

Seeking the Amygdala: Novel Use of Diffusion Tensor Imaging to Delineate the Basolateral Amygdala
 , , , , and
, , , , and {kind=link}
{kind=link}
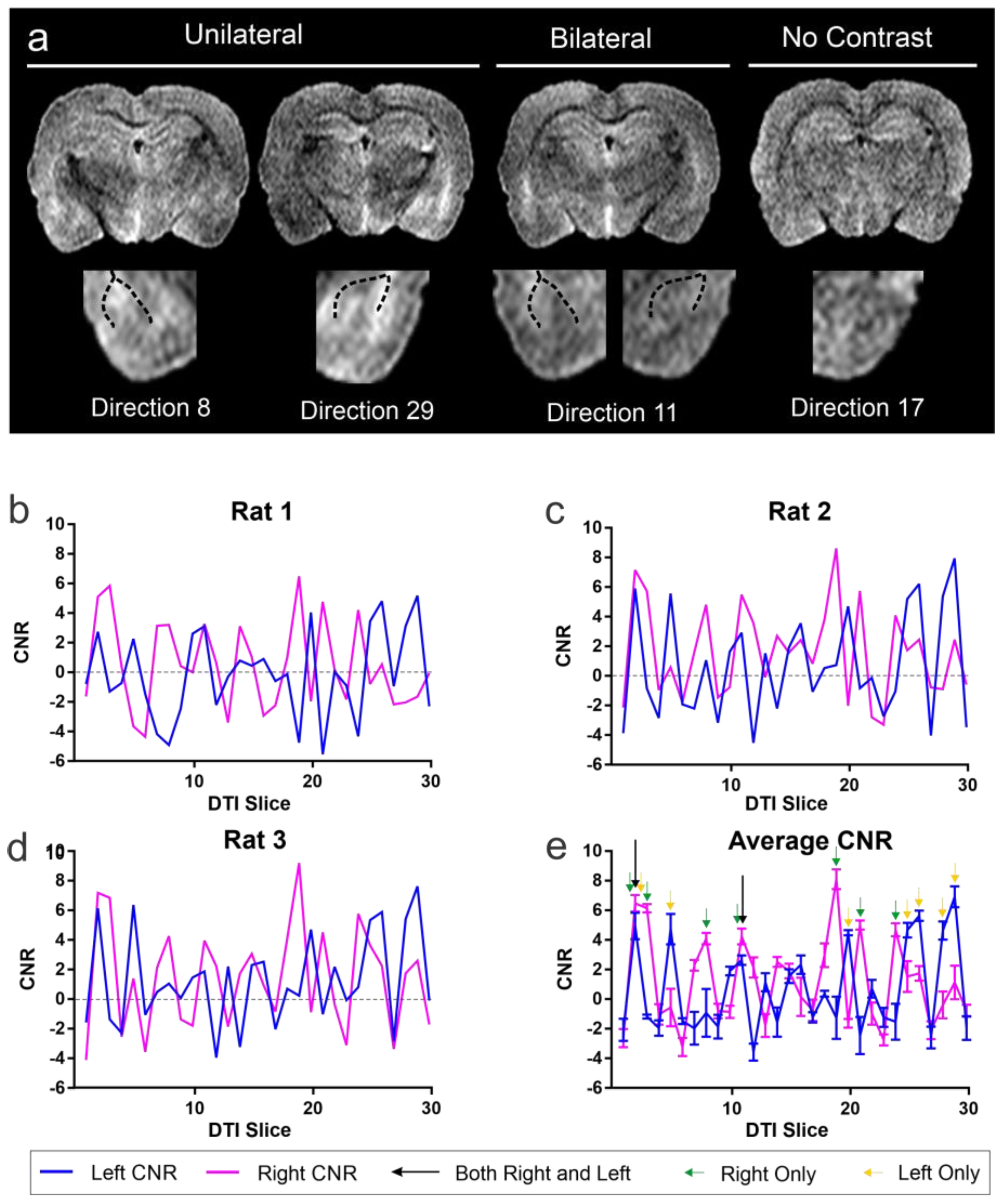{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. T1- and T2-Weighted Magnetic Resonance Imaging (MRI)
2.3. Diffusion Tensor Imaging (DTI)
2.4. BLA Volumetric Analysis from DTI
2.5. Histology and Immunohistochemistry
2.6. Statistical Analysis
3. Results
3.1. Neuroimaging and Contrast Enhancement of the Rodent Amygdala
3.2. Exogenous Contrast Does Not Enhance Definition of the BLA
3.3. High Resolution Neuroimaging and Conventional DTI Do Not Enhance BLA Definition
3.4. Specific Directionally Encoded DTI Directions Enhance BLA Visualization
3.5. Use of DTI and Slice-Shifting for Accurate Measures of BLA Volume
3.6. Histological Comparison and Validation of MRI-Derived BLA Volumes
3.7. Additional Validation of MRI-Derived BLA Regions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Chareyron, L.J.; Banta Lavenex, P.; Amaral, D.G.; Lavenex, P. Stereological analysis of the rat and monkey amygdala. J. Comp. Neurol. 2011, 519, 3218–3239. [Google Scholar] [CrossRef]
- LeDoux, J. The amygdala. Curr. Biol. 2007, 17, R868–R874. [Google Scholar] [CrossRef]
- Pitkanen, A.; Savander, V.; LeDoux, J.E. Organization of intra-amygdaloid circuitries in the rat: An emerging framework for understanding functions of the amygdala. Trends Neurosci. 1997, 20, 517–523. [Google Scholar] [CrossRef]
- Hakamata, Y.; Mizukami, S.; Izawa, S.; Okamura, H.; Mihara, K.; Marusak, H.; Moriguchi, Y.; Hori, H.; Hanakawa, T.; Inoue, Y.; et al. Implicit and explicit emotional memory recall in anxiety and depression: Role of basolateral amygdala and cortisol-norepinephrine interaction. Psychoneuroendocrinology 2022, 136, 105598. [Google Scholar] [CrossRef]
- Klune, C.B.; Jin, B.; DeNardo, L.A. Linking mPFC circuit maturation to the developmental regulation of emotional memory and cognitive flexibility. eLife 2021, 10, e64567. [Google Scholar] [CrossRef]
- Murray, R.J.; Brosch, T.; Sander, D. The functional profile of the human amygdala in affective processing: Insights from intracranial recordings. Cortex J. Devoted Study Nerv. Syst. Behav. 2014, 60, 10–33. [Google Scholar] [CrossRef]
- Duvarci, S.; Pare, D. Amygdala microcircuits controlling learned fear. Neuron 2014, 82, 966–980. [Google Scholar] [CrossRef]
- Roozendaal, B.; Brunson, K.L.; Holloway, B.L.; McGaugh, J.L.; Baram, T.Z. Involvement of stress-released corticotropin-releasing hormone in the basolateral amygdala in regulating memory consolidation. Proc. Natl. Acad. Sci. USA 2002, 99, 13908–13913. [Google Scholar] [CrossRef]
