
Lactobacillus plantarum Generate Electricity through Flavin Mononucleotide-Mediated Extracellular Electron Transfer to Upregulate Epithelial Type I Collagen Expression and Thereby Promote Microbial Adhesion to Intestine

 , , , ,
, , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Identification and Cultivation
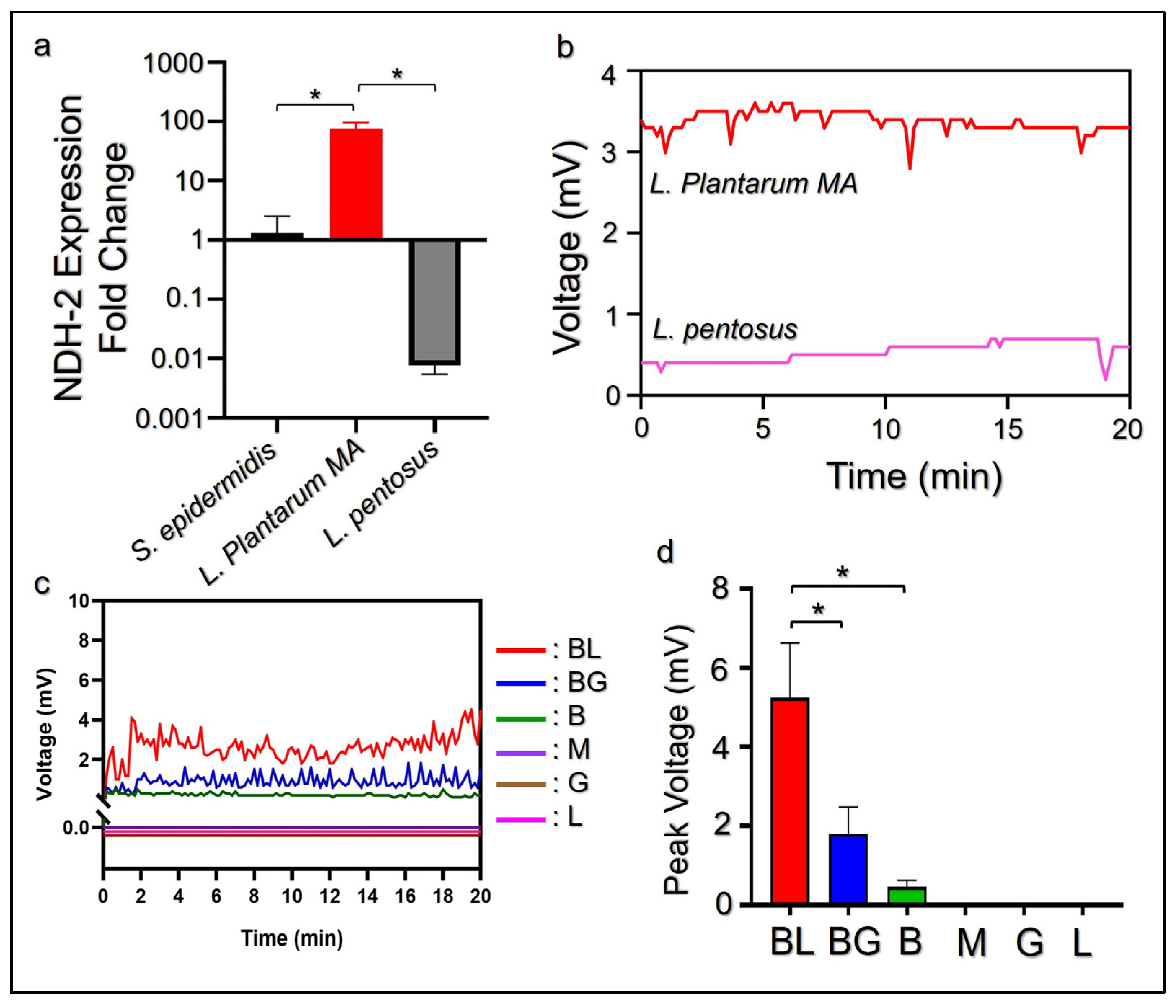
2.2. Quantification of Microbial NDH-2 Expression by RT-qPCR
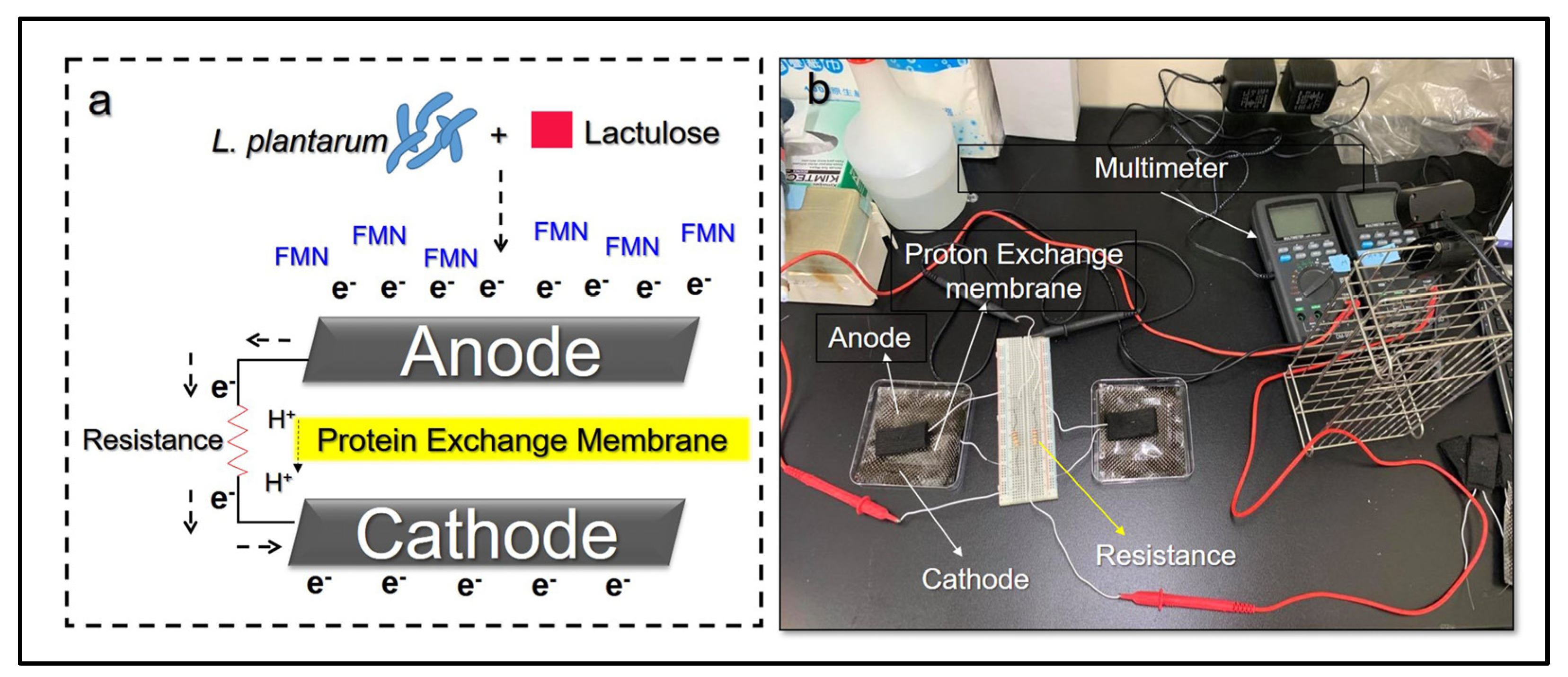
2.3. Detection of Microbial Electricity In Vitro
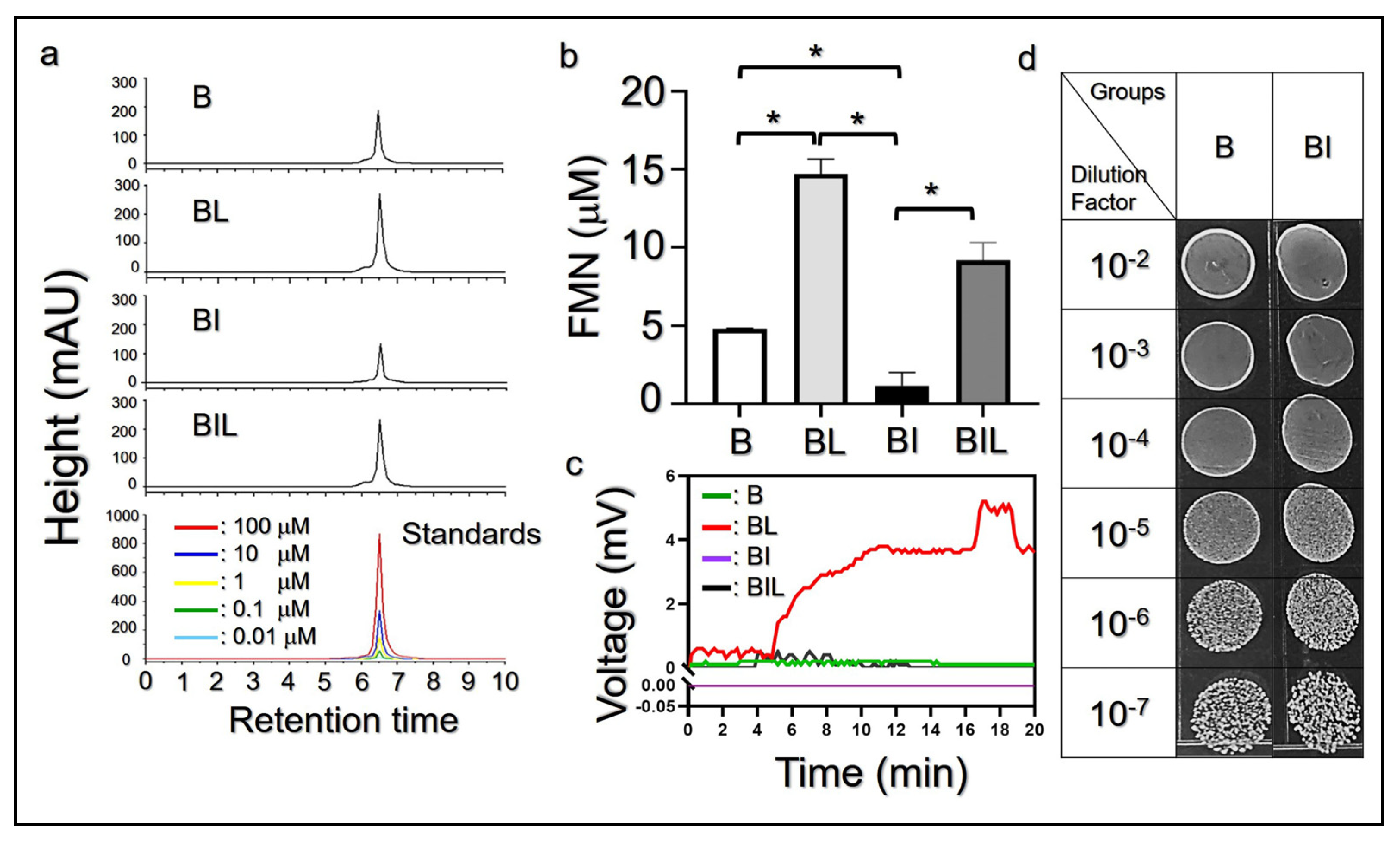
2.4. Detection of Endogenous FMN Yielded from L. plantarum MA
2.5. Cell Culture
2.6. Microbial Adhesion Assay In Vitro
2.7. Ethics Statement for the Animal Study
2.8. Microbial Adhesion Assay Ex Vivo
2.9. Measurement of Col-I Expression of the Cecum after Various Treatments Ex Vivo
2.10. Statistical Analysis
3. Results and Discussion
3.1. Identification and Electrogenic Properties of L. plantarum MA
3.2. Effect of FMN on Electricity Generation of L. plantarum MA
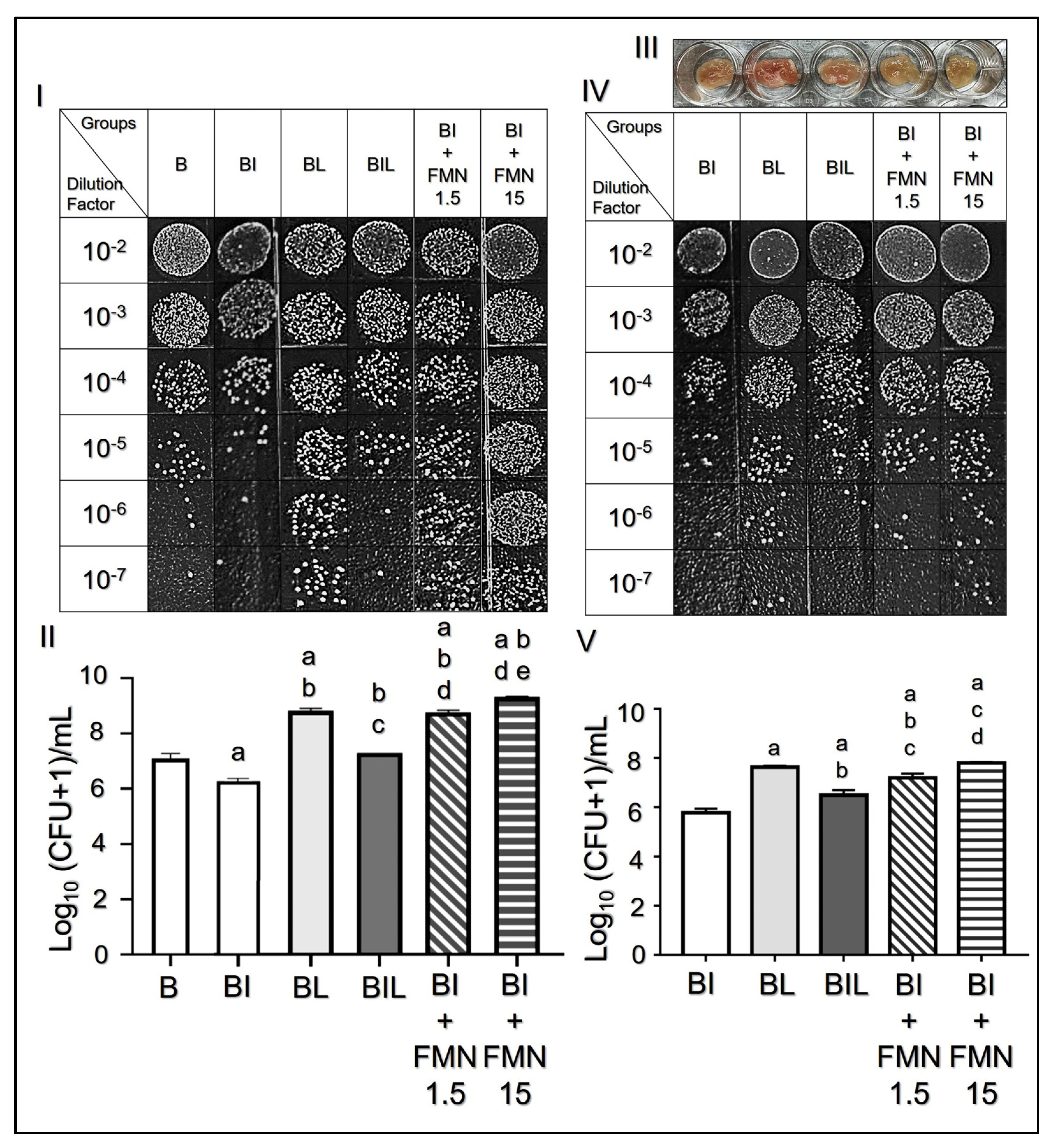
3.3. Effects of FMN-Mediated Electricity on Microbial Adhesion In Vitro and Ex Vivo
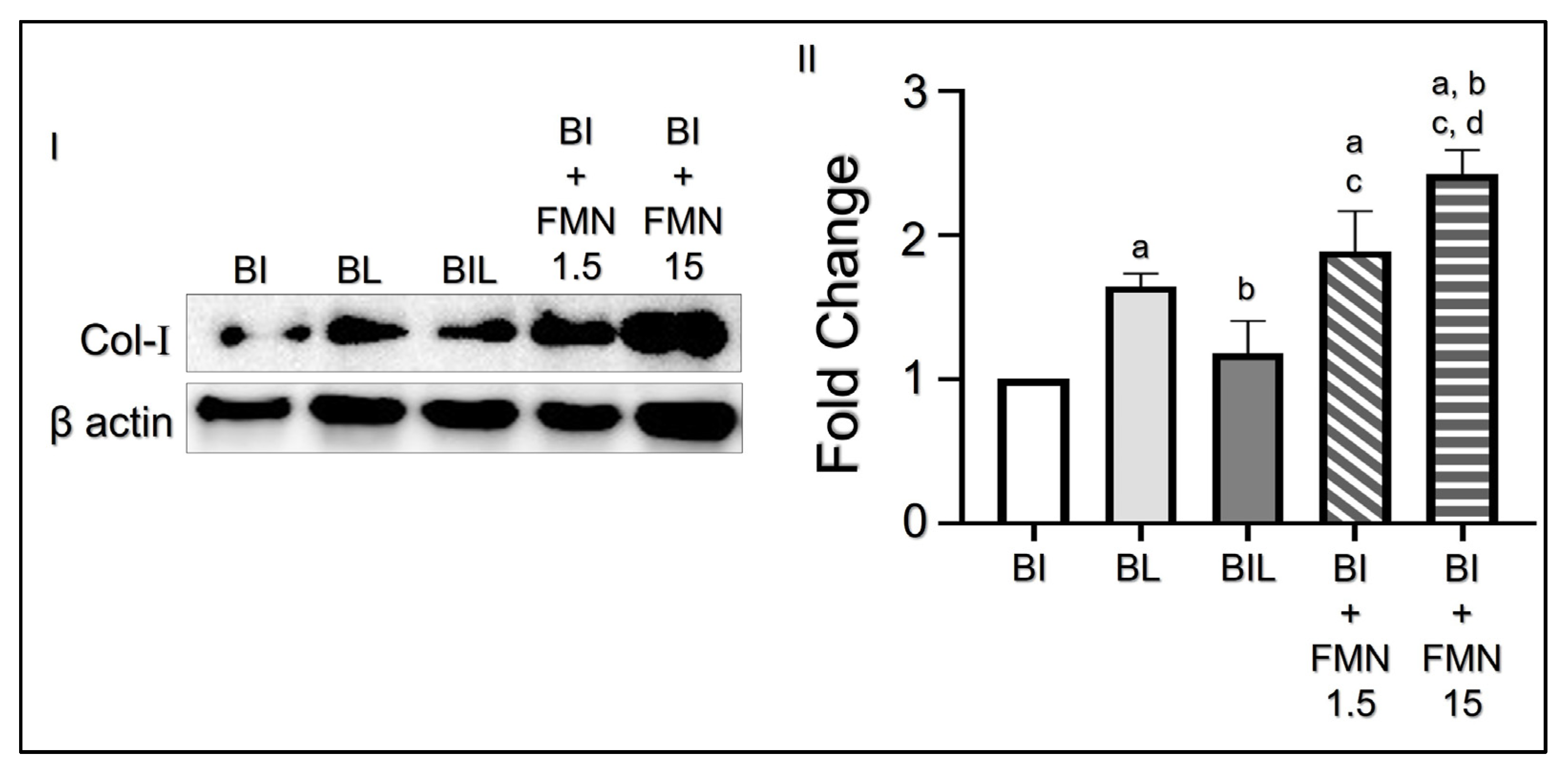
3.4. FMN Upregulates Col-I Collagen Expression on Cecum
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alizadeh Behbahani, B.; Noshad, M.; Falah, F. Inhibition of Escherichia coli adhesion to human intestinal Caco-2 cells by probiotic candidate Lactobacillus plantarum strain L15. Microb. Pathog. 2019, 136, 103677. [Google Scholar] [CrossRef] [PubMed]
