
Assessment of Individual Radiosensitivity in Breast Cancer Patients Using a Combination of Biomolecular Markers




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Considerations
2.2. Chemicals
2.3. Patients and Cells
2.4. In Vitro Irradiation
2.5. In Vitro Irradiation
2.6. γH2AX/53BP1 DNA Repair Foci
2.7. Apoptosis
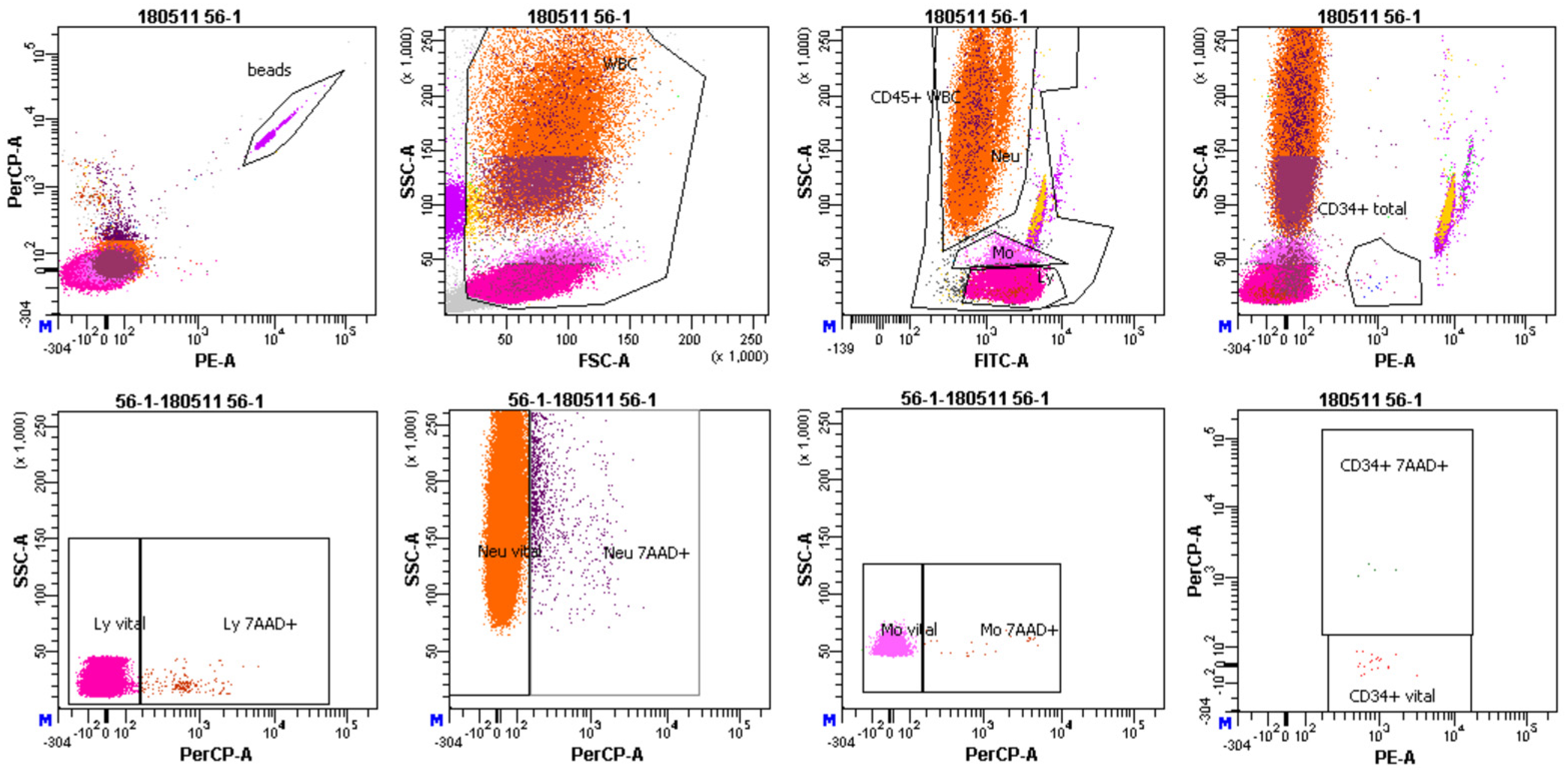
2.8. Chromosomal Aberrations and Cytokinesis-Blocked Micronuclei (CBMN) Assay
2.9. Statistical Analysis
3. Results
3.1. DNA Repair Foci in PBL of BC Patients
3.2. Cell Death in MNC Populations of BC Patients
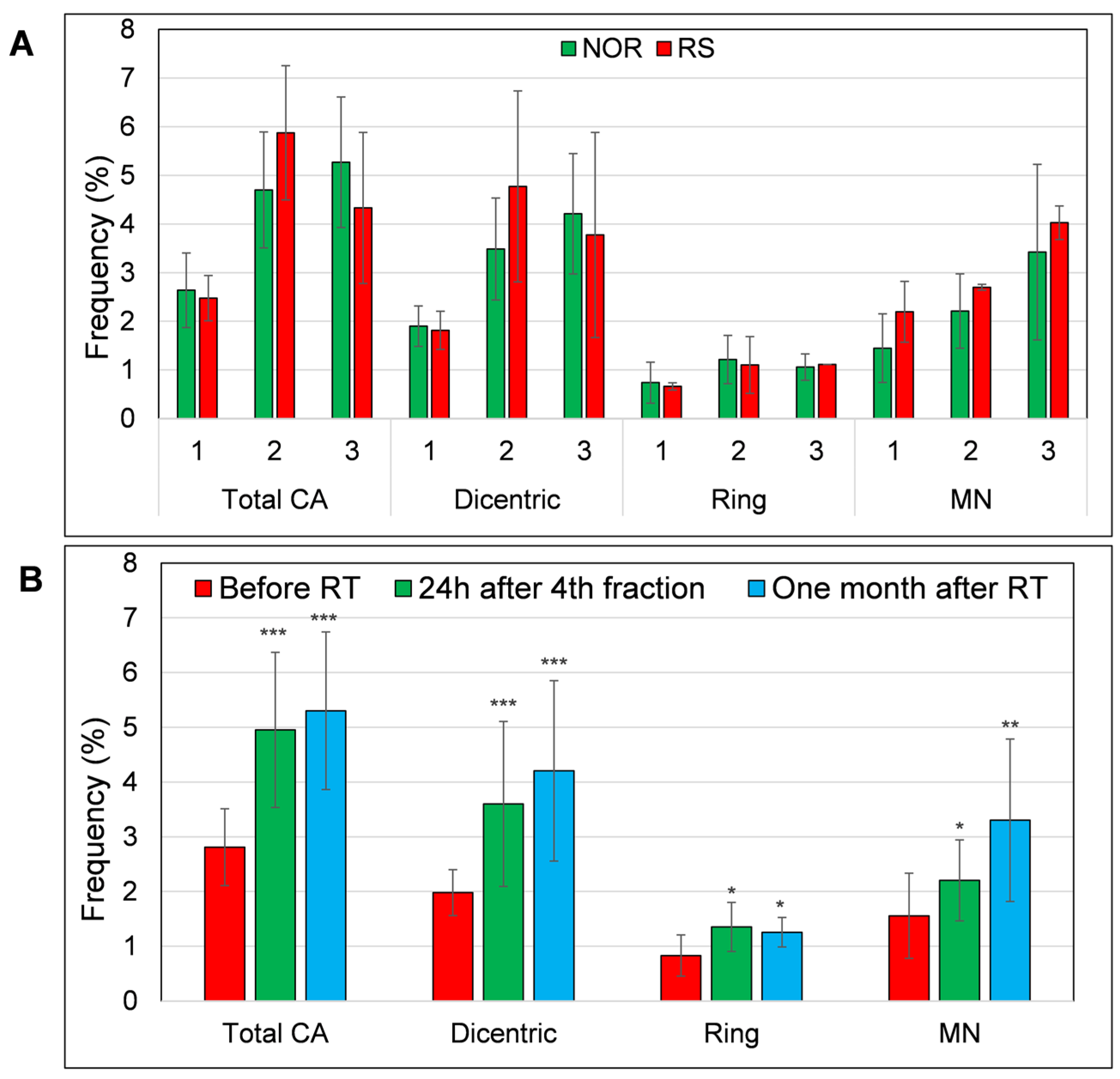
3.3. MN and Chromosomal Aberrations in PBL of BC Patients
3.4. DNA Repair Foci in PBL from BC Patients after In Vitro Irradiation
3.5. Apoptosis in PBL from BC Patients after In Vitro Ionizing Radiation Exposure
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Goodarzi, A.A.; Jeggo, P.; Lobrich, M. The influence of heterochromatin on DNA double strand break repair: Getting the strong, silent type to relax. DNA Repair 2010, 9, 1273–1282. [Google Scholar] [CrossRef] [PubMed]
- Markova, E.; Somsedikova, A.; Vasilyev, S.; Pobijakova, M.; Lackova, A.; Lukacko, P.; Belyaev, I. DNA repair foci and late apoptosis/necrosis in peripheral blood lymphocytes of breast cancer patients undergoing radiotherapy. Int. J. Radiat. Biol. 2015, 91, 934–945. [Google Scholar] [CrossRef] [PubMed]
- Djuzenova, C.S.; Elsner, I.; Katzer, A.; Worschech, E.; Distel, L.V.; Flentje, M.; Polat, B. Radiosensitivity in breast cancer assessed by the histone gamma-H2AX and 53BP1 foci. Radiat. Oncol. 2013, 8, 98. [Google Scholar] [CrossRef] [PubMed]
- Azria, D.; Riou, O.; Castan, F.; Nguyen, T.D.; Peignaux, K.; Lemanski, C.; Lagrange, J.L.; Kirova, Y.; Lartigau, E.; Belkacemi, Y.; et al. Radiation-induced CD8 T-lymphocyte Apoptosis as a Predictor of Breast Fibrosis After Radiotherapy: Results of the Prospective Multicenter French Trial. EBioMedicine 2015, 2, 1965–1973. [Google Scholar] [CrossRef]
- Aznar, M.C.; Duane, F.K.; Darby, S.C.; Wang, Z.; Taylor, C.W. Exposure of the lungs in breast cancer radiotherapy: A systematic review of lung doses published 2010–2015. Radiother. Oncol. J. Eur. Soc. Ther. Radiol. Oncol. 2018, 126, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Lobachevsky, P.; Leong, T.; Daly, P.; Smith, J.; Best, N.; Tomaszewski, J.; Thompson, E.R.; Li, N.; Campbell, I.G.; Martin, R.F.; et al. Compromized DNA repair as a basis for identification of cancer radiotherapy patients with extreme radiosensitivity. Cancer Lett. 2016, 383, 212–219. [Google Scholar] [CrossRef]
