
Fibromyalgia in Pregnancy: Neuro-Endocrine Fluctuations Provide Insight into Pathophysiology and Neuromodulation Treatment

 ,
,  ,
,
Abstract
1. Introduction
2. Methods
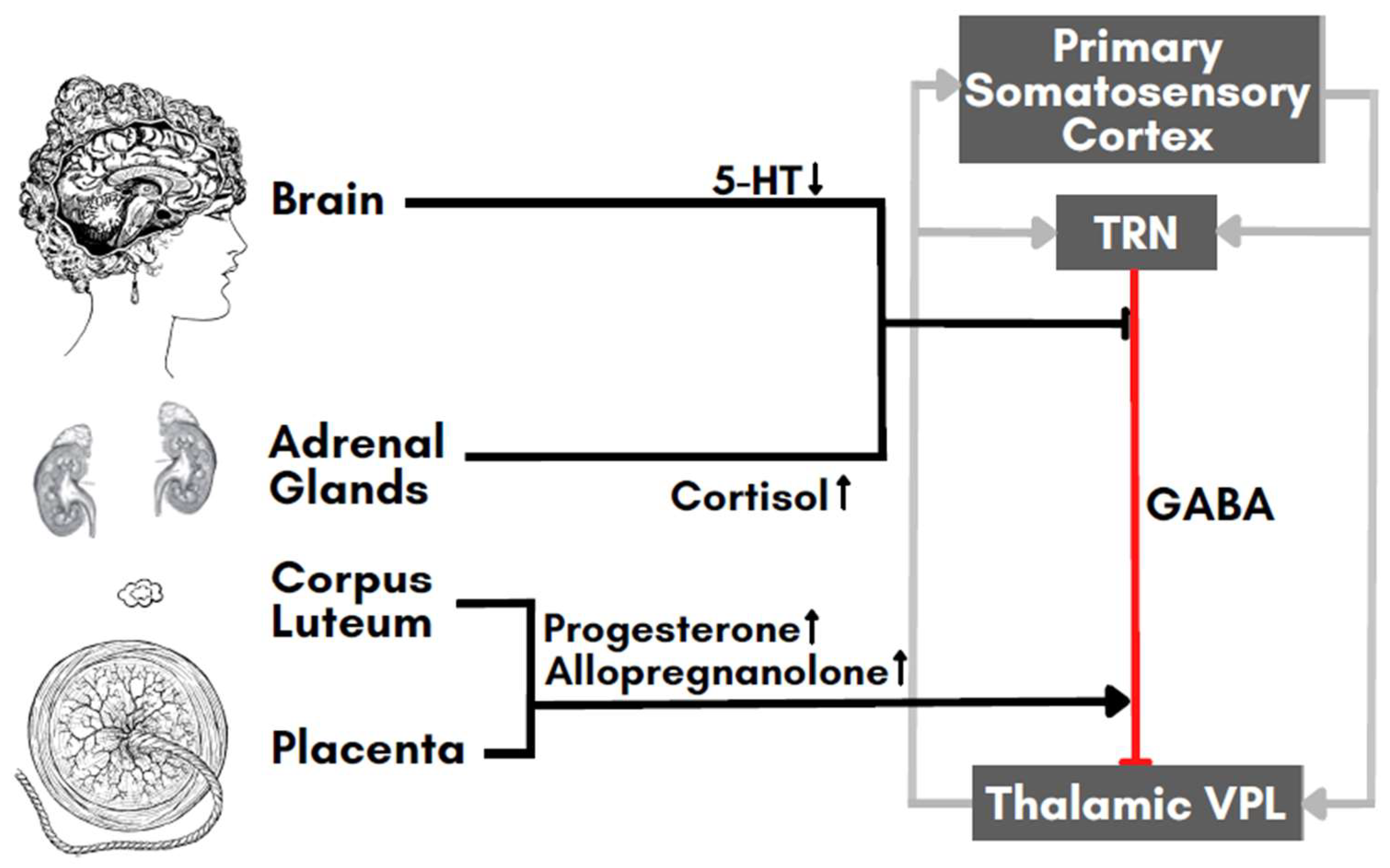
3. Interactions between FM and Pregnancy
4. Estrogen, Progesterone, and Cortisol
5. Prolactin and Immune Mediators
6. Neurotransmitters
7. Perspectives for a Validation of Our Hypothesis
8. Future Interventions for Neuromodulation in Pregnant FM Patients

{kind=link}
| Shenoy et al., 2014 [123] | Sreeraj et al., 2016 [124] | Strube et al., 2016 [125] | Palm et al., 2017 [126] | Vigod et al., 2019 [112] | Kurzeck et al., 2021 [118] | |
| Study Type | Case Report | Case Report | Case Report | Uncontrolled | RCT | Open Label |
| N | 1 | 1 | 1 | 3 | 20 | 6 |
| Age | 25 | 23 | 36 | 23, 28, 32 | >18 | 23–43 |
| Dropouts | 0 | 0 | 0 | 0(1) * | 4 ** | 0(2) * |
| Diagnosis | AH | MDD | AH | MDD | MDD | MDD |
| Scale(s) | PSYRATS | HAMD, HAMA | PANSS, AHRS, CDSS, CGI, GAF | HAMD-21, BDI, TMT-A/B | MADRS | HAMD-21, BDI, CGI, TMT-A/B |
| Treatment | Add-On | Mono | Mono | Mono | Mono | Mono |
| tDCS prior to Pregnancy | Yes | No | N/A | N/A | N/A | N/A |
| Weeks in Gestation | 18 | 6 | 32 | 19–31 | 14–32 | 12–33 |
| Parameters | 2 mA, 2 × 20 min | 2 mA, 30 min | 2 mA, 2 × 30 min | 2 mA, 2 × 30 min (2 mA, 30 min) | 2 mA, 30 min | 2 mA, 2 × 30 min (2 mA, 30 min) |
| No. of Sessions | 10 | 10 | 20 | 20 (30) | 15 | 20 (30) |
| Response | Near remission | Remission | 41% improvement (CDSS) | 33.3% remission | 75% vs. 12.5% | 39.3% reduction (HAMD) 57.1% reduction (BDI) 28.6% reduction (CGI) |
| Comments | Add-on tDCS resulted in near remission of auditory hallucination. tDCS was well tolerated and no changes in autonomic function, ventilation rate, or core body temperature were observed. | tDCS was well tolerated without any adverse events. In 3 out of the 10 tDCS sessions, patients experienced transient, mild burning sensation at the target side and fleeting perception of phosphenes during the fade-in phase, which is an anticipated tDCS side effect. | No improvement in auditory hallucinations was recorded. Patients tolerated tDCS well with no reported, noticeable side effects. Fetal examination at 35th gestational week revealed no changes or abnormalities. Delivery of a healthy child occurred with no complications. | Statistically significant changes could be observed. One patient achieved remission. tDCS was well tolerated without adverse events. | No abnormalities or serious pregnancy complications were reported in either group. Percent fractions of 87.5% and 77.8% in the tDCS group and sham group, respectively, were satisfied to extremely satisfied with the treatment and viewed tDCS as an acceptable and alternative treatment option. | Significant changes were observed. tDCS was well tolerated without adverse events. In Phase 1, 33.3% achieved response in HAMD scores; 33.3% showed response and 16.7% remission in BDI scores. In Phase 2, one patient achieved remission for both HAMD and BDI. |
- Perform hormonal-related longitudinal studies in FM patients during different hormonal phases: menses, pregnancy, menopause. Progesterone, prolactin, estrogen, and testosterone should be carefully evaluated and matched with symptom fluctuations.
