
Natural IgG Anti-F (ab’)2 Autoantibody Activity in Children with Autism

Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Cognitive and Behavioral Assessments
2.3. Blood Drawing Procedures and Titration of NAAbs
2.4. Statistical Analysis
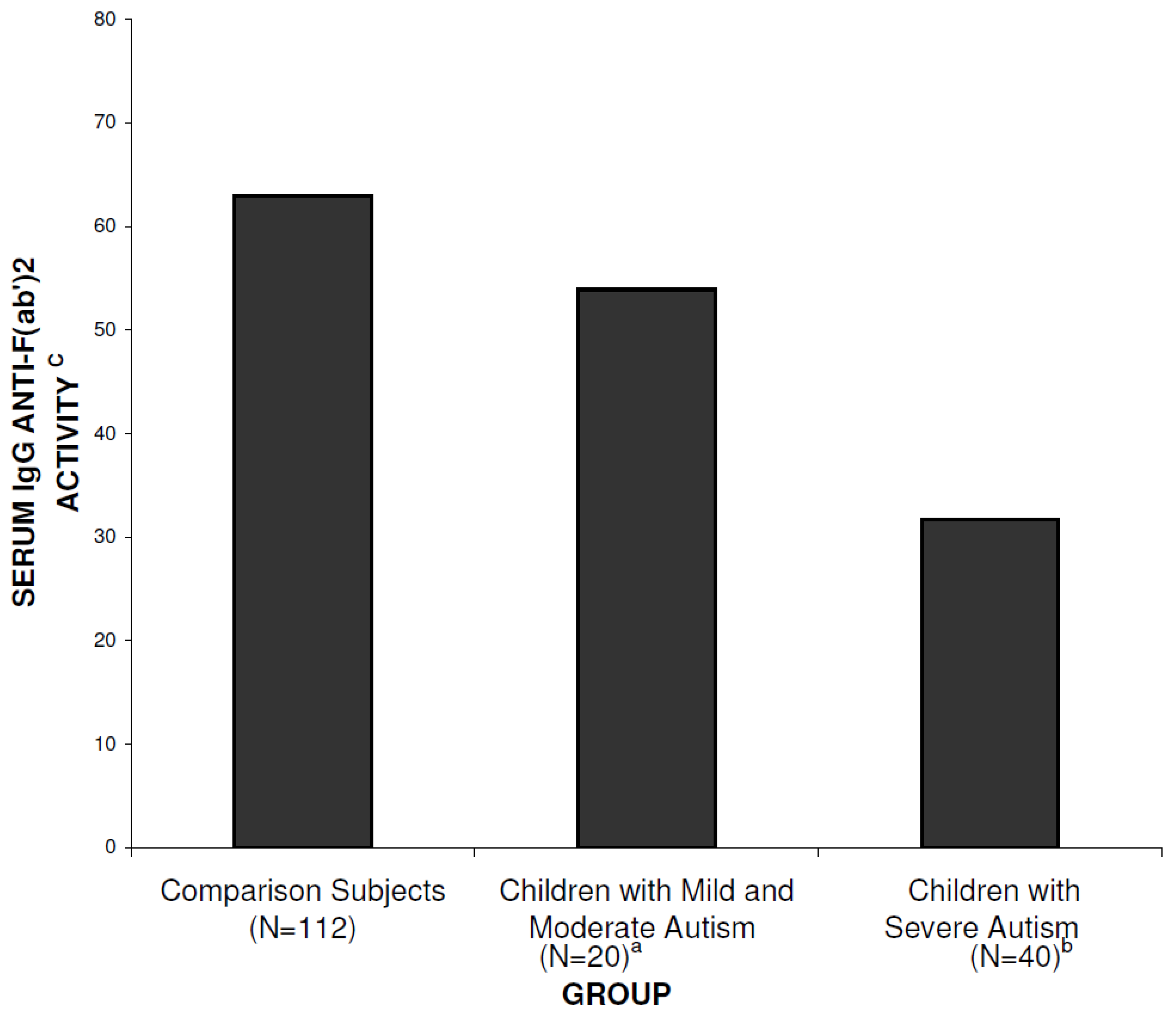
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Saresella, M.; Marventano, I.; Guerini, F.R.; Mancuso, R.; Ceresa, L.; Zanzottera, M.; Rusconi, B.; Maggioni, E.; Tinelli, C.; Clerici, M. An autistic endophenotype results in complex immune dysfunction in healthy siblings of autistic children. Biol. Psychiatry 2009, 66, 978–984. [Google Scholar] [CrossRef]
- Morgan, J.T.; Chana, G.; Pardo, C.A.; Achim, C.; Semendeferi, K.; Buckwalter, J.; Courchesne, E.; Everall, I.P. Microglial activation and increased microglial density observed in the dorsolateral prefrontal cortex in autism. Biol. Psychiatry 2010, 68, 368–376. [Google Scholar] [CrossRef]
- Mead, J.; Aschwood, P. Evidence supporting an altered immune response in ASD. Immun. Lett. 2015, 163, 49–55. [Google Scholar] [CrossRef]
- Sweeten, T.L.; Posey, D.J.; McDougle, C.J. High blood monocyte counts and neopterin levels in children with autistic disorder. Am. J. Psychiatry 2003, 160, 1691–1693. [Google Scholar] [CrossRef]
- Enstrom, A.M.; Onore, C.E.; Van de Water, J.A.; Ashwood, P. Differential monocyte responses to TLR ligands in children with autism spectrum disorders. Brain Behav. Immun. 2010, 24, 64–71. [Google Scholar] [CrossRef]
- Ashwood, P.; Corbett, B.A.; Kantor, A.; Schulman, H.; Van de Water, J.; Amaral, D.G. In search of cellular immunophenotypes in the blood of children with autism. PLoS ONE 2011, 6, e19299. [Google Scholar] [CrossRef]
- Cryan, J.F.; Dinan, T.G. Mind-altering microorganisms: The impact of the gut microbiota on brain and behaviour. Nat. Rev. Neurosci. 2012, 13, 701–712. [Google Scholar] [CrossRef]
- Olszak, T.; An, D.; Zeissig, S.; Vera, M.P.; Richter, J.; Franke, A.; Glickman, J.N.; Siebert, R.; Baron, R.M.; Kasper, D.L.; et al. Microbial exposure during early life has persistent effects on natural killer T cell function. Science 2012, 336, 489–493. [Google Scholar] [CrossRef]
- Kwekkeboom, J. Modulation of dendritic cells and regulatory T cells by naturally occurring antibodies. Abv. Exp. Biol. 2012, 750, 133–144. [Google Scholar] [CrossRef]
- Ashwood, P.; Krakowiak, P.; Hertz-Picciotto, I.; Hansen, R.; Pessah, I.N.; Van de Water, J. Altered T cell responses in children with autism. Brain Behav. Immun. 2011, 25, 840–849. [Google Scholar] [CrossRef]
- Hsiao, E.Y.; McBride, S.W.; Hsien, S.; Sharon, G.; Hyde, E.R.; McCue, T.; Codelli, J.A.; Chow, J.; Reisman, S.E.; Petrosino, J.F.; et al. The microbiota modulates gut physiology and behavioral abnormalities associated with autism. Cell 2013, 155, 1451–1463. [Google Scholar] [CrossRef]
- Ashwood, P.; Krakowiak, P.; Hertz-Picciotto, I.; Hansen, R.; Pessah, I.; Van de Water, J. Elevated plasma cytokines in autism spectrum disorders provide evidence of immune dysfunction and are associated with impared bahavioral outcome. Brain Behav. Immun. 2011, 25, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Frye, R.E.; Sequeira, J.M.; Quadros, E.V.; James, S.J.; Rossignol, D.A. Cerebral folate receptor autoantibodies in autism spectrum disorder. Mol. Psychiatry 2013, 18, 369–381. [Google Scholar] [CrossRef] [PubMed]
- Mostafa, G.A.; Al-Ayadhi, L.Y. Increased serum levels of anti-ganglioside M1 auto-antibodies in autistic children: Relation to the disease severity. J. Neuroinflamm. 2011, 8, 39. [Google Scholar] [CrossRef]
- Moeller, S.; Lau, N.M.; Green, P.H.; Hellberg, D.; Higgins, J.J.; Rajadhyaksha, A.M.; Alaedini, A. Lack of association between autism and anti-GM1 ganglioside antibody. Neurology 2013, 81, 1640–1641. [Google Scholar] [CrossRef]