- Siva, S.; Lobachevsky, P.; MacManus, M.P.; Kron, T.; Moller, A.; Lobb, R.J.; Ventura, J.; Best, N.; Smith, J.; Ball, D.; et al. Radiotherapy for Non-Small Cell Lung Cancer Induces DNA Damage Response in Both Irradiated and Out-of-field Normal Tissues. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2016, 22, 4817–4826. [Google Scholar] [CrossRef]
- Lobachevsky, P.N.; Bucknell, N.W.; Mason, J.; Russo, D.; Yin, X.; Selbie, L.; Ball, D.L.; Kron, T.; Hofman, M.; Siva, S.; et al. Monitoring DNA Damage and Repair in Peripheral Blood Mononuclear Cells of Lung Cancer Radiotherapy Patients. Cancers 2020, 12, 2517. [Google Scholar] [CrossRef]
- Rogakou, E.P.; Boon, C.; Redon, C.; Bonner, W.M. Megabase chromatin domains involved in DNA double-strand breaks in vivo. J. Cell Biol. 1999, 146, 905–916. [Google Scholar] [CrossRef]
- Schultz, L.B.; Chehab, N.H.; Malikzay, A.; Halazonetis, T.D. p53 binding protein 1 (53BP1) is an early participant in the cellular response to DNA double-strand breaks. J. Cell Biol. 2000, 151, 1381–1390. [Google Scholar] [CrossRef]
- Belyaev, I.Y. Radiation-induced DNA repair foci: Spatio-temporal aspects of formation, application for assessment of radiosensitivity and biological dosimetry. Mutat. Res. 2010, 704, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Habash, M.; Bohorquez, L.C.; Kyriakou, E.; Kron, T.; Martin, O.A.; Blyth, B.J. Clinical and Functional Assays of Radiosensitivity and Radiation-Induced Second Cancer. Cancers 2017, 9, 147. [Google Scholar] [CrossRef] [PubMed]
- Borrego-Soto, G.; Ortiz-Lopez, R.; Rojas-Martinez, A. Ionizing radiation-induced DNA injury and damage detection in patients with breast cancer. Genet. Mol. Biol. 2015, 38, 420–432. [Google Scholar] [CrossRef]
- Horn, S.; Barnard, S.; Brady, D.; Prise, K.M.; Rothkamm, K. Combined analysis of gamma-H2AX/53BP1 foci and caspase activation in lymphocyte subsets detects recent and more remote radiation exposures. Radiat. Res. 2013, 180, 603–609. [Google Scholar] [CrossRef]
- Somaiah, N.; Chua, M.L.K.; Bourne, S.; Daley, F.; Hern, R.A.; Nuta, O.; Gothard, L.; Boyle, S.; Herskind, C.; Pearson, A.; et al. Correlation between DNA damage responses of skin to a test dose of radiation and late adverse effects of earlier breast radiotherapy. Radiother. Oncol. 2016, 119, 244–249. [Google Scholar] [CrossRef]