- Uugantsetseg, E.; Batjargal, B. Antioxidant activity of probiotic lactic acid bacteria isolated from Mongolian airag. Mong. J. Chem. 2014, 15, 73–78. [Google Scholar] [CrossRef]
- Light, S.H.; Su, L.; Rivera-Lugo, R.; Cornejo, J.A.; Louie, A.; Iavarone, A.T.; Ajo-Franklin, C.M.; Portnoy, D.A. A flavin-based extracellular electron transfer mechanism in diverse Gram-positive bacteria. Nature 2018, 562, 140–144. [Google Scholar] [CrossRef] [PubMed]
- Cahoon, L.A.; Freitag, N.E. The electrifying energy of gut microbes. Nature 2018, 562, 43–44. [Google Scholar] [CrossRef] [PubMed]
- Pham, M.T.; Yang, J.J.; Balasubramaniam, A.; Rahim, A.R.; Adi, P.; Do, T.T.M.; Herr, D.R.; Huang, C.M. Leuconostoc mesenteroides mediates an electrogenic pathway to attenuate the accumulation of abdominal fat mass induced by high fat diet. Sci. Rep. 2020, 10, 21916. [Google Scholar] [CrossRef]
- Schwab, L.; Rago, L.; Koch, C.; Harnisch, F. Identification of Clostridium cochlearium as an electroactive microorganism from the mouse gut microbiome. Bioelectrochemistry 2019, 130, 107334. [Google Scholar] [CrossRef]
- Tian, T.; Fan, X.; Feng, M.; Su, L.; Zhang, W.; Chi, H.; Fu, D. Flavin-mediated extracellular electron transfer in Gram-positive bacteria Bacillus cereus DIF1 and Rhodococcus ruber DIF2. RSC Adv. 2019, 9, 40903–40909. [Google Scholar] [CrossRef]
