parente, V.; Ratti, A.; Verde, F.; Ticozzi, N.; Comi, G.P.; Ottoboni, L.; et al. Identification of Novel Biomarkers of Spinal Muscular Atrophy and Therapeutic Response by Proteomic and Metabolomic Profiling of Human Biological Fluid Samples. Biomedicines 2023, 11, 1254. https://doi.org/10.3390/biomedicines11051254
Meneri M, Abati E, Gagliardi D, Faravelli I, Parente V, Ratti A, Verde F, Ticozzi N, Comi GP, Ottoboni L, et al. Identification of Novel Biomarkers of Spinal Muscular Atrophy and Therapeutic Response by Proteomic and Metabolomic Profiling of Human Biological Fluid Samples. Biomedicines. 2023; 11(5):1254. https://doi.org/10.3390/biomedicines11051254
Chicago/Turabian StyleMeneri, Megi, Elena Abati, Delia Gagliardi, Irene Faravelli, Valeria Parente, Antonia Ratti, Federico Verde, Nicola Ticozzi, Giacomo P. Comi, Linda Ottoboni, and et al. 2023. "Identification of Novel Biomarkers of Spinal Muscular Atrophy and Therapeutic Response by Proteomic and Metabolomic Profiling of Human Biological Fluid Samples" Biomedicines 11, no. 5: 1254. https://doi.org/10.3390/biomedicines11051254
APA StyleMeneri, M., Abati, E., Gagliardi, D., Faravelli, I., Parente, V., Ratti, A., Verde, F., Ticozzi, N., Comi, G. P., Ottoboni, L., & Corti, S. (2023). Identification of Novel Biomarkers of Spinal Muscular Atrophy and Therapeutic Response by Proteomic and Metabolomic Profiling of Human Biological Fluid Samples. Biomedicines, 11(5), 1254. https://doi.org/10.3390/biomedicines11051254