- Talbot, C.J.; Veldwijk, M.R.; Azria, D.; Batini, C.; Bierbaum, M.; Brengues, M.; Chang-Claude, J.; Johnson, K.; Keller, A.; Smith, S.; et al. Multi-centre technical evaluation of the radiation-induced lymphocyte apoptosis assay as a predictive test for radiotherapy toxicity. Clin. Transl. Radiat. Oncol. 2019, 18, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Scott, D. Chromosomal radiosensitivity, cancer predisposition and response to radiotherapy. Strahlenther. Onkol. 2000, 176, 229–234. [Google Scholar] [CrossRef]
- Imano, N.; Nishibuchi, I.; Kawabata, E.; Kinugasa, Y.; Shi, L.; Sakai, C.; Ishida, M.; Sakane, H.; Akita, T.; Ishida, T.; et al. Evaluating Individual Radiosensitivity for the Prediction of Acute Toxicities of Chemoradiotherapy in Esophageal Cancer Patients. Radiat. Res. 2021, 195, 244–252. [Google Scholar] [CrossRef]
- Chaouni, S.; Lecomte, D.D.; Stefan, D.; Leduc, A.; Barraux, V.; Leconte, A.; Grellard, J.M.; Habrand, J.L.; Guillamin, M.; Sichel, F.; et al. The Possibility of Using Genotoxicity, Oxidative Stress and Inflammation Blood Biomarkers to Predict the Occurrence of Late Cutaneous Side Effects after Radiotherapy. Antioxidants 2020, 9, 220. [Google Scholar] [CrossRef]
- Rzeszowska-Wolny, J.; Palyvoda, O.; Polanska, J.; Wygoda, A.; Hancock, R. Relationships between acute reactions to radiotherapy in head and neck cancer patients and parameters of radiation-induced DNA damage and repair in their lymphocytes. Int. J. Radiat. Biol. 2008, 84, 635–642. [Google Scholar] [CrossRef]
- Da Silva, E.B.; Cavalcanti, M.B.; Da Silva, C.S.F.; Fernandes, T.D.E.; Melo, J.A.; Lucena, L.; Netto, A.M.; Amaral, A. Micronucleus assay for predicting side effects of radiotherapy for cervical cancer. Biotech. Histochem. 2021, 96, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Cox, J.D.; Stetz, J.; Pajak, T.F. Toxicity criteria of the Radiation Therapy Oncology Group (RTOG) and the European Organization for Research and Treatment of Cancer (EORTC). Int. J. Radiat. Oncol. Biol. Phys. 1995, 31, 1341–1346. [Google Scholar] [CrossRef] [PubMed]
- Durdik, M.; Kosik, P.; Gursky, J.; Vokalova, L.; Markova, E.; Belyaev, I. Imaging flow cytometry as a sensitive tool to detect low-dose-induced DNA damage by analyzing 53BP1 and gammaH2AX foci in human lymphocytes. Cytometry. Part A J. Int. Soc. Anal. Cytol. 2015, 87, 1070–1078. [Google Scholar] [CrossRef] [PubMed]