- Drevets, W.C. Functional anatomical abnormalities in limbic and prefrontal cortical structures in major depression. Prog. Brain Res. 2000, 126, 413–431. [Google Scholar] [CrossRef]
- McEwen, B.S. Plasticity of the hippocampus: Adaptation to chronic stress and allostatic load. Ann. N. Y. Acad. Sci. 2001, 933, 265–277. [Google Scholar] [CrossRef]
- Birnie, M.T.; Kooiker, C.L.; Short, A.K.; Bolton, J.L.; Chen, Y.; Baram, T.Z. Plasticity of the Reward Circuitry After Early-Life Adversity: Mechanisms and Significance. Biol. Psychiatry 2020, 87, 875–884. [Google Scholar] [CrossRef] [PubMed]
- Bolton, J.L.; Molet, J.; Regev, L.; Chen, Y.; Rismanchi, N.; Haddad, E.; Yang, D.Z.; Obenaus, A.; Baram, T.Z. Anhedonia Following Early-Life Adversity Involves Aberrant Interaction of Reward and Anxiety Circuits and Is Reversed by Partial Silencing of Amygdala Corticotropin-Releasing Hormone Gene. Biol. Psychiatry 2018, 83, 137–147. [Google Scholar] [CrossRef]
- Kemppainen, S.; Pitkanen, A. Distribution of parvalbumin, calretinin, and calbindin-D(28k) immunoreactivity in the rat amygdaloid complex and colocalization with gamma-aminobutyric acid. J. Comp. Neurol. 2000, 426, 441–467. [Google Scholar] [CrossRef]
- Majak, K.; Pitkanen, A. Projections from the periamygdaloid cortex to the amygdaloid complex, the hippocampal formation, and the parahippocampal region: A PHA-L study in the rat. Hippocampus 2003, 13, 922–942. [Google Scholar] [CrossRef]
- Pitkanen, A.; Pikkarainen, M.; Nurminen, N.; Ylinen, A. Reciprocal connections between the amygdala and the hippocampal formation, perirhinal cortex, and postrhinal cortex in rat. A review. Ann. N. Y. Acad. Sci. 2000, 911, 369–391. [Google Scholar] [CrossRef]
- Shammah-Lagnado, S.J.; Alheid, G.F.; Heimer, L. Striatal and central extended amygdala parts of the interstitial nucleus of the posterior limb of the anterior commissure: Evidence from tract-tracing techniques in the rat. J. Comp. Neurol. 2001, 439, 104–126. [Google Scholar] [CrossRef]
- Unal, G.; Pare, J.F.; Smith, Y.; Pare, D. Cortical inputs innervate calbindin-immunoreactive interneurons of the rat basolateral amygdaloid complex. J. Comp. Neurol. 2014, 522, 1915–1928. [Google Scholar] [CrossRef] [PubMed]
- Hintiryan, H.; Bowman, I.; Johnson, D.L.; Korobkova, L.; Zhu, M.; Khanjani, N.; Gou, L.; Gao, L.; Yamashita, S.; Bienkowski, M.S.; et al. Connectivity characterization of the mouse basolateral amygdalar complex. Nat. Commun. 2021, 12, 2859. [Google Scholar] [CrossRef]