- Zhang, E.; Cai, Y.; Luo, Y.; Piao, Z. Riboflavin-shuttled extracellular electron transfer from Enterococcus faecalis to electrodes in microbial fuel cells. Can. J. Microbiol. 2014, 60, 753–759. [Google Scholar] [CrossRef]
- Khan, M.T.; Browne, W.R.; van Dijl, J.M.; Harmsen, H.J. How can Faecalibacterium prausnitzii employ riboflavin for extracellular electron transfer? Antioxid. Redox Signal. 2012, 17, 1433–1440. [Google Scholar] [CrossRef]
- Masuda, M.; Freguia, S.; Wang, Y.F.; Tsujimura, S.; Kano, K. Flavins contained in yeast extract are exploited for anodic electron transfer by Lactococcus lactis. Bioelectrochemistry 2010, 78, 173–175. [Google Scholar] [CrossRef]
- Arena, M.P.; Silvain, A.; Normanno, G.; Grieco, F.; Drider, D.; Spano, G.; Fiocco, D. Use of Lactobacillus plantarum Strains as a Bio-Control Strategy against Food-Borne Pathogenic Microorganisms. Front. Microbiol. 2016, 7, 464. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-W.; Liong, M.T.; Tsai, Y.C. New perspectives of Lactobacillus plantarum as a probiotic: The gut-heart-brain axis. J. Microbiol. 2018, 56, 601–613. [Google Scholar] [CrossRef] [PubMed]
- Adlerberth, I.; Cerquetti, M.; Poilane, I.; Wold, A.; Collignon, A. Mechanisms of Colonisation and Colonisation Resistance of the Digestive Tract Part 1: Bacteria/host Interactions. Microb. Ecol. Health Dis. 2000, 12, 223–239. [Google Scholar]