- Further assess the involvement of cortisol and serotonin through repetitive blood and saliva sampling.
- Assess the feasibility of tDCS in women planning a pregnancy affected by FM.
- Assess home-based, remote tDCS treatment in combination with lifestyle changes, given they have been proven to be successful at reducing the patient’s symptoms [33].
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kia, S.; Choy, E. Update on Treatment Guideline in Fibromyalgia Syndrome with Focus on Pharmacology. Biomedicines 2017, 5, 20. [Google Scholar] [CrossRef] [PubMed]
- Boomershine, C.S. Fibromyalgia: The prototypical central sensitivity syndrome. Curr. Rheumatol. Rev. 2015, 11, 131–145. [Google Scholar] [CrossRef] [PubMed]
- Alciati, A.; Nucera, V.; Masala, I.F.; Giallanza, M.; La Corte, L.; Giorgi, V.; Sarzi-Puttini, P.; Atzeni, F. One year in review 2021: Fibromyalgia. Clin. Exp. Rheumatol. 2021, 39 (Suppl. 130), 3–12. [Google Scholar] [CrossRef] [PubMed]
- Hudson, J.I.; Goldenberg, D.L.; Pope, H.G., Jr.; Keck, P.E., Jr.; Schlesinger, L. Comorbidity of fibromyalgia with medical and psychiatric disorders. Am. J. Med. 1992, 92, 363–367. [Google Scholar] [CrossRef] [PubMed]
- Pieretti, S.; Di Giannuario, A.; Di Giovannandrea, R.; Marzoli, F.; Piccaro, G.; Minosi, P.; Aloisi, A.M. Gender differences in pain and its relief. Ann. Ist. Super. Sanita 2016, 52, 184–189. [Google Scholar] [CrossRef]
- Arout, C.A.; Sofuoglu, M.; Bastian, L.A.; Rosenheck, R.A. Gender Differences in the Prevalence of Fibromyalgia and in Concomitant Medical and Psychiatric Disorders: A National Veterans Health Administration Study. J. Womens Health 2018, 27, 1035–1044. [Google Scholar] [CrossRef]
- Clauw, D.J. Fibromyalgia: An overview. Am. J. Med. 2009, 122, S3–S13. [Google Scholar] [CrossRef]
- McNally, J.D.; Matheson, D.A.; Bakowsky, V.S. The epidemiology of self-reported fibromyalgia in Canada. Chronic Dis. Can. 2006, 27, 9–16. [Google Scholar]
- McLeod, J.D. Juvenile fibromyalgia syndrome and improved recognition by pediatric primary care providers. J. Pediatr. Health Care 2014, 28, e9–e18. [Google Scholar] [CrossRef]
- Rossi, M.F.; Tumminello, A.; Marconi, M.; Gualano, M.R.; Santoro, P.E.; Malorni, W.; Moscato, U. Sex and gender differences in migraines: A narrative review. Neurol. Sci. 2022, 43, 5729–5734. [Google Scholar] [CrossRef]
- Abu-Raya, B.; Michalski, C.; Sadarangani, M.; Lavoie, P.M. Maternal Immunological Adaptation During Normal Pregnancy. Front. Immunol. 2020, 11, 575197. [Google Scholar] [CrossRef] [PubMed]
- Staud, R. Cytokine and immune system abnormalities in fibromyalgia and other central sensitivity syndromes. Curr. Rheumatol. Rev. 2015, 11, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Goebel, A.; Krock, E.; Gentry, C.; Israel, M.R.; Jurczak, A.; Urbina, C.M.; Sandor, K.; Vastani, N.; Maurer, M.; Cuhadar, U.; et al. Passive transfer of fibromyalgia symptoms from patients to mice. J. Clin. Investig. 2021, 131. [Google Scholar] [CrossRef] [PubMed]
- Wise, P.M. Estrogens and neuroprotection. Trends Endocrinol. Metab. 2002, 13, 229–230. [Google Scholar] [CrossRef]
- Roof, R.L.; Hall, E.D. Gender differences in acute CNS trauma and stroke: Neuroprotective effects of estrogen and progesterone. J. Neurotrauma 2000, 17, 367–388. [Google Scholar] [CrossRef]
- Brinton, R.D.; Thompson, R.F.; Foy, M.R.; Baudry, M.; Wang, J.; Finch, C.E.; Morgan, T.E.; Pike, C.J.; Mack, W.J.; Stanczyk, F.Z.; et al. Progesterone receptors: Form and function in brain. Front. Neuroendocr. 2008, 29, 313–339. [Google Scholar] [CrossRef]
- Brinton, R.D. Cellular and molecular mechanisms of estrogen regulation of memory function and neuroprotection against Alzheimer’s disease: Recent insights and remaining challenges. Learn. Mem. 2001, 8, 121–133. [Google Scholar] [CrossRef]
- Wiesenfeld-Hallin, Z. Sex differences in pain perception. Gend. Med. 2005, 2, 137–145. [Google Scholar] [CrossRef]
- Aloisi, A.M.; Sorda, G. Relationship of female sex hormones with pain perception: Focus on estrogens. Pain Manag. 2011, 1, 229–238. [Google Scholar] [CrossRef]
- Haus, E.; Smolensky, M.H. Biologic rhythms in the immune system. Chronobiol. Int. 1999, 16, 581–622. [Google Scholar] [CrossRef]
- Taneja, V. Sex Hormones Determine Immune Response. Front. Immunol. 2018, 9, 1931. [Google Scholar] [CrossRef] [PubMed]
- Gregus, A.M.; Levine, I.S.; Eddinger, K.A.; Yaksh, T.L.; Buczynski, M.W. Sex differences in neuroimmune and glial mechanisms of pain. Pain 2021, 162, 2186–2200. [Google Scholar] [CrossRef] [PubMed]