- Yuwiler, A.; Shih, J.C.; Chen, C.-H.; Ritvo, E.R.; Hanna, G.; Ellison, G.W.; King, B.H. Hyperserotoninemia and antiserotonin antibodies in autism and other disorders. J. Autism Dev. Disord. 1992, 22, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Todd, R.D.; Ciaranello, R.D. Demonstration of inter- and intraspecies differences in serotonin binding sites by antibodies from an autistic child. Neurobiology 1985, 82, 612–616. [Google Scholar] [CrossRef] [PubMed]
- Stein, M.; Miller, A.H.; Trestman, R.L. Depression, the immune system, and health and illness. Arch. Gen. Psychiatry 1991, 48, 171–177. [Google Scholar] [CrossRef]
- Croonenberghs, J.; Wauters, A.; Devreese, K.; Verkerk, R.; Scharpe, S.; Bosmans, E.; Egyed, B.; Deboutte, D.; Maes, M. Increased serum albumin, γ globulin, immunoglobulin IgG, and IgG2 and IgG4 in autism. Psychol. Med. 2002, 32, 1457–1463. [Google Scholar] [CrossRef]
- Heuer, L.; Ashwood, P.; Schauer, J.; Goines, P.; Krakowiak, P.; Hertz-Picciotto, I.; Hansen, R.; Croen, L.A.; Pessah, I.N.; Van de Water, J. Reduced levels of immunoglobulin in children with autism correlates with behavorial symptoms. Autism Res. 2008, 1, 275–283. [Google Scholar] [CrossRef]
- Perry, V.; O’Connor, V. C1q the perfect complement for a synaptic feast. Nat. Rev Neurosci. 2008, 9, 807–811. [Google Scholar] [CrossRef]
- Voineagu, I.; Wang, X.; Johnston, P.; Lowe, J.K.; Tian, Y.; Horvath, S.; Mill, J.; Cantor, R.M.; Blencowe, B.J.; Geschwind, D.H. Transcriptomic analysis of autistic brain reveals convergent molecular pathology. Nature 2011, 474, 380–384. [Google Scholar] [CrossRef] [PubMed]
- Swedo, S.E.; Leonard, H.L.; Garvey, M.; Mittleman, M.B.B.; Allen, A.J.; Perlmutter, S.; Dow, S.; Zamkoff, J.; Dubbert, B.K.; Lougee, L.; et al. Pediatric Autoimmune neuropsychiatric disorders associated with streptococcal infections: Clinical description of the first 50 cases. Am. J. Psychiatry 1998, 155, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Hollander, E.; DelGiudice-Asch, G.; Simon, L.; Schmeidler, J.; Cartwright, C.; DeCaria, C.M.; Kwon, J.; Cunningham-Rundles, C.; Chapman, F.; Zabriskie, J.B. B Lymphocyte antigen D8/17 and repetitive behaviours in autism. Am. J. Psychiatry 1999, 156, 317–320. [Google Scholar] [CrossRef] [PubMed]
- Perlmutter, S.J.; Leitman, S.F.; Garvey, M.A.; Hamburger, S.; Feldman, E.; Leonard, H.L.; Swedo, S.E. Therapeutic plasma exchange and intravenous immunoglobulin for obsessive-compulsive disorder and tic disorders in childhood. Lancet 1999, 354, 1153–1158. [Google Scholar] [CrossRef]
- Marcerollo, A.; Martino, D. Pediatric autoimmune neuropsychiatric disorders associated with streptococcal infections (PANDAS): An evolving concept. Tremor Other Hyperkinetic Mov. 2013, 3. [Google Scholar]
- Cohn, M. Autoimmunity: Rationalizing possible pathways from initiation to disease. J. Theor. Biol. 2015, 375, 40–51. [Google Scholar] [CrossRef]
- Avrameas, S.; Selmi, C. Natural autoantibodies in the physiology and pathophysiology of the immune system. J. Autoimmun. 2013, 41, 46–49. [Google Scholar] [CrossRef]
- Gold, M.; Pul, R.; Bach, J.-P.; Stangel, M.; Dodel, R. Pathogenic and physiological autoantibodies in the central nervous system. Immunol. Rev. 2012, 248, 68–86. [Google Scholar] [CrossRef]
- Lord, C. Diagnostic instruments in autism spectrum disorders. In Handbook of Autism and Pervasive Developmental Disorders, 2nd ed.; Cohen, D.J., Volkmar, F.R., Eds.; John Wiley and Sons Inc.: New York, NY, USA, 1997. [Google Scholar]
- Risi, S.; Lord, C.; Gotham, K.; Corsello, C.; Chrysler, C.; Szatmari, P.; Cook, E.H.; Leventhal, B.L.; Pickles, A. Combining information from multiple sources in the diagnosis of autism spectrum disorders. J. Am. Acad. Child Adolesc. Psychiatry 2006, 45, 1094–1103. [Google Scholar] [CrossRef]