- Graham, M.F.; Diegelmann, R.F.; Elson, C.O.; Lindblad, W.J.; Gotschalk, N.; Gay, S.; Gay, R. Collagen content and types in the intestinal strictures of Crohn’s disease. Gastroenterology 1988, 94, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Wolf, B.; Garleb, K.; Ataya, D.; Casas, I. Safety and tolerance of Lactobacillus reuteri in healthy adult male subjects. Microb. Ecol. Health Dis. 1995, 8, 41–50. [Google Scholar] [CrossRef]
- Aleljung, P.; Shen, W.; Rozalska, B.; Hellman, U.; Ljungh, Å.; Wadström, T. Purification of collagen-binding proteins of Lactobacillus reuteri NCIB 11951. Curr. Microbiol. 1994, 28, 231–236. [Google Scholar] [CrossRef]
- Toba, T.; Virkola, R.; Westerlund, B.; Bjorkman, Y.; Sillanpaa, J.; Vartio, T.; Kalkkinen, N.; Korhonen, T.K. A collagen-binding S-layer protein in Lactobacillus crispatus. Appl. Environ. Microbiol. 1995, 61, 2467–2471. [Google Scholar] [CrossRef]
- Schneitz, C.; Nuotio, L.; Lounatma, K. Adhesion of Lactobacillus acidophilus to avian intestinal epithelial cells mediated by the crystalline bacterial cell surface layer (S-layer). J. Appl. Bacteriol. 1993, 74, 290–294. [Google Scholar] [CrossRef]
- Roos, S.; Aleljung, P.; Robert, N.; Lee, B.; Wadström, T.; Lindberg, M.; Jonsson, H. A collagen binding protein from Lactobacillus reuteri is part of an ABC transporter system? FEMS Microbiol. Lett. 1996, 144, 33–38. [Google Scholar] [CrossRef]