- Goncalves Pereira, P.M.; Insausti, R.; Artacho-Perula, E.; Salmenpera, T.; Kalviainen, R.; Pitkanen, A. MR volumetric analysis of the piriform cortex and cortical amygdala in drug-refractory temporal lobe epilepsy. Am. J. Neuroradiol. 2005, 26, 319–332. [Google Scholar]
- Lupton, M.K.; Strike, L.; Hansell, N.K.; Wen, W.; Mather, K.A.; Armstrong, N.J.; Thalamuthu, A.; McMahon, K.L.; de Zubicaray, G.I.; Assareh, A.A.; et al. The effect of increased genetic risk for Alzheimer’s disease on hippocampal and amygdala volume. Neurobiol. Aging 2016, 40, 68–77. [Google Scholar] [CrossRef]
- O’Doherty, D.C.; Chitty, K.M.; Saddiqui, S.; Bennett, M.R.; Lagopoulos, J. A systematic review and meta-analysis of magnetic resonance imaging measurement of structural volumes in posttraumatic stress disorder. Psychiatry Res. 2015, 232, 1–33. [Google Scholar] [CrossRef]
- Manelis, A.; Ladouceur, C.D.; Graur, S.; Monk, K.; Bonar, L.K.; Hickey, M.B.; Dwojak, A.C.; Axelson, D.; Goldstein, B.I.; Goldstein, T.R.; et al. Altered amygdala-prefrontal response to facial emotion in offspring of parents with bipolar disorder. Brain J. Neurol. 2015, 138, 2777–2790. [Google Scholar] [CrossRef]
- Tajima-Pozo, K.; Yus, M.; Ruiz-Manrique, G.; Lewczuk, A.; Arrazola, J.; Montanes-Rada, F. Amygdala Abnormalities in Adults With ADHD. J. Atten. Disord. 2016, 22, 671–678. [Google Scholar] [CrossRef]
- Delgado y Palacios, R.; Verhoye, M.; Henningsen, K.; Wiborg, O.; Van der Linden, A. Diffusion kurtosis imaging and high-resolution MRI demonstrate structural aberrations of caudate putamen and amygdala after chronic mild stress. PLoS ONE 2014, 9, e95077. [Google Scholar] [CrossRef]
- Saygin, Z.M.; Osher, D.E.; Augustinack, J.; Fischl, B.; Gabrieli, J.D. Connectivity-based segmentation of human amygdala nuclei using probabilistic tractography. NeuroImage 2011, 56, 1353–1361. [Google Scholar] [CrossRef]
- Solano-Castiella, E.; Anwander, A.; Lohmann, G.; Weiss, M.; Docherty, C.; Geyer, S.; Reimer, E.; Friederici, A.D.; Turner, R. Diffusion tensor imaging segments the human amygdala in vivo. NeuroImage 2010, 49, 2958–2965. [Google Scholar] [CrossRef]
- Aoki, I.; Wu, Y.J.; Silva, A.C.; Lynch, R.M.; Koretsky, A.P. In vivo detection of neuroarchitecture in the rodent brain using manganese-enhanced MRI. NeuroImage 2004, 22, 1046–1059. [Google Scholar] [CrossRef]
- Bangasser, D.A.; Lee, C.S.; Cook, P.A.; Gee, J.C.; Bhatnagar, S.; Valentino, R.J. Manganese-enhanced magnetic resonance imaging (MEMRI) reveals brain circuitry involved in responding to an acute novel stress in rats with a history of repeated social stress. Physiol. Behav. 2013, 122, 228–236. [Google Scholar] [CrossRef]