- Robert, C.; Pasquier, L.; Cohen, D.; Fradin, M.; Canitano, R.; Damaj, L.; Odent, S.; Tordjman, S. Role of genetics in the etiology of autistic spectrum disorder: Towards a hierarchical diagnostic strategy. Int. J. Mol. Sci. 2017, 18, 618. [Google Scholar] [CrossRef] [PubMed]
- Tordjman, S.; Gutknecht, L.; Carlier, M.; Spitz, E.; Antoine, C.; Slama, F.; Carsalade, V.; Cohen, D.J.; Ferrari, P.; Roubertoux, P.L.; et al. Role of the serotonin transporter gene in the behavioral expression of autism. Mol. Psychiatry 2001, 6, 434–439. [Google Scholar] [CrossRef] [PubMed]
- Torrente, F.; Ashwood, P.; Day, R.; Machado, N.; Furlano, R.I.; Anthony, A.; Davies, S.E.; Wakefield, A.J.; Thomson, M.A.; Walker-Smith, J.A.; et al. Small intestinal enteropathy with epithelial IgG and complement deposition in children with regressive autism. Mol. Psychiatry 2002, 7, 375–382. [Google Scholar] [CrossRef] [PubMed]
- Sweeten, T.L.; Bowyer, S.L.; Posey, D.J.; Halberstadt, G.M.; McDougle, C.J. Increased prevalence of familial autoimmunity in probands with pervasive developmental disorders. Pediatrics 2003, 112, e420. [Google Scholar] [CrossRef] [PubMed]
- Molenaar, P.C.; Campbell, C.G. The New Person-Specific Paradigm in Psychology. Curr. Dir. Psychol. Sci. 2009, 18, 112–117. [Google Scholar] [CrossRef]
- Anspach, M.R.; Varela, F. Introduction aux sciences cognitives. In Le Système Immunitaire: Un “soi” Cognitif Autonome; Andler, D., Ed.; Galimard: Paris, France, 2004; Chapter 19; pp. 585–605. [Google Scholar]
- Varela, F.J.; Coutinho, A.; Stewart, J. What is the immune network for? In SFI Studies in the Sciences of Complexity; Stein, W., Varela, F., Eds.; CRC Press: Boca Raton, FL, USA, 1993; Volume III. [Google Scholar]
- Coutinho, A.; Hori, S.; Carvalho, T.; Caramalho, I.; Demengeot, J. Regulatory T cells: The physiology of autoreactivity in dominant tolerance and “quality control” of immune responses. Immunol. Rev. 2011, 182, 89–98. [Google Scholar] [CrossRef]
- Avrameas, S.C. Autopolyreactivity Confers a Holistic Role in the immune system. Scand. J. Immunol. 2016, 83, 227–234. [Google Scholar] [CrossRef]
- Cheng, M.H.; Anderson, M.S. Monogenic autoimmunity. Annu. Rev. Immunol. 2012, 30, 393–427. [Google Scholar] [CrossRef] [PubMed]
- French, S.S.; Moore, M.C.; Demas, G.E. Ecological immunology: The organism in context. Integr. Comp. Biol. 2009, 49, 246–253. [Google Scholar] [CrossRef]


{kind=link}
| Studies | Measure | Individuals with Autism (n) | Controls (n) | Results |
|---|---|---|---|---|
| Warren et al., 1987 * | Investigation of the natural cytotoxic potential of peripheral blood mononuclear cells using K562 tumor cells as target cells | Children, adolescents, and adults with autism (age: 3 to 28) (n = 31) | Age-matched healthy volunteers (n = 15) Healthy adults (n = 23) | Reduced levels of cytotoxicity not correlated with quantitative alteration in NK cells. |
| Gupta et al., 1998 * | Th1-like (IL-2, IFN-γ) and Th2-like (IL-4, IL-6, and IL-10) cytokines examined in CD4+ and CD8+ T cells. Intracellular cytokines measured using specific antibodies to various cytokines and anti-CD4 or anti-CD8 monoclonal antibodies by FACScan | Children with autism (n = 20) | Typically developing (TD) children (n = 20) | Lower proportion of IFN-γ + CD4+ T cells, IL-2+CD4+ T cells (Th1), IFN-γ+CD8+ and IL-2+CD8+ T cells (TC1) in children with autism compared to TD controls. Increased IL-4+CD4+ T cells (Th2) and IL-4+CD8+ T cells (TC2) in autism. The proportion of IL-6+ CD4+, IL-6+CD8+, IL-10+CD4+, IL-10+CD8+ T cells was comparable in autism and control groups. |
| Sweeten et al., 2003 [4] | Leukocyte counts and plasma neopterin levels | Children with autism (n = 31) | TD children (n = 28) | Higher monocyte count and neopterin level in children with autism compared to TD children. |