- Sillanpää, J.; Martínez, B.; Antikainen, J.; Toba, T.; Kalkkinen, N.; Tankka, S.; Lounatmaa, K.; Keränen, J.; Höök, M.; Westerlund-Wikström, B.; et al. Characterization of the collagen-binding S-layer protein CbsA of Lactobacillus crispatus. J. Bacteriol. 2000, 182, 6440–6450. [Google Scholar] [CrossRef]
- Mao, B.; Li, D.; Zhao, J.; Liu, X.; Gu, Z.; Chen, Y.Q.; Zhang, H.; Chen, W. In Vitro Fermentation of Lactulose by Human Gut Bacteria. J. Agric. Food Chem. 2014, 62, 10970–10977. [Google Scholar] [CrossRef] [PubMed]
- Salminen, S.; Salminen, E. Lactulose, Lactic Acid Bacteria, Intestinal Microecology and Mucosal Protection. Scand. J. Gastroenterol. 1997, 32, 45–48. [Google Scholar] [CrossRef] [PubMed]
- Calik, A.; Ergün, A. Effect of lactulose supplementation on growth performance, intestinal histomorphology, cecal microbial population, and short-chain fatty acid composition of broiler chickens. J Poult. Sci. 2015, 94, 2173–2182. [Google Scholar] [CrossRef]
- Vilas Boas, J.; Oliveira, V.B.; Marcon, L.R.C.; Simões, M.; Pinto, A.M.F.R. Optimization of a single chamber microbial fuel cell using Lactobacillus pentosus: Influence of design and operating parameters. Sci. Total Environ. 2019, 648, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Balasubramaniam, A.; Adi, P.; Do Thi, T.M.; Yang, J.H.; Labibah, A.S.; Huang, C.M. Skin bacteria mediate glycerol fermentation to produce electricity and resist UV-B. Microorganisms 2020, 8, 1092. [Google Scholar] [CrossRef] [PubMed]