- Sorge, R.E.; Mapplebeck, J.C.; Rosen, S.; Beggs, S.; Taves, S.; Alexander, J.K.; Martin, L.J.; Austin, J.S.; Sotocinal, S.G.; Chen, D.; et al. Different immune cells mediate mechanical pain hypersensitivity in male and female mice. Nat. Neurosci. 2015, 18, 1081–1083. [Google Scholar] [CrossRef] [PubMed]
- Meester, I.; Rivera-Silva, G.F.; Gonzalez-Salazar, F. Immune System Sex Differences May Bridge the Gap Between Sex and Gender in Fibromyalgia. Front. Neurosci. 2019, 13, 1414. [Google Scholar] [CrossRef]
- Demori, I.; Giordano, G.; Mucci, V.; Losacco, S.; Marinelli, L.; Massobrio, P.; Blanchini, F.; Burlando, B. Thalamocortical bistable switch as a theoretical model of fibromyalgia pathogenesis inferred from a literature survey. J. Comput. Neurosci. 2022, 50, 471–484. [Google Scholar] [CrossRef]
- Jones, E.G. Anatomy of cerebral cortex: Columnar input-output organization. In The Organization of the Cerebral Cortex; Schmitt, F.O., Worden, F.G., Adelmann, G., Dennis, S.G., Eds.; MIT Press: Cambridge, MA, USA, 1981; pp. 199–235. [Google Scholar]
- Li, C.; Lei, Y.; Tian, Y.; Xu, S.; Shen, X.; Wu, H.; Bao, S.; Wang, F. The etiological contribution of GABAergic plasticity to the pathogenesis of neuropathic pain. Mol. Pain 2019, 15, 1744806919847366. [Google Scholar] [CrossRef]
- Foerster, B.R.; Petrou, M.; Edden, R.A.; Sundgren, P.C.; Schmidt-Wilcke, T.; Lowe, S.E.; Harte, S.E.; Clauw, D.J.; Harris, R.E. Reduced insular gamma-aminobutyric acid in fibromyalgia. Arthritis. Rheum. 2012, 64, 579–583. [Google Scholar] [CrossRef]
- Sluka, K.A.; Clauw, D.J. Neurobiology of fibromyalgia and chronic widespread pain. Neuroscience 2016, 338, 114–129. [Google Scholar] [CrossRef]
- Pomares, F.B.; Roy, S.; Funck, T.; Feier, N.A.; Thiel, A.; Fitzcharles, M.A.; Schweinhardt, P. Upregulation of cortical GABAA receptor concentration in fibromyalgia. Pain 2020, 161, 74–82. [Google Scholar] [CrossRef]
- Becker, S.; Schweinhardt, P. Dysfunctional neurotransmitter systems in fibromyalgia, their role in central stress circuitry and pharmacological actions on these systems. Pain Res. Treat. 2012, 2012, 741746. [Google Scholar] [CrossRef]
- Vanderwall, A.G.; Milligan, E.D. Cytokines in Pain: Harnessing Endogenous Anti-Inflammatory Signaling for Improved Pain Management. Front. Immunol. 2019, 10, 3009. [Google Scholar] [CrossRef] [PubMed]
- Demori, I.; Molinari, E.; Rapallo, F.; Mucci, V.; Marinelli, L.; Losacco, S.; Burlando, B. Online Questionnaire with Fibromyalgia Patients Reveals Correlations among Type of Pain, Psychological Alterations, and Effectiveness of Non-Pharmacological Therapies. Healthcare 2022, 10, 1975. [Google Scholar] [CrossRef] [PubMed]
- Sackeim, H.A.; Aaronson, S.T.; Carpenter, L.L.; Hutton, T.M.; Mina, M.; Pages, K.; Verdoliva, S.; West, W.S. Clinical outcomes in a large registry of patients with major depressive disorder treated with Transcranial Magnetic Stimulation. J. Affect. Disord. 2020, 277, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Kedzior, K.K.; Azorina, V.; Reitz, S.K. More female patients and fewer stimuli per session are associated with the short-term antidepressant properties of repetitive transcranial magnetic stimulation (rTMS): A meta-analysis of 54 sham-controlled studies published between 1997-2013. Neuropsychiatr. Dis. Treat. 2014, 10, 727–756. [Google Scholar] [CrossRef]
- Zuo, X.N.; Kelly, C.; Di Martino, A.; Mennes, M.; Margulies, D.S.; Bangaru, S.; Grzadzinski, R.; Evans, A.C.; Zang, Y.F.; Castellanos, F.X.; et al. Growing together and growing apart: Regional and sex differences in the lifespan developmental trajectories of functional homotopy. J. Neurosci. 2010, 30, 15034–15043. [Google Scholar] [CrossRef]
- Scheinost, D.; Finn, E.S.; Tokoglu, F.; Shen, X.; Papademetris, X.; Hampson, M.; Constable, R.T. Sex differences in normal age trajectories of functional brain networks. Hum. Brain Mapp. 2015, 36, 1524–1535. [Google Scholar] [CrossRef]
- Ruigrok, A.N.; Salimi-Khorshidi, G.; Lai, M.C.; Baron-Cohen, S.; Lombardo, M.V.; Tait, R.J.; Suckling, J. A meta-analysis of sex differences in human brain structure. Neurosci. Biobehav. Rev. 2014, 39, 34–50. [Google Scholar] [CrossRef]
- Cosgrove, K.P.; Mazure, C.M.; Staley, J.K. Evolving knowledge of sex differences in brain structure, function, and chemistry. Biol. Psychiatry 2007, 62, 847–855. [Google Scholar] [CrossRef]
- Luders, E.; Narr, K.L.; Thompson, P.M.; Rex, D.E.; Jancke, L.; Steinmetz, H.; Toga, A.W. Gender differences in cortical complexity. Nat. Neurosci. 2004, 7, 799–800. [Google Scholar] [CrossRef]
- Smith, M.J.; Adams, L.F.; Schmidt, P.J.; Rubinow, D.R.; Wassermann, E.M. Effects of ovarian hormones on human cortical excitability. Ann. Neurol. 2002, 51, 599–603. [Google Scholar] [CrossRef]