- Bouilleret, V.; Cardamone, L.; Liu, Y.R.; Fang, K.; Myers, D.E.; O’Brien, T.J. Progressive brain changes on serial manganese-enhanced MRI following traumatic brain injury in the rat. J. Neurotrauma 2009, 26, 1999–2013. [Google Scholar] [CrossRef]
- Obenaus, A.; Jacobs, R.E. Magnetic resonance imaging of functional anatomy: Use for small animal epilepsy models. Epilepsia 2007, 48 (Suppl. S4), 11–17. [Google Scholar] [CrossRef]
- Soares, J.M.; Marques, P.; Alves, V.; Sousa, N. A hitchhiker’s guide to diffusion tensor imaging. Front. Neurosci. 2013, 7, 31. [Google Scholar] [CrossRef]
- Wendel, K.M.; Short, A.K.; Noarbe, B.P.; Haddad, E.; Palma, A.M.; Yassa, M.A.; Baram, T.Z.; Obenaus, A. Early life adversity in male mice sculpts reward circuits. Neurobiol. Stress 2021, 15, 100409. [Google Scholar] [CrossRef]
- Bracht, T.; Linden, D.; Keedwell, P. A review of white matter microstructure alterations of pathways of the reward circuit in depression. J. Affect. Disord. 2015, 187, 45–53. [Google Scholar] [CrossRef]
- Wu, E.X.; Cheung, M.M. MR diffusion kurtosis imaging for neural tissue characterization. NMR Biomed. 2010, 23, 836–848. [Google Scholar] [CrossRef]
- Papp, E.A.; Leergaard, T.B.; Calabrese, E.; Johnson, G.A.; Bjaalie, J.G. Waxholm Space atlas of the Sprague Dawley rat brain. NeuroImage 2014, 97, 374–386. [Google Scholar] [CrossRef]
- Calabrese, E.; Johnson, G.A. Diffusion tensor magnetic resonance histology reveals microstructural changes in the developing rat brain. NeuroImage 2013, 79, 329–339. [Google Scholar] [CrossRef]
- Haacke, E.M.; Cheng, N.Y.; House, M.J.; Liu, Q.; Neelavalli, J.; Ogg, R.J.; Khan, A.; Ayaz, M.; Kirsch, W.; Obenaus, A. Imaging iron stores in the brain using magnetic resonance imaging. Magn. Reson. Imaging 2005, 23, 1–25. [Google Scholar] [CrossRef]
- McEwen, B.S.; Chattarji, S. Molecular mechanisms of neuroplasticity and pharmacological implications: The example of tianeptine. Eur. Neuropsychopharmacol. 2004, 14 (Suppl. S5), S497–S502. [Google Scholar] [CrossRef]
- Kassem, M.S.; Lagopoulos, J.; Stait-Gardner, T.; Price, W.S.; Chohan, T.W.; Arnold, J.C.; Hatton, S.N.; Bennett, M.R. Stress-induced grey matter loss determined by MRI is primarily due to loss of dendrites and their synapses. Mol. Neurobiol. 2013, 47, 645–661. [Google Scholar] [CrossRef]
- Spinelli, S.; Muller, T.; Friedel, M.; Sigrist, H.; Lesch, K.P.; Henkelman, M.; Rudin, M.; Seifritz, E.; Pryce, C.R. Effects of repeated adolescent stress and serotonin transporter gene partial knockout in mice on behaviors and brain structures relevant to major depression. Front. Behav. Neurosci. 2013, 7, 215. [Google Scholar] [CrossRef]