| Saresalla et al., 2009 [1] | Immunophenotypic analysis; immunofluorescent staining and virological analysis | Children with autism (n = 20) | Non-autistic siblings (NAS) (n = 15). TD children (n = 20) | Decreased CD8+ T lymphocytes and diminution of CD8+ effector memory as well as CD4+ terminally differentiated lymphocytes in children with autism and NAS compared to TD children. |
| Enstrom et al., 2009 * | Gene expression screen and cellular functional analysis on peripheral blood | Children with autism spectrum disorder (ASD) (n = 52) | TD children (n = 27) | Upregulated RNA expression of NK cell receptors and effector molecules in ASD. Flow cytometric analysis of NK cells demonstrated increased production of perforin, granzyme B, and interferon gamma (IFNγ) under resting conditions in ASD children. Reduced cytotoxicity of NK cells and lower presence of perforin, granzyme B, IFNγ in NK cells in ASD children compared to TD controls. |
| Morgan et al., 2010 [2] | Microglial cell density stereologically estimated via optical fractionator and average somal volume quantified via isotropic nucleator in the dorsolateral prefrontal cortex | Children, adolescents, and adults with autism (age: 3 to 41) (n = 13) | TD children, adolescents, and adults (age: 1 to 44) (n = 9) | Microglia appeared activated in 9 out of 13 individuals with autism. Morphological alterations included somal enlargement, process retraction and thickening, and extension of filopodia from processes. Average microglial somal volume was significantly increased in white matter, with a trend in gray matter. Microglial cell density was increased in gray matter. |
| Ashwood et al., 2011 [6] | Number and phenotype of circulating blood cells (644 primary and secondary variables, including cell counts and the abundance of cell surface antigens) assessed using microvolume laser scanning cytometry | Children with autism (n = 70), including low functioning children (n = 35) and high functioning children (n = 35) | TD children (n = 35) | The absolute number of B cells per volume of blood was over 20% higher for children with autism, and the absolute number of NK cells was about 40% higher. The absolute number of T cells was not different across groups, but the number of cellular activation markers, including HLA-DR and CD26 on T cells and CD38 on B cells, was significantly higher in the autism group compared to the control group. |
| Ashwood et al., 2010 [10] | Cellular responses following in vitro stimulation of peripheral blood mononuclear cells with PHA and tetanus | Children with ASD (n = 66) | TD children (n = 73) | Increased production of GM-CSF, TNFα, IL-13 and decreased production of IL-12p40 following PHA stimulation in the ASD group compared to the TD control group. Increased pro-inflammatory or TH1 cytokines associated with greater impairments in ASD core features and aberrant behaviors; production of GM-CSF and TH2 cytokines was associated with better cognitive and adaptive function. Reduced frequency in ASD of CD3+, CD4+ and CD8+ T cells expressing activation markers CD134 and CD25 (but not CD69, HLA-DR or CD137) following stimulation. |
| Torres et al., 2012 * | Killer-cell immunoglobulin-like receptor (KIR) proteins expressed on NK cells using the whole genome amplification and HLA genotyping | Children with autism (n = 158) | TD children (n = 176) | Increased expression in four activating KIR genes (2DS5, 3DS1, 2DS1 and 2DS4) and the cognate HLA-C2 ligand of the activating KIR gene 2DS1. |
| Studies | Measure | Individuals with Autism or Animal Models (n) | Controls (n) | Results |
|---|---|---|---|---|