- Brown, K.; Abbott, D.W.; Uwiera, R.R.E.; Inglis, G.D. Removal of the cecum affects intestinal fermentation, enteric bacterial community structure, and acute colitis in mice. Gut Microbes 2018, 9, 218–235. [Google Scholar] [CrossRef] [PubMed]
- Finke, N.; Vandieken, V.; Jørgensen, B.B. Acetate, lactate, propionate, and isobutyrate as electron donors for iron and sulfate reduction in Arctic marine sediments, Svalbard. FEMS Microbiol. Ecol. 2007, 59, 10–22. [Google Scholar] [CrossRef]
- Hernandez-Hernandez, O.; Muthaiyan, A.; Moreno, F.J.; Montilla, A.; Sanz, M.L.; Ricke, S.C. Effect of prebiotic carbohydrates on the growth and tolerance of Lactobacillus. Food Microbiol. 2012, 30, 355–361. [Google Scholar] [CrossRef]
- Kao, M.-S.; Yang, J.H.; Balasubramaniam, A.; Traisaeng, S.; Jackson Yang, A.; Yang, J.J.; Salamon, B.P.; Herr, D.R.; Huang, C.M. Colonization of nasal cavities by Staphylococcus epidermidis mitigates SARS-CoV-2 nucleocapsid phosphoprotein-induced interleukin (IL)-6 in the lung. Microb. Biotechnol. 2022, 15, 1984–1994. [Google Scholar] [CrossRef]
- Mansjö, M.; Johansson, J. The riboflavin analog roseoflavin targets an FMN-riboswitch and blocks Listeria monocytogenes growth, but also stimulates virulence gene-expression and infection. RNA Biol. 2011, 8, 674–680. [Google Scholar] [CrossRef]