- Inghilleri, M.; Conte, A.; Curra, A.; Frasca, V.; Lorenzano, C.; Berardelli, A. Ovarian hormones and cortical excitability. An rTMS study in humans. Clin. Neurophysiol. 2004, 115, 1063–1068. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.J.; Keel, J.C.; Greenberg, B.D.; Adams, L.F.; Schmidt, P.J.; Rubinow, D.A.; Wassermann, E.M. Menstrual cycle effects on cortical excitability. Neurology 1999, 53, 2069–2072. [Google Scholar] [CrossRef]
- Bunai, T.; Hirosawa, T.; Kikuchi, M.; Fukai, M.; Yokokura, M.; Ito, S.; Takata, Y.; Terada, T.; Ouchi, Y. tDCS-induced modulation of GABA concentration and dopamine release in the human brain: A combination study of magnetic resonance spectroscopy and positron emission tomography. Brain Stimul. 2021, 14, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Saa’d, S.; Many, A.; Jacob, G.; Ablin, J.N. High prevalence of fibromyalgia symptoms among healthy full-term pregnant women. Rheumatol. Int. 2013, 33, 1555–1560. [Google Scholar] [CrossRef]
- Koné, M.C.; Kambiré, N.A.; Kouakou, K.; Ahoua, Y. Fibromyalgia of Women Who Gave Birth and Pregnancy Outcome Parameters. Open J. Epidemiol. 2022, 12, 1–11. [Google Scholar] [CrossRef]
- Zioni, T.; Buskila, D.; Aricha-Tamir, B.; Wiznitzer, A.; Sheiner, E. Pregnancy outcome in patients with fibromyalgia syndrome. J. Matern Fetal Neonatal. Med. 2011, 24, 1325–1328. [Google Scholar] [CrossRef] [PubMed]
- Genc, H.; Atasever, M.; Duyur Cakit, B.; Seval, M.; Koc, A. The Effects of Fibromyalgia Syndrome on Physical Function and Psychological Status of Pregnant Females. Arch. Rheumatol. 2017, 32, 129–140. [Google Scholar] [CrossRef]
- Ostensen, M.; Rugelsjoen, A.; Wigers, S.H. The effect of reproductive events and alterations of sex hormone levels on the symptoms of fibromyalgia. Scand. J. Rheumatol. 1997, 26, 355–360. [Google Scholar] [CrossRef]
- Backstrom, T.; Haage, D.; Lofgren, M.; Johansson, I.M.; Stromberg, J.; Nyberg, S.; Andreen, L.; Ossewaarde, L.; van Wingen, G.A.; Turkmen, S.; et al. Paradoxical effects of GABA-A modulators may explain sex steroid induced negative mood symptoms in some persons. Neuroscience 2011, 191, 46–54. [Google Scholar] [CrossRef]
- Alonso, C.; Loevinger, B.L.; Muller, D.; Coe, C.L. Menstrual cycle influences on pain and emotion in women with fibromyalgia. J. Psychosom. Res. 2004, 57, 451–458. [Google Scholar] [CrossRef]
- Gur, A.; Cevik, R.; Sarac, A.J.; Colpan, L.; Em, S. Hypothalamic-pituitary-gonadal axis and cortisol in young women with primary fibromyalgia: The potential roles of depression, fatigue, and sleep disturbance in the occurrence of hypocortisolism. Ann. Rheum. Dis. 2004, 63, 1504–1506. [Google Scholar] [CrossRef]
- Dunnett, A.J.; Roy, D.; Stewart, A.; McPartland, J.M. The diagnosis of fibromyalgia in women may be influenced by menstrual cycle phase. J. Bodyw. Mov. Ther. 2007, 11, 99–105. [Google Scholar] [CrossRef]
- Chen, Q.; Zhang, W.; Sadana, N.; Chen, X. Estrogen receptors in pain modulation: Cellular signaling. Biol. Sex Differ. 2021, 12, 22. [Google Scholar] [CrossRef] [PubMed]
- Aloisi, A.M.; Bachiocco, V.; Costantino, A.; Stefani, R.; Ceccarelli, I.; Bertaccini, A.; Meriggiola, M.C. Cross-sex hormone administration changes pain in transsexual women and men. Pain 2007, 132 (Suppl. 1), S60–S67. [Google Scholar] [CrossRef] [PubMed]
- Vacca, V.; Marinelli, S.; Pieroni, L.; Urbani, A.; Luvisetto, S.; Pavone, F. 17beta-estradiol counteracts neuropathic pain: A behavioural, immunohistochemical, and proteomic investigation on sex-related differences in mice. Sci. Rep. 2016, 6, 18980. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.N.; McFarland, K.; Olsson, R.; Burstein, E.S. Estrogen Receptor Beta Selective Agonists as Agents to Treat Chemotherapeutic-Induced Neuropathic Pain. ACS Chem. Neurosci. 2016, 7, 1180–1187. [Google Scholar] [CrossRef]
- Schertzinger, M.; Wesson-Sides, K.; Parkitny, L.; Younger, J. Daily Fluctuations of Progesterone and Testosterone Are Associated With Fibromyalgia Pain Severity. J. Pain 2018, 19, 410–417. [Google Scholar] [CrossRef] [PubMed]
- Lambert, J.J.; Cooper, M.A.; Simmons, R.D.; Weir, C.J.; Belelli, D. Neurosteroids: Endogenous allosteric modulators of GABA(A) receptors. Psychoneuroendocrinology 2009, 34 (Suppl. 1), S48–S58. [Google Scholar] [CrossRef]
- Abbassi-Ghanavati, M.; Greer, L.G.; Cunningham, F.G. Pregnancy and laboratory studies: A reference table for clinicians. Obs. Gynecol. 2009, 114, 1326–1331. [Google Scholar] [CrossRef]
- Haldeman-Englert, C.; Turley, R.; Novick, T. Health Encyclopedia. Available online: https://www.urmc.rochester.edu/encyclopedia/content.aspx?ContentTypeID=167&ContentID=progesterone (accessed on 15 December 2022).