- Durdik, M.; Kosik, P.; Kruzliakova, J.; Jakl, L.; Markova, E.; Belyaev, I. Hematopoietic stem/progenitor cells are less prone to undergo apoptosis than lymphocytes despite similar DNA damage response. Oncotarget 2017, 8, 48846–48853. [Google Scholar] [CrossRef]
- Moorhead, P.S.; Nowell, P.C.; Mellman, W.J.; Battips, D.M.; Hungerford, D.A. Chromosome Preparations of Leukocytes Cultured from Human Peripheral Blood. Exp. Cell Res. 1960, 20, 613–616. [Google Scholar] [CrossRef]
- Fenech, M.; Morley, A.A. Measurement of Micronuclei in Lymphocytes. Mutat. Res. 1985, 147, 29–36. [Google Scholar] [CrossRef]
- Droge, L.H.; Hennies, S.; Lorenzen, S.; Conradi, L.C.; Quack, H.; Liersch, T.; Helms, C.; Frank, M.A.; Schirmer, M.A.; Rave-Frank, M.; et al. Prognostic value of the micronucleus assay for clinical endpoints in neoadjuvant radiochemotherapy for rectal cancer. BMC Cancer 2021, 21, 219. [Google Scholar] [CrossRef]
- Moquet, J.; Higueras, M.; Donovan, E.; Boyle, S.; Barnard, S.; Bricknell, C.; Sun, M.; Gothard, L.; O’Brien, G.; Cruz-Garcia, L.; et al. Dicentric Dose Estimates for Patients Undergoing Radiotherapy in the RTGene Study to Assess Blood Dosimetric Models and the New Bayesian Method for Gradient Exposure. Radiat. Res. 2018, 190, 596–604. [Google Scholar] [CrossRef]
- Torudd, J.; Protopopova, M.; Sarimov, R.; Nygren, J.; Eriksson, S.; Markova, E.; Chovanec, M.; Selivanova, G.; Belyaev, I.Y. Dose-response for radiation-induced apoptosis, residual 53BP1 foci and DNA-loop relaxation in human lymphocytes. Int. J. Radiat. Biol. 2005, 81, 125–138. [Google Scholar] [CrossRef]
- Henriquez-Hernandez, L.A.; Carmona-Vigo, R.; Pinar, B.; Bordon, E.; Lloret, M.; Nunez, M.I.; Rodriguez-Gallego, C.; Lara, P.C. Combined low initial DNA damage and high radiation-induced apoptosis confers clinical resistance to long-term toxicity in breast cancer patients treated with high-dose radiotherapy. Radiat. Oncol. 2011, 6, 60. [Google Scholar] [CrossRef]
- Chua, M.L.; Somaiah, N.; A’Hern, R.; Davies, S.; Gothard, L.; Yarnold, J.; Rothkamm, K. Residual DNA and chromosomal damage in ex vivo irradiated blood lymphocytes correlated with late normal tissue response to breast radiotherapy. Radiother. Oncol. J. Eur. Soc. Ther. Radiol. Oncol. 2011, 99, 362–366. [Google Scholar] [CrossRef] [PubMed]
- Bourton, E.C.; Plowman, P.N.; Smith, D.; Arlett, C.F.; Parris, C.N. Prolonged expression of the gamma-H2AX DNA repair biomarker correlates with excess acute and chronic toxicity from radiotherapy treatment. Int. J. Cancer 2011, 129, 2928–2934. [Google Scholar] [CrossRef]