| Sandler et al., 1999 a | An intervention trial using a minimally absorbed oral antibiotic. Short-term improvement was noted using multiple pre- and post-therapy evaluations: Behavior and communication analog rating scales Quantitative fecal flora stool specimen data from children with autism prior to vancomycin therapy | Children with regressive-onset autism (n = 11) Children with autism (n = 4) | Healthy adults (n = 104) | An open-label vancomycin trial indicates the possibility of a gut flora-brain connection in a subset of children with autism and diarrhea. Improvement for children with autism in communication and behavior following the vancomycin trial. Anaerobic cocci were absent from the stools of each of the 4 children with autism tested whereas they were present in 93% of the adults’ specimens. |
| Finegold et al., 2002 a | Comparison of faecal flora; clostridial counts | Children with regressive autism (n = 13) | Typically developing (TD) children (n = 8) | Clostridial counts in fecal flora were higher in children with regressive autism; children with autism had 9 species of Clostridium not found in TD children, whereas controls yielded only 3 species not found in children with autism. |
| Parracho et al., 2005 b | Fecal bacterial populations were assessed using a culture-independent technique, fluorescence in situ hybridization, with oligonucleotide probes targeting predominant components of the gut flora | Children with autism spectrum disorder (ASD) (n = 58) | Healthy siblings (n = 12) TD children (n = 10) | The fecal flora of ASD patients contained a higher incidence of the Clostridium histolyticum group of bacteria compared to healthy children. The non-autistic sibling group had an intermediate level of the Clostridium histolyticum group, which was not significantly different from either of the other subject groups. |
| Finegold et al., 2010 b | Fecal microbial flora using the bacterial tag encoded FLX amplicon pyrosequencing (bTEFAP) procedure | Children with autism and gastrointestinal symptoms (n = 33) | Healthy siblings (n = 7) TD children (n = 8) | High levels of Bacteroidetes and Desulfovibrio species were found in the severely autistic group. |
| MacFabe et al., 2010 a | Effects of propionic acid (PPA) on brain tissue and behavior (social, restricted interest and reversal in the T-maze task) | Rats in propionic acid (PPA) (n = 20) | Rats in phosphate-buffered saline (PBS) (n = 17) | PPA rats showed reactive astrogliosis and activated microglia (indicating an innate neuroinflammatory response) and impairment behavior (restricted behavioral interest to a specific object among a group of objects, impaired social behavior, and impaired reversal in a T-maze task) compared to PBS controls. |
| Adams et al., 2011 b | Assessment of gastrointestinal flora, gastrointestinal status and yeast using standard culture growth-based techniques and other markers of digestive function | Children with ASD (n = 58) | TD children (n = 39) | Abnormally low levels of lysozyme and total short chain fatty acids in the ASD group, including acetate, proprionate, and valerate. The ASD group had lower levels of species of Bifidobacter and higher levels of species of Lactobacillus, but similar levels of other bacteria and yeast compared to the control group. |
| Williams et al., 2011 b | Expression of human genes involved in carbohydrate digestion and transport along with bacterial community composition in intestinal biopsies. Composition of intestinal bacteria was examined using metagenomic analysis | Children with autistic disorder (AD) and gastrointestinal disease (n = 15) | TD children with gastrointestinal disease (n = 7) | Deficient ileal transcripts encoding disaccharidases and hexose transporters in the AD group, and impaired primary pathway for carbohydrate digestion and transport in enterocytes associated with expression of the intestinal transcription factor, CDX2. Composition of dysbiosis was decreased in Bacteroidetes but increased in the ratio of Firmicutes to Bacteroidetes and in Betaproteobacteria. Expression levels of disaccharidases and transporters associated with the abundance of affected bacterial phylotypes. |