- Solano, M.E.; Arck, P.C. Steroids, Pregnancy and Fetal Development. Front. Immunol. 2019, 10, 3017. [Google Scholar] [CrossRef]
- Carr, B.R.; Parker, C.R., Jr.; Madden, J.D.; MacDonald, P.C.; Porter, J.C. Maternal plasma adrenocorticotropin and cortisol relationships throughout human pregnancy. Am. J. Obs. Gynecol. 1981, 139, 416–422. [Google Scholar] [CrossRef] [PubMed]
- Wieczorek, A.; Perani, C.V.; Nixon, M.; Constancia, M.; Sandovici, I.; Zazara, D.E.; Leone, G.; Zhang, M.Z.; Arck, P.C.; Solano, M.E. Sex-specific regulation of stress-induced fetal glucocorticoid surge by the mouse placenta. Am. J. Physiol. Endocrinol. Metab. 2019, 317, E109–E120. [Google Scholar] [CrossRef] [PubMed]
- Brunton, P.J. Neuroactive steroids and stress axis regulation: Pregnancy and beyond. J. Steroid. Biochem. Mol. Biol. 2016, 160, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Brunton, P.J.; McKay, A.J.; Ochedalski, T.; Piastowska, A.; Rebas, E.; Lachowicz, A.; Russell, J.A. Central opioid inhibition of neuroendocrine stress responses in pregnancy in the rat is induced by the neurosteroid allopregnanolone. J. Neurosci. 2009, 29, 6449–6460. [Google Scholar] [CrossRef]
- Groeneweg, F.L.; Karst, H.; de Kloet, E.R.; Joels, M. Rapid non-genomic effects of corticosteroids and their role in the central stress response. J. Endocrinol. 2011, 209, 153–167. [Google Scholar] [CrossRef]
- Shen, H.; Gong, Q.H.; Aoki, C.; Yuan, M.; Ruderman, Y.; Dattilo, M.; Williams, K.; Smith, S.S. Reversal of neurosteroid effects at alpha4beta2delta GABAA receptors triggers anxiety at puberty. Nat. Neurosci. 2007, 10, 469–477. [Google Scholar] [CrossRef]
- Fries, E.; Hesse, J.; Hellhammer, J.; Hellhammer, D.H. A new view on hypocortisolism. Psychoneuroendocrinology 2005, 30, 1010–1016. [Google Scholar] [CrossRef]
- Griep, E.N.; Boersma, J.W.; Lentjes, E.G.; Prins, A.P.; van der Korst, J.K.; de Kloet, E.R. Function of the hypothalamic-pituitary-adrenal axis in patients with fibromyalgia and low back pain. J. Rheumatol. 1998, 25, 1374–1381. [Google Scholar]
- Soma-Pillay, P.; Nelson-Piercy, C.; Tolppanen, H.; Mebazaa, A. Physiological changes in pregnancy. Cardiovasc. J. Afr. 2016, 27, 89–94. [Google Scholar] [CrossRef]
- Barnea, E.R.; Fares, F.; Shahar, K. Stimulatory effect of prolactin on human placental progesterone secretion at term in vitro: Possible inhibitory effect on oestradiol secretion. Placenta 1989, 10, 37–43. [Google Scholar] [CrossRef]
- Levine, S.; Muneyyirci-Delale, O. Stress-Induced Hyperprolactinemia: Pathophysiology and Clinical Approach. Obs. Gynecol. Int. 2018, 2018, 9253083. [Google Scholar] [CrossRef]
- Buskila, D.; Sukenik, S.; Shoenfeld, Y. The possible role of prolactin in autoimmunity. Am. J. Reprod. Immunol. 1991, 26, 118–123. [Google Scholar] [CrossRef] [PubMed]
- Swart, J.M.; Grattan, D.R.; Ladyman, S.R.; Brown, R.S.E. Changes in maternal motivation across reproductive states in mice: A role for prolactin receptor activation on GABA neurons. Horm. Behav. 2021, 135, 105041. [Google Scholar] [CrossRef] [PubMed]
- Locatelli, V.; Apud, J.A.; Gudelsky, G.A.; Cocchi, D.; Masotto, C.; Casanueva, F.; Racagni, G.; Muller, E.E. Prolactin in cerebrospinal fluid increases the synthesis and release of hypothalamic gamma-aminobutyric acid. J. Endocrinol. 1985, 106, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Bole-Feysot, C.; Goffin, V.; Edery, M.; Binart, N.; Kelly, P.A. Prolactin (PRL) and its receptor: Actions, signal transduction pathways and phenotypes observed in PRL receptor knockout mice. Endocr. Rev. 1998, 19, 225–268. [Google Scholar] [CrossRef]
- Maddipati, K.R. Non-inflammatory Physiology of “Inflammatory” Mediators—Unalamation, a New Paradigm. Front. Immunol. 2020, 11, 580117. [Google Scholar] [CrossRef]
- Jarmund, A.H.; Giskeodegard, G.F.; Ryssdal, M.; Steinkjer, B.; Stokkeland, L.M.T.; Madssen, T.S.; Stafne, S.N.; Stridsklev, S.; Moholdt, T.; Heimstad, R.; et al. Cytokine Patterns in Maternal Serum From First Trimester to Term and Beyond. Front. Immunol. 2021, 12, 752660. [Google Scholar] [CrossRef]
- Mor, G.; Aldo, P.; Alvero, A.B. The unique immunological and microbial aspects of pregnancy. Nat. Rev. Immunol. 2017, 17, 469–482. [Google Scholar] [CrossRef]