- Mumbrekar, K.D.; Fernandes, D.J.; Goutham, H.V.; Sharan, K.; Vadhiraja, B.M.; Satyamoorthy, K.; Bola Sadashiva, S.R. Influence of double-strand break repair on radiation therapy-induced acute skin reactions in breast cancer patients. Int. J. Radiat. Oncol. Biol. Phys. 2014, 88, 671–676. [Google Scholar] [CrossRef]
- Sterpone, S.; Cornetta, T.; Padua, L.; Mastellone, V.; Giammarino, D.; Testa, A.; Tirindelli, D.; Cozzi, R.; Donato, V. DNA repair capacity and acute radiotherapy adverse effects in Italian breast cancer patients. Mutat. Res. 2010, 684, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Brzozowska, K.; Pinkawa, M.; Eble, M.J.; Muller, W.U.; Wojcik, A.; Kriehuber, R.; Schmitz, S. In vivo versus in vitro individual radiosensitivity analysed in healthy donors and in prostate cancer patients with and without severe side effects after radiotherapy. Int. J. Radiat. Biol. 2012, 88, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Werbrouck, J.; De Ruyck, K.; Beels, L.; Vral, A.; Van Eijkeren, M.; De Neve, W.; Thierens, H. Prediction of late normal tissue complications in RT treated gynaecological cancer patients: Potential of the gamma-H2AX foci assay and association with chromosomal radiosensitivity. Oncol. Rep. 2010, 23, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Jakl, L.; Markova, E.; Kolarikova, L.; Belyaev, I. Biodosimetry of Low Dose Ionizing Radiation Using DNA Repair Foci in Human Lymphocytes. Genes 2020, 11, 58. [Google Scholar] [CrossRef]
- Markova, E.; Schultz, N.; Belyaev, I.Y. Kinetics and dose-response of residual 53BP1/gamma-H2AX foci: Co-localization, relationship with DSB repair and clonogenic survival. Int. J. Radiat. Biol. 2007, 83, 319–329. [Google Scholar] [CrossRef]
- Werbrouck, J.; Duprez, F.; De Neve, W.; Thierens, H. Lack of a correlation between gammaH2AX foci kinetics in lymphocytes and the severity of acute normal tissue reactions during IMRT treatment for head and neck cancer. Int. J. Radiat. Biol. 2011, 87, 46–56. [Google Scholar] [CrossRef]
- Olive, P.L.; Banath, J.P.; Keyes, M. Residual gammaH2AX after irradiation of human lymphocytes and monocytes in vitro and its relation to late effects after prostate brachytherapy. Radiother. Oncol. J. Eur. Soc. Ther. Radiol. Oncol. 2008, 86, 336–346. [Google Scholar] [CrossRef]