- Entis, J.J.; Doerga, P.; Barrett, L.F.; Dickerson, B.C. A reliable protocol for the manual segmentation of the human amygdala and its subregions using ultra-high resolution MRI. NeuroImage 2012, 60, 1226–1235. [Google Scholar] [CrossRef]
- Solodkin, A.; Chen, E.E.; Van Hoesen, G.W.; Heimer, L.; Shereen, A.; Kruggel, F.; Mastrianni, J. In vivo parahippocampal white matter pathology as a biomarker of disease progression to Alzheimer’s disease. J. Comp. Neurol. 2013, 521, 4300–4317. [Google Scholar] [CrossRef]
- Zalsman, G.; Gutman, A.; Shbiro, L.; Rosenan, R.; Mann, J.J.; Weller, A. Genetic vulnerability, timing of short-term stress and mood regulation: A rodent diffusion tensor imaging study. Eur. Neuropsychopharmacol. 2015, 25, 2075–2085. [Google Scholar] [CrossRef]
- Solano-Castiella, E.; Schafer, A.; Reimer, E.; Turke, E.; Proger, T.; Lohmann, G.; Trampel, R.; Turner, R. Parcellation of human amygdala in vivo using ultra high field structural MRI. NeuroImage 2011, 58, 741–748. [Google Scholar] [CrossRef]
- Bianchi, A.; Bhanu, B.; Obenaus, A. Dynamic low-level context for the detection of mild traumatic brain injury. IEEE Trans. Bio-Med. Eng. 2015, 62, 145–153. [Google Scholar] [CrossRef]
- Donovan, V.; Bianchi, A.; Hartman, R.; Bhanu, B.; Carson, M.J.; Obenaus, A. Computational analysis reveals increased blood deposition following repeated mild traumatic brain injury. NeuroImage. Clin. 2012, 1, 18–28. [Google Scholar] [CrossRef]
- Ghosh, N.; Yuan, X.; Turenius, C.I.; Tone, B.; Ambadipudi, K.; Snyder, E.Y.; Obenaus, A.; Ashwal, S. Automated core-penumbra quantification in neonatal ischemic brain injury. J. Cereb. Blood Flow Metab. 2012, 32, 2161–2170. [Google Scholar] [CrossRef]
- Kruggel, F.; Paul, J.S.; Gertz, H.J. Texture-based segmentation of diffuse lesions of the brain’s white matter. NeuroImage 2008, 39, 987–996. [Google Scholar] [CrossRef]
- Goncalves, L.; Silva, R.; Pinto-Ribeiro, F.; Pego, J.M.; Bessa, J.M.; Pertovaara, A.; Sousa, N.; Almeida, A. Neuropathic pain is associated with depressive behaviour and induces neuroplasticity in the amygdala of the rat. Exp. Neurol. 2008, 213, 48–56. [Google Scholar] [CrossRef]
- Pego, J.M.; Morgado, P.; Pinto, L.G.; Cerqueira, J.J.; Almeida, O.F.; Sousa, N. Dissociation of the morphological correlates of stress-induced anxiety and fear. Eur. J. Neurosci. 2008, 27, 1503–1516. [Google Scholar] [CrossRef]