| Williams et al., 2012 b | Novel PCR-based methods for detection, quantitation, and phylogenetic characterization of Sutterella species in intestinal biopsy samples | Children with autism and gastrointestinal dysfunction (n = 23) | TD children with gastrointestinal dysfunction (n = 9) | Sutterella 16S rRNA gene sequences were found in 12 of 23 children with autism but in none of the controls. High levels of intestinal and mucoepithelial-associated Sutterella species were associated with gastrointestinal disturbances in children with autism. |
| Wang et al., 2012 a | Concentrations of short chain fatty acids, phenols and ammonia measured in fecal samples | Children with ASD (n = 23) | TD children (n = 31) | Levels of fecal total short chain fatty acid, fecal ammonia, fecal acetic, butyric, isobutyric, valeric and isovaleric acids were higher in the ASD group compared to the control group. Similar levels of fecal phenol and pH between groups. |
| Thomas et al., 2012 a | Effects of treatments (intracerebroventricular infusion procedure) on behavior and phospholipid components: automated activity monitors, behavioral testing, lipid extraction, electrospray ionization mass spectrometry analysis | Long-Evans rats with propionic acid (PPA) treatment (n = 12) | Long-Evans rats with phosphate-buffered saline (PBS) (n = 12) | PPA-rats displayed abnormal ASD-like behaviors associated with alteration in brain and blood phospholipid molecular species. |
| Gondalia et al., 2012 b | Parental questionnaires on child diagnosis and symptoms, including gastrointestinal symptoms. Collection of stool samples for analyses: DNA extraction, bacterial identification (numbers of species, phylum levels, etc.) |
Children with autism but without gastrointestinal dysfunction (n = 23) Children with autism and gastrointestinal dysfunction (n = 28) | TD siblings (n = 53) | Firmicutes (70%), Bacteroidetes (20%) and Proteobacteria (4%) were the most dominant phyla in samples. |
| Kang et al., 2013 b | Pyrosequencing the V2/V3 regions in bacterial 16S rDNA from fecal DNA samples | Children with autism and gastrointestinal disorders (n = 20) | TD children without gastrointestinal disorders (n = 20) | The presence of autistic symptoms, rather than the severity of gastrointestinal symptoms, was associated with less diverse gut microbiomes. There were significantly lower abundances of Prevotella, Coprococcus and unclassified Veillonellaceae in children with autism. |
| Hsiao et al., 2013 [11] | Microbial composition Gut permeability ASD-related defects in communicative, stereotypic, anxiety-like and sensorimotor behaviors | Offspring from MIA (maternal immune activation) pregnant C57BL/6N mice treated with human commensal Bacteroides fragilis | Offspring from MIA pregnant C57BL/6N mice treated with vehicle in 1.5% sodium bicarbonate | Oral treatment of MIA offspring with Bacteroides fragilis corrects gut permeability, alters microbial composition and improves ASD-related defects in communication, stereotyped, anxiety-like and sensorimotor behaviors. |
| Studies | Measure | Individuals with Autism (n) | Controls (n) | Results |
|---|---|---|---|---|