- Stelzer, I.A.; Arck, P.C. Immunity and the Endocrine System. Encycl. Immunobiol. 2016, 5, 73–85. [Google Scholar]
- Uceyler, N.; Hauser, W.; Sommer, C. Systematic review with meta-analysis: Cytokines in fibromyalgia syndrome. BMC Musculoskelet. Disord. 2011, 12, 245. [Google Scholar] [CrossRef]
- Al-Nimer, M.S.M.; Mohammad, T.A.M.; Alsakeni, R.A. Serum levels of serotonin as a biomarker of newly diagnosed fibromyalgia in women: Its relation to the platelet indices. J. Res. Med. Sci. 2018, 23, 71. [Google Scholar] [CrossRef] [PubMed]
- Hardeland, R. Neurobiology, pathophysiology, and treatment of melatonin deficiency and dysfunction. Sci. World J. 2012, 2012, 640389. [Google Scholar] [CrossRef] [PubMed]
- O’Malley, P.G.; Balden, E.; Tomkins, G.; Santoro, J.; Kroenke, K.; Jackson, J.L. Treatment of fibromyalgia with antidepressants: A meta-analysis. J. Gen. Intern. Med. 2000, 15, 659–666. [Google Scholar] [CrossRef] [PubMed]
- Cheng, K.J. Neurobiological mechanisms of acupuncture for some common illnesses: A clinician’s perspective. J. Acupunct. Meridian Stud. 2014, 7, 105–114. [Google Scholar] [CrossRef]
- Zhao, Z.Q. Neural mechanism underlying acupuncture analgesia. Prog. Neurobiol. 2008, 85, 355–375. [Google Scholar] [CrossRef]
- Goldenberg, D.L.; Clauw, D.J.; Palmer, R.E.; Clair, A.G. Opioid Use in Fibromyalgia: A Cautionary Tale. Mayo Clin. Proc. 2016, 91, 640–648. [Google Scholar] [CrossRef]
- Munsch, T.; Freichel, M.; Flockerzi, V.; Pape, H.C. Contribution of transient receptor potential channels to the control of GABA release from dendrites. Proc. Natl. Acad. Sci. USA 2003, 100, 16065–16070. [Google Scholar] [CrossRef]
- Eaton, S.A.; Salt, T.E. Modulatory effects of serotonin on excitatory amino acid responses and sensory synaptic transmission in the ventrobasal thalamus. Neuroscience 1989, 33, 285–292. [Google Scholar] [CrossRef]
- Smolen, A.; Smolen, T.N.; Han, P.C. Alterations in regional brain GABA concentration and turnover during pregnancy. Pharm. Biochem. Behav. 1993, 44, 63–69. [Google Scholar] [CrossRef]
- Maguire, J.; Mody, I. GABA(A)R plasticity during pregnancy: Relevance to postpartum depression. Neuron 2008, 59, 207–213. [Google Scholar] [CrossRef]
- Atasever, M.; Kalem, M.N.; Sonmez, C.; Seval, M.M.; Yuce, T.; Aker, S.S.; Koc, A.; Genc, H. Lower serotonin level and higher rate of fibromyalgia syndrome with advancing pregnancy. J. Matern. Fetal Neonatal Med. 2017, 30, 2204–2211. [Google Scholar] [CrossRef] [PubMed]
- Coluzzi, F.; Valensise, H.; Sacco, M.; Allegri, M. Chronic pain management in pregnancy and lactation. Minerva Anestesiol 2014, 80, 211–224. [Google Scholar] [PubMed]
- Concas, A.; Mostallino, M.C.; Porcu, P.; Follesa, P.; Barbaccia, M.L.; Trabucchi, M.; Purdy, R.H.; Grisenti, P.; Biggio, G. Role of brain allopregnanolone in the plasticity of gamma-aminobutyric acid type A receptor in rat brain during pregnancy and after delivery. Proc. Natl. Acad. Sci. USA 1998, 95, 13284–13289. [Google Scholar] [CrossRef] [PubMed]
- Follesa, P.; Serra, M.; Cagetti, E.; Pisu, M.G.; Porta, S.; Floris, S.; Massa, F.; Sanna, E.; Biggio, G. Allopregnanolone synthesis in cerebellar granule cells: Roles in regulation of GABA(A) receptor expression and function during progesterone treatment and withdrawal. Mol. Pharm. 2000, 57, 1262–1270. [Google Scholar]
- Walker, K.A.; Ficek, B.N.; Westbrook, R. Understanding the Role of Systemic Inflammation in Alzheimer’s Disease. ACS Chem. Neurosci. 2019, 10, 3340–3342. [Google Scholar] [CrossRef]
- Rossi, S.; Studer, V.; Motta, C.; De Chiara, V.; Barbieri, F.; Bernardi, G.; Centonze, D. Inflammation inhibits GABA transmission in multiple sclerosis. Mult. Scler. 2012, 18, 1633–1635. [Google Scholar] [CrossRef] [PubMed]
- Guerriero, R.M.; Giza, C.C.; Rotenberg, A. Glutamate and GABA imbalance following traumatic brain injury. Curr. Neurol. Neurosci. Rep. 2015, 15, 27. [Google Scholar] [CrossRef] [PubMed]
- Saitow, F.; Murano, M.; Suzuki, H. Modulatory effects of serotonin on GABAergic synaptic transmission and membrane properties in the deep cerebellar nuclei. J. Neurophysiol. 2009, 101, 1361–1374. [Google Scholar] [CrossRef]
- Rudroff, T.; Workman, C.D.; Fietsam, A.C.; Kamholz, J. Response Variability in Transcranial Direct Current Stimulation: Why Sex Matters. Front. Psychiatry 2020, 11, 585. [Google Scholar] [CrossRef]