- Gomez-Carretero, S.; Libberton, B.; Svennersten, K.; Persson, K.; Jager, E.; Berggren, M.; Rhen, M.; Richter-Dahlfors, A. Redox-active conducting polymers modulate Salmonella biofilm formation by controlling availability of electron acceptors. Npj Biofilms Microbiomes 2017, 3, 19. [Google Scholar] [CrossRef] [PubMed]
- Al-Tawaha, R.; Meng, C. Potential benefits of Lactobacillus plantarum as probiotic and its advantages in human health and industrial applications: A review. Adv. Environ. Biol. 2018, 12, 16–27. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microbial Species | Primers (F: Forward; R: Reverse) |
|---|---|
| L. plantarum MA | F: AACACGCCATCAACTGGATGT R: TTTAGTCCAGCAGTCCCATAACC |
| L. pentosus | F: GCAGAACTCGACCCTGCAAA R: TGCGTCCGATTGAGCATGT |
| S. epidermidis | F: TGGACAGCAGGCATACAACC R: TCTGCTAGTTGAGCACTGGG |
| Microbial Species | Adhesion Target | Treatment (Dose) |
|---|---|---|
| L. plantarum MA | Caco-2 Cells | * None |
| Glucose (2% w/w) | ||
| Lactulose (2% w/w) | ||
| Roseoflavin (1 µM) | ||
| Roseoflavin (1 µM) + Lactulose (2% w/w) | ||
| Roseoflavin (1 µM) + 1.5 µM FMN | ||
| Roseoflavin (1 µM) + 15 µM FMN |
| Microbial Species | Adhesion Target | Treatment (Dose) |
|---|---|---|
| L. plantarum MA | Mice cecum | Roseoflavin (1 µM) |
| Lactulose (2% w/w) | ||
| Roseoflavin (1 µM) + Lactulose (2% w/w) | ||
| Roseoflavin (1 µM) + 1.5 µM FMN | ||