| Croonenberghs et al., 2002 [19] | Total serum protein (TSP) and serum concentrations of albumin, alpha1 globulin, alpha2 globulin, beta globulin and gamma globulins, IgA, IgM and IgG and the IgG subclasses IgG1, IgG2, IgG3 and IgG4 | Adolescents with autism (age: 13 to 19) (n = 18) | Typically developing (TD) adolescents (n = 22) | Increased concentrations of TSP in autism related to increased serum concentrations of albumin; significant correlations between gamma globulin and social problems, especially social withdrawal. Increased serum concentrations of IgG, IgG2 and IgG4 and albumin were significantly correlated with social problems and social withdrawal, respectively. |
| Croonenberghs et al., 2002 * | Production of interleukin (IL)-6, IL-10, the IL-1 receptor antagonist (IL-1 RA), interferon (IFN)-γ and tumor necrosis factor (TNF)-α by whole blood and serum concentrations of IL-6, IL-2 receptor (IL-2R) and IL-1RA | Adolescents with autism (age: 12 to 18) (n = 13) | TD adolescents (n = 13) | Increased production of IFN-γ, IL-1RA, IL-6 and TNF-α in adolescents with autism. Similar serum concentrations of IL-6, IL-2R and IL-1RA between adolescents with autism and TD adolescents. |
| Cabanlit et al., 2007 * | IgG antibodies against protein extracts from specific regions of the human adult brain, including the hypothalamus and thalamus | Children with autism spectrum disorder (ASD) (n = 63) | TD children (n = 63) Healthy siblings (n = 25) Children with other developmental disabilities (n = 21) | Multiple brain-specific autoantibodies are present at significantly higher reactivity in ASD children. |
| Heuer et al., 2008 [20] | Plasma levels of immunoglobulin (IgG, IgM, IgA and IgE) | Children with autism (n = 116) | TD children (n = 96) Children with developmental delays (DD) children (n = 32) Children with ASD but not full autism (n = 27) | Reduced level of IgG in children with autism compared to TD and DD children. Reduced level of IgM in children with autism compared to TD children. IgG and IgM levels were negatively correlated with aberrant behavior checklist scores for all children. |
| Enstrom et al., 2009 * | Circulating plasma levels of IgG1, IgG2, IgG3 and IgG4 | Children with autism (n = 114) | TD children (n = 96) DD children (n = 31) | Increased levels of the IgG4 subclass in children with autism compared to TD children and DD children |
| Li et al., 2009 * | Levels of cytokines in frozen human brain tissue | Children, adolescents and adults with ASD (age: 4 to 37) (n = 8) |
TD children, adolescents, and adults (age: 4 to 39) (n = 8) |
Increased brain levels of proinflammatory cytokines (TNF-α, IL-6 and GM-CSF), Th1 cytokine (IFN-γ) and chemokine (IL-8) in individuals with ASD compared to TD controls. No significant difference for levels of Th2 cytokines (IL-4, IL-5 and IL-10) in the two groups. Increase in the Th1/Th2 ratio in individuals with ASD. |
| Saresalla et al., 2009 [1] | Intracellular cytokines, perforin and granzyme production; monoclonal antibodies, autoantibodies, and virological analysis | Children with autism (n = 20) | Non-autistic siblings (NAS) (n = 15) TD children (n = 20) | Increased proinflammatory and interleukin-10-producing immune cells in children with autism and NAS compared to TD children. Detection of serum autoantibodies (GM1) in 10% of children with autism. |
| Onore et al., 2009 * | Cellular release of IL-17 and IL-23 following an in vitro immunological challenge of peripheral blood mononuclear cells (PBMC) | Children with ASD (n = 34) | TD children (n = 26) | Following stimulation, the concentration of IL-23, but not IL-17, was significantly lower in the ASD group compared to the TD group. |
| Enstrom et al., 2010 [5] | Stimulated cell cultures in vitro of peripheral blood monocytes with distinct Toll-like receptors (TLR) ligands: TLR 2 (lipoteichoic acid; LTA), TLR 3 (poly I:C), TLR 4 (lipopolysaccharide; LPS), TLR 5 (flagellin), and TLR 9 (CpG-B). Pro-inflammatory cytokine responses for IL-1b, IL-6, IL-8, TNFα, MCP-1, and GM-CSF were determined by multiplex Luminex analysis. | Children with ASD (n = 17) | TD children (n = 16) | Increased pro-inflammatory IL-1b, IL-6, and TNFα responses following TLR 2 stimulation, and IL-1b response following TLR 4 stimulation in children with ASD compared to TD children. Decreased IL-1b, IL-6, GM-CSF, and TNFα responses following TLR 9 stimulation in children with ASD compared to TD children. |