- Daviu, N.; Bruchas, M.R.; Moghaddam, B.; Sandi, C.; Beyeler, A. Neurobiological links between stress and anxiety. Neurobiol. Stress 2019, 11, 100191. [Google Scholar] [CrossRef]
- Crowley, R.; Bendor, D.; Javadi, A.H. A review of neurobiological factors underlying the selective enhancement of memory at encoding, consolidation, and retrieval. Prog. Neurobiol. 2019, 179, 101615. [Google Scholar] [CrossRef]
- Zhang, L.; Hu, X.; Lu, L.; Li, B.; Hu, X.; Bu, X.; Li, H.; Tang, S.; Gao, Y.; Yang, Y.; et al. Anatomic alterations across amygdala subnuclei in medication-free patients with obsessive-compulsive disorder. J. Psychiatry Neurosci. 2020, 45, 190114. [Google Scholar] [CrossRef]
- Morey, R.A.; Clarke, E.K.; Haswell, C.C.; Phillips, R.D.; Clausen, A.N.; Mufford, M.S.; Saygin, Z.; Workgroup, V.A.M.-A.M.; Wagner, H.R.; LaBar, K.S. Amygdala Nuclei Volume and Shape in Military Veterans With Posttraumatic Stress Disorder. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2020, 5, 281–290. [Google Scholar] [CrossRef]
- Brown, S.S.G.; Rutland, J.W.; Verma, G.; Feldman, R.E.; Alper, J.; Schneider, M.; Delman, B.N.; Murrough, J.M.; Balchandani, P. Structural MRI at 7T reveals amygdala nuclei and hippocampal subfield volumetric association with Major Depressive Disorder symptom severity. Sci. Rep. 2019, 9, 10166. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Obenaus, A.; Kinney-Lang, E.; Jullienne, A.; Haddad, E.; Wendel, K.M.; Shereen, A.D.; Solodkin, A.; Dunn, J.F.; Baram, T.Z. Seeking the Amygdala: Novel Use of Diffusion Tensor Imaging to Delineate the Basolateral Amygdala. Biomedicines 2023, 11, 535. https://doi.org/10.3390/biomedicines11020535
Obenaus A, Kinney-Lang E, Jullienne A, Haddad E, Wendel KM, Shereen AD, Solodkin A, Dunn JF, Baram TZ. Seeking the Amygdala: Novel Use of Diffusion Tensor Imaging to Delineate the Basolateral Amygdala. Biomedicines. 2023; 11(2):535. https://doi.org/10.3390/biomedicines11020535
Chicago/Turabian StyleObenaus, Andre, Eli Kinney-Lang, Amandine Jullienne, Elizabeth Haddad, Kara M. Wendel, A. Duke Shereen, Ana Solodkin, Jeffrey F. Dunn, and Tallie Z. Baram. 2023. "Seeking the Amygdala: Novel Use of Diffusion Tensor Imaging to Delineate the Basolateral Amygdala" Biomedicines 11, no. 2: 535. https://doi.org/10.3390/biomedicines11020535
APA StyleObenaus, A., Kinney-Lang, E., Jullienne, A., Haddad, E., Wendel, K. M., Shereen, A. D., Solodkin, A., Dunn, J. F., & Baram, T. Z. (2023). Seeking the Amygdala: Novel Use of Diffusion Tensor Imaging to Delineate the Basolateral Amygdala. Biomedicines, 11(2), 535. https://doi.org/10.3390/biomedicines11020535