| Roseoflavin (1 µM) + 15 µM FMN |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ganzorig, B.; Zayabaatar, E.; Pham, M.T.; Marito, S.; Huang, C.-M.; Lee, Y.-H. Lactobacillus plantarum Generate Electricity through Flavin Mononucleotide-Mediated Extracellular Electron Transfer to Upregulate Epithelial Type I Collagen Expression and Thereby Promote Microbial Adhesion to Intestine. Biomedicines 2023, 11, 677. https://doi.org/10.3390/biomedicines11030677
Ganzorig B, Zayabaatar E, Pham MT, Marito S, Huang C-M, Lee Y-H. Lactobacillus plantarum Generate Electricity through Flavin Mononucleotide-Mediated Extracellular Electron Transfer to Upregulate Epithelial Type I Collagen Expression and Thereby Promote Microbial Adhesion to Intestine. Biomedicines. 2023; 11(3):677. https://doi.org/10.3390/biomedicines11030677
Chicago/Turabian StyleGanzorig, Binderiya, Enkhbat Zayabaatar, Minh Tan Pham, Shinta Marito, Chun-Ming Huang, and Yu-Hsiang Lee. 2023. "Lactobacillus plantarum Generate Electricity through Flavin Mononucleotide-Mediated Extracellular Electron Transfer to Upregulate Epithelial Type I Collagen Expression and Thereby Promote Microbial Adhesion to Intestine" Biomedicines 11, no. 3: 677. https://doi.org/10.3390/biomedicines11030677
APA StyleGanzorig, B., Zayabaatar, E., Pham, M. T., Marito, S., Huang, C.-M., & Lee, Y.-H. (2023). Lactobacillus plantarum Generate Electricity through Flavin Mononucleotide-Mediated Extracellular Electron Transfer to Upregulate Epithelial Type I Collagen Expression and Thereby Promote Microbial Adhesion to Intestine. Biomedicines, 11(3), 677. https://doi.org/10.3390/biomedicines11030677