| Ashwood et al., 2011 [12] | Assessment plasma cytokine production by multiplex Luminex analysis | Children with ASD (n = 97) | TD children (n = 87) Children with developmental disabilities other than autism (n = 39) | Increased levels of cytokines (IL-1b, IL-6, IL-8 and IL-12p40) in the ASD group, especially for the regressive form of ASD, compared to the TD group. |
| Wei et al., 2011 * | An in vitro adenoviral gene delivery approach was used to over-express IL-6 in cultured cerebellar granule cells. Cell adhesion and migration assays, DiI labeling, TO-PRO-3 staining and immunofluorescence were used to examine cell adhesion and migration, dendritic spine morphology, cell apoptosis and synaptic protein expression, respectively. | Children with autism (n = 6) | TD children (n = 6) | IL-6 was increased in the cerebellum of children with autism. IL-6 over-expression in granule cells caused impairments in granule cell adhesion and migration, and stimulated the formation of granule cell excitatory synapses (without affecting inhibitory synapses), but had little effect on the formation of dendritic spines or granule cell apoptosis. |
| Goines et al., 2011 * | Measured in banked serum of levels of 17 cytokines and chemokines using Luminex technology | Women at 15 to 19 weeks of gestation who gave birth to a child with ASD (n = 84) | Women at 15 to 19 weeks of gestation who gave birth to a DD child (n = 49) Women at 15 to 19 weeks of gestation who gave birth to a TD child (n = 159) | Elevated concentrations of IFN-γ, IL-4 and IL-5 in mid-gestation maternal serum were significantly associated with a 50% increased risk of ASD. Elevated concentrations of IL-2, IL-4 and IL-6 were significantly associated with an increased risk of DD without autism. |
| Al-Ayadhi et al., 2012 * | Serum levels of IL-17A by ELISA | Children with autism (n = 45) | TD children (n = 40) | Abnormally higher serum IL-17A levels in children with autism associated with autism severity. |
| Ricci et al., 2013 * | Serum levels of cytokines and BDNF | Children, adolescents and young adults with ASD (age: 2 to 21) (n = 29) | TD children, adolescents, and young adults (n = 29) | Elevated levels in the ASD group of pro-inflammatory cytokine (including interleukin-1, interleukin-6, interleukin-12, interleukin-23, tumor necrosis factor-α and BDNF). |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tordjman, S.; Charrier, A.; Kazatchkine, M.; Roubertoux, P.; Botbol, M.; Bronsard, G.; Avrameas, S. Natural IgG Anti-F (ab’)2 Autoantibody Activity in Children with Autism. Biomedicines 2023, 11, 715. https://doi.org/10.3390/biomedicines11030715
Tordjman S, Charrier A, Kazatchkine M, Roubertoux P, Botbol M, Bronsard G, Avrameas S. Natural IgG Anti-F (ab’)2 Autoantibody Activity in Children with Autism. Biomedicines. 2023; 11(3):715. https://doi.org/10.3390/biomedicines11030715
Chicago/Turabian StyleTordjman, Sylvie, Annaëlle Charrier, Michel Kazatchkine, Pierre Roubertoux, Michel Botbol, Guillaume Bronsard, and Stratis Avrameas. 2023. "Natural IgG Anti-F (ab’)2 Autoantibody Activity in Children with Autism" Biomedicines 11, no. 3: 715. https://doi.org/10.3390/biomedicines11030715
APA StyleTordjman, S., Charrier, A., Kazatchkine, M., Roubertoux, P., Botbol, M., Bronsard, G., & Avrameas, S. (2023). Natural IgG Anti-F (ab’)2 Autoantibody Activity in Children with Autism. Biomedicines, 11(3), 715. https://doi.org/10.3390/biomedicines11030715