- Martin, N.T.; Nahas, S.A.; Tunuguntla, R.; Fike, F.; Gatti, R.A. Assessing ’radiosensitivity’ with kinetic profiles of gamma-H2AX, 53BP1 and BRCA1 foci. Radiother. Oncol. 2011, 101, 35–38. [Google Scholar] [CrossRef]
- Van Oorschot, B.; Hovingh, S.E.; Moerland, P.D.; Medema, J.P.; Stalpers, L.J.A.; Vrieling, H.; Franken, N.A.P. Reduced Activity of Double-Strand Break Repair Genes in Prostate Cancer Patients With Late Normal Tissue Radiation Toxicity. Int. J. Radiat. Oncol. 2014, 88, 664–670. [Google Scholar] [CrossRef]
- Foro, P.; Algara, M.; Lozano, J.; Rodriguez, N.; Sanz, X.; Torres, E.; Carles, J.; Reig, A.; Membrive, I.; Quera, J.; et al. Relationship Between Radiation-Induced Apoptosis of T Lymphocytes and Chronic Toxicity in Patients With Prostate Cancer Treated by Radiation Therapy: A Prospective Study. Int. J. Radiat. Oncol. 2014, 88, 1057–1063. [Google Scholar] [CrossRef]
- Schnarr, K.; Boreham, D.; Sathya, J.; Julian, J.; Dayes, I.S. Radiation-induced lymphocyte apoptosis to predict radiation therapy late toxicity in prostate cancer patients. Int. J. Radiat. Oncol. Biol. Phys. 2009, 74, 1424–1430. [Google Scholar] [CrossRef]
- Tang, Y.M.; Zhang, Y.Y.; Guo, L.; Peng, Y.; Luo, Q.L.; Xing, Y.G. Relationship between individual radiosensitivity and radiation encephalopathy of nasopharyngeal carcinoma after radiotherapy. Strahlenther. Onkol. 2008, 184, 510–514. [Google Scholar] [CrossRef]
- Vandevoorde, C.; Depuydt, J.; Veldeman, L.; De Neve, W.; Sebastia, N.; Wieme, G.; Baert, A.; De Langhe, S.; Philippe, J.; Thierens, H.; et al. In vitro cellular radiosensitivity in relationship to late normal tissue reactions in breast cancer patients: A multi-endpoint case-control study. Int. J. Radiat. Biol. 2016, 92, 823–836. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, S.; Brzozowska, K.; Pinkawa, M.; Eble, M.; Kriehuber, R. Chromosomal Radiosensitivity Analyzed by FISH in Lymphocytes of Prostate Cancer Patients and Healthy Donors. Radiat. Res. 2013, 180, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Vinnikov, V.; Hande, M.P.; Wilkins, R.; Wojcik, A.; Zubizarreta, E.; Belyakov, O. Prediction of the Acute or Late Radiation Toxicity Effects in Radiotherapy Patients Using Ex Vivo Induced Biodosimetric Markers: A Review. J. Pers. Med. 2020, 10, 285. [Google Scholar] [CrossRef] [PubMed]
- Rozgaj, R.; Kasuba, V.; Sentija, K.; Prlic, I. Radiation-induced chromosomal aberrations and haematological alterations in hospital workers. Occup. Med. 1999, 49, 353–360. [Google Scholar] [CrossRef]