- Chinn, S.; Caldwell, W.; Gritsenko, K. Fibromyalgia Pathogenesis and Treatment Options Update. Curr. Pain Headache Rep. 2016, 20, 25. [Google Scholar] [CrossRef]
- Kim, S.C.; Landon, J.E.; Solomon, D.H. Clinical characteristics and medication uses among fibromyalgia patients newly prescribed amitriptyline, duloxetine, gabapentin, or pregabalin. Arthritis Care Res. 2013, 65, 1813–1819. [Google Scholar] [CrossRef] [PubMed]
- Fitelson, E.; Kim, S.; Baker, A.S.; Leight, K. Treatment of postpartum depression: Clinical, psychological and pharmacological options. Int. J. Womens Health 2010, 3, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Gentile, S.; Fusco, M.L. Managing fibromyalgia syndrome in pregnancy no bridges between USA and EU. Arch. Womens Ment. Health 2019, 22, 711–721. [Google Scholar] [CrossRef] [PubMed]
- Abbott, L.F.; Nelson, S.B. Synaptic plasticity: Taming the beast. Nat. Neurosci. 2000, 3, 1178–1183. [Google Scholar] [CrossRef] [PubMed]
- Carcea, I.; Froemke, R.C. Cortical plasticity, excitatory-inhibitory balance, and sensory perception. Prog. Brain Res. 2013, 207, 65–90. [Google Scholar] [CrossRef]
- Hendry, S.H.; Schwark, H.D.; Jones, E.G.; Yan, J. Numbers and proportions of GABA-immunoreactive neurons in different areas of monkey cerebral cortex. J. Neurosci. 1987, 7, 1503–1519. [Google Scholar] [CrossRef]
- Wang, M. Neurosteroids and GABA-A Receptor Function. Front. Endocrinol. 2011, 2, 44. [Google Scholar] [CrossRef]
- Chaieb, L.; Antal, A.; Paulus, W. Gender-specific modulation of short-term neuroplasticity in the visual cortex induced by transcranial direct current stimulation. Vis. Neurosci. 2008, 25, 77–81. [Google Scholar] [CrossRef]
- Dedoncker, J.; Brunoni, A.R.; Baeken, C.; Vanderhasselt, M.A. A Systematic Review and Meta-Analysis of the Effects of Transcranial Direct Current Stimulation (tDCS) Over the Dorsolateral Prefrontal Cortex in Healthy and Neuropsychiatric Samples: Influence of Stimulation Parameters. Brain Stimul. 2016, 9, 501–517. [Google Scholar] [CrossRef]
- Vigod, S.N.; Murphy, K.E.; Dennis, C.L.; Oberlander, T.F.; Ray, J.G.; Daskalakis, Z.J.; Blumberger, D.M. Transcranial direct current stimulation (tDCS) for depression in pregnancy: A pilot randomized controlled trial. Brain Stimul. 2019, 12, 1475–1483. [Google Scholar] [CrossRef]
- Raimundo, R.J.S.; Uribe, C.E.; Brasil-Neto, J.P. Lack of clinically detectable acute changes on autonomic or thermoregulatory functions in healthy subjects after transcranial direct current stimulation (tDCS). Brain Stimul. 2012, 5, 196–200. [Google Scholar] [CrossRef] [PubMed]
- Fregni, F.; Gimenes, R.; Valle, A.C.; Ferreira, M.J.; Rocha, R.R.; Natalle, L.; Bravo, R.; Rigonatti, S.P.; Freedman, S.D.; Nitsche, M.A.; et al. A randomized, sham-controlled, proof of principle study of transcranial direct current stimulation for the treatment of pain in fibromyalgia. Arthritis Rheum. 2006, 54, 3988–3998. [Google Scholar] [CrossRef] [PubMed]
- Fregni, F.; El-Hagrassy, M.M.; Pacheco-Barrios, K.; Carvalho, S.; Leite, J.; Simis, M.; Brunelin, J.; Nakamura-Palacios, E.M.; Marangolo, P.; Venkatasubramanian, G.; et al. Evidence-Based Guidelines and Secondary Meta-Analysis for the Use of Transcranial Direct Current Stimulation in Neurological and Psychiatric Disorders. Int. J. Neuropsychopharmacol. 2021, 24, 256–313. [Google Scholar] [CrossRef] [PubMed]
- Palm, U.; Kumpf, U.; Behler, N.; Wulf, L.; Kirsch, B.; Worsching, J.; Keeser, D.; Hasan, A.; Padberg, F. Home Use, Remotely Supervised, and Remotely Controlled Transcranial Direct Current Stimulation: A Systematic Review of the Available Evidence. Neuromodulation 2018, 21, 323–333. [Google Scholar] [CrossRef]
- Merzagora, A.C.; Foffani, G.; Panyavin, I.; Mordillo-Mateos, L.; Aguilar, J.; Onaral, B.; Oliviero, A. Prefrontal hemodynamic changes produced by anodal direct current stimulation. Neuroimage 2010, 49, 2304–2310. [Google Scholar] [CrossRef]
- Kurzeck, A.K.; Dechantsreiter, E.; Wilkening, A.; Kumpf, U.; Nenov-Matt, T.; Padberg, F.; Palm, U. Transcranial Direct Current Stimulation (tDCS) for Depression during Pregnancy: Results from an Open-Label Pilot Study. Brain Sci. 2021, 11, 947. [Google Scholar] [CrossRef]