- Gamulin, M.; Kopjar, N.; Grgic, M.; Ramic, S.; Bisof, V.; Garaj-Vrhovac, V. Genome damage in oropharyngeal cancer patients treated by radiotherapy. Croat. Med. J. 2008, 49, 515–527. [Google Scholar] [CrossRef]
- Gamulin, M.; Kopjar, N.; Grgic, M.; Ramic, S.; Viculin, T.; Petkovic, M.; Garaj-Vrhovac, V. Cytogenetic follow-up in testicular seminoma patients exposed to adjuvant radiotherapy. Coll. Antropol. 2010, 34, 455–465. [Google Scholar] [PubMed]
- Melnikov, A.A.; Vasilyev, S.A.; Musabaeva, L.I.; Velikaya, V.V.; Gribova, O.V.; Startseva Zh, A.; Urazova, L.N.; Lebedev, I.N.; Choynzonov, E.L. Cytogenetic effects of neutron therapy in patients with parotid gland tumors and relapse of breast cancer. Exp. Oncol. 2012, 34, 354–357. [Google Scholar] [PubMed]
- Hartel, C.; Nasonova, E.; Fuss, M.C.; Nikoghosyan, A.V.; Debus, J.; Ritter, S. Persistence of radiation-induced aberrations in patients after radiotherapy with C-ions and IMRT. Clin. Transl. Radiat. Oncol. 2018, 13, 57–63. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Durdik, M.; Markova, E.; Kosik, P.; Vigasova, K.; Gulati, S.; Jakl, L.; Vrobelova, K.; Fekete, M.; Zavacka, I.; Pobijakova, M.; et al. Assessment of Individual Radiosensitivity in Breast Cancer Patients Using a Combination of Biomolecular Markers. Biomedicines 2023, 11, 1122. https://doi.org/10.3390/biomedicines11041122
Durdik M, Markova E, Kosik P, Vigasova K, Gulati S, Jakl L, Vrobelova K, Fekete M, Zavacka I, Pobijakova M, et al. Assessment of Individual Radiosensitivity in Breast Cancer Patients Using a Combination of Biomolecular Markers. Biomedicines. 2023; 11(4):1122. https://doi.org/10.3390/biomedicines11041122
Chicago/Turabian StyleDurdik, Matus, Eva Markova, Pavol Kosik, Katarina Vigasova, Sachin Gulati, Lukas Jakl, Katarina Vrobelova, Marta Fekete, Ingrid Zavacka, Margita Pobijakova, and et al. 2023. "Assessment of Individual Radiosensitivity in Breast Cancer Patients Using a Combination of Biomolecular Markers" Biomedicines 11, no. 4: 1122. https://doi.org/10.3390/biomedicines11041122
APA StyleDurdik, M., Markova, E., Kosik, P., Vigasova, K., Gulati, S., Jakl, L., Vrobelova, K., Fekete, M., Zavacka, I., Pobijakova, M., Dolinska, Z., & Belyaev, I. (2023). Assessment of Individual Radiosensitivity in Breast Cancer Patients Using a Combination of Biomolecular Markers. Biomedicines, 11(4), 1122. https://doi.org/10.3390/biomedicines11041122