- Konstantinou, P.; Kassianos, A.P.; Georgiou, G.; Panayides, A.; Papageorgiou, A.; Almas, I.; Wozniak, G.; Karekla, M. Barriers, facilitators, and interventions for medication adherence across chronic conditions with the highest non-adherence rates: A scoping review with recommendations for intervention development. Transl. Behav. Med. 2020, 10, 1390–1398. [Google Scholar] [CrossRef]
- Kurzeck, A.K.; Kirsch, B.; Weidinger, E.; Padberg, F.; Palm, U. Transcranial Direct Current Stimulation (tDCS) for Depression during Pregnancy: Scientific Evidence and What Is Being Said in the Media-A Systematic Review. Brain Sci. 2018, 8, 155. [Google Scholar] [CrossRef]
- Braz Ade, S.; de Paula, A.P.; Diniz Mde, F.; de Almeida, R.N. Non-pharmacological therapy and complementary and alternative medicine in fibromyalgia. Rev. Bras. Reumatol. 2011, 51, 269–282. [Google Scholar]
- Petzinger, G.M.; Fisher, B.E.; McEwen, S.; Beeler, J.A.; Walsh, J.P.; Jakowec, M.W. Exercise-enhanced neuroplasticity targeting motor and cognitive circuitry in Parkinson’s disease. Lancet Neurol. 2013, 12, 716–726. [Google Scholar] [CrossRef]
- Shenoy, S.; Bose, A.; Chhabra, H.; Dinakaran, D.; Agarwal, S.M.; Shivakumar, V.; Narayanaswamy, J.C.; Sivakumar, P.T.; Venkatasubramanian, G. Transcranial direct current stimulation (tDCS) for auditory verbal hallucinations in schizophrenia during pregnancy: A case report. Brain Stimul. 2015, 8, 163–164. [Google Scholar] [CrossRef] [PubMed]
- Sreeraj, V.S.; Bose, A.; Shanbhag, V.; Narayanaswamy, J.C.; Venkatasubramanian, G.; Benegal, V. Monotherapy With tDCS for Treatment of Depressive Episode During Pregnancy: A Case Report. Brain Stimul. 2016, 9, 457–458. [Google Scholar] [CrossRef] [PubMed]
- Strube, W.; Kirsch, B.; Padberg, F.; Hasan, A.; Palm, U. Transcranial Direct Current Stimulation as Monotherapy for the Treatment of Auditory Hallucinations During Pregnancy: A Case Report. J. Clin. Psychopharmacol. 2016, 36, 534–535. [Google Scholar] [CrossRef] [PubMed]
- Palm, U.; Kirsch, B.; Leitner, B.; Popovic, D.; Padberg, F. P017 Transcranial direct current stimulation (tDCS) for the treatment of depression during pregnancy: A pilot study. Clin. Neurophysiol. 2017, 128, e17–e18. [Google Scholar] [CrossRef]
- Donahue, M.J.; Near, J.; Blicher, J.U.; Jezzard, P. Baseline GABA concentration and fMRI response. Neuroimage 2010, 53, 392–398. [Google Scholar] [CrossRef]
- Kiemes, A.; Davies, C.; Kempton, M.J.; Lukow, P.B.; Bennallick, C.; Stone, J.M.; Modinos, G. GABA, Glutamate and Neural Activity: A Systematic Review With Meta-Analysis of Multimodal (1)H-MRS-fMRI Studies. Front. Psychiatry 2021, 12, 644315. [Google Scholar] [CrossRef]

| Pregnancy Trimesters | ||||
|---|---|---|---|---|
| Hormone | Menstrual Cycle (min and max) | First | Second | Third |
| Progesterone (ng/mL) | 2–25 | 8–48 | 32–80 | 99–342 |
| 17β-estradiol (pg/mL) | 30–400 | 188–2497 | 1278–7192 | 6137–3460 |
| Prolactin (ng/mL) | < 20 | 36–213 | 110–330 | 137–372 |
| Cortisol (µg/dL) | 10–20 (CAR) | 7–19 | 10–42 | 12–50 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mucci, V.; Demori, I.; Browne, C.J.; Deblieck, C.; Burlando, B. Fibromyalgia in Pregnancy: Neuro-Endocrine Fluctuations Provide Insight into Pathophysiology and Neuromodulation Treatment. Biomedicines 2023, 11, 615. https://doi.org/10.3390/biomedicines11020615
Mucci V, Demori I, Browne CJ, Deblieck C, Burlando B. Fibromyalgia in Pregnancy: Neuro-Endocrine Fluctuations Provide Insight into Pathophysiology and Neuromodulation Treatment. Biomedicines. 2023; 11(2):615. https://doi.org/10.3390/biomedicines11020615
Chicago/Turabian StyleMucci, Viviana, Ilaria Demori, Cherylea J. Browne, Choi Deblieck, and Bruno Burlando. 2023. "Fibromyalgia in Pregnancy: Neuro-Endocrine Fluctuations Provide Insight into Pathophysiology and Neuromodulation Treatment" Biomedicines 11, no. 2: 615. https://doi.org/10.3390/biomedicines11020615
APA StyleMucci, V., Demori, I., Browne, C. J., Deblieck, C., & Burlando, B. (2023). Fibromyalgia in Pregnancy: Neuro-Endocrine Fluctuations Provide Insight into Pathophysiology and Neuromodulation Treatment. Biomedicines, 11(2), 615. https://doi.org/10.3390/biomedicines11020615