
Conversion of M1 Macrophages to Foam Cells: Transcriptome Differences Determined by Sex

 , , ,
, , ,  , , ,
, , ,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Low-Density Lipoprotein (LDL) Isolation
2.3. In Vitro Isolation and Cultivation of Monocytes
2.4. Macrophage Differentiation and LDL Treatment
2.5. Microarray Expression and Analysis
2.6. Ingenuity Pathway Analysis (IPA)
2.7. Statistical Analyses
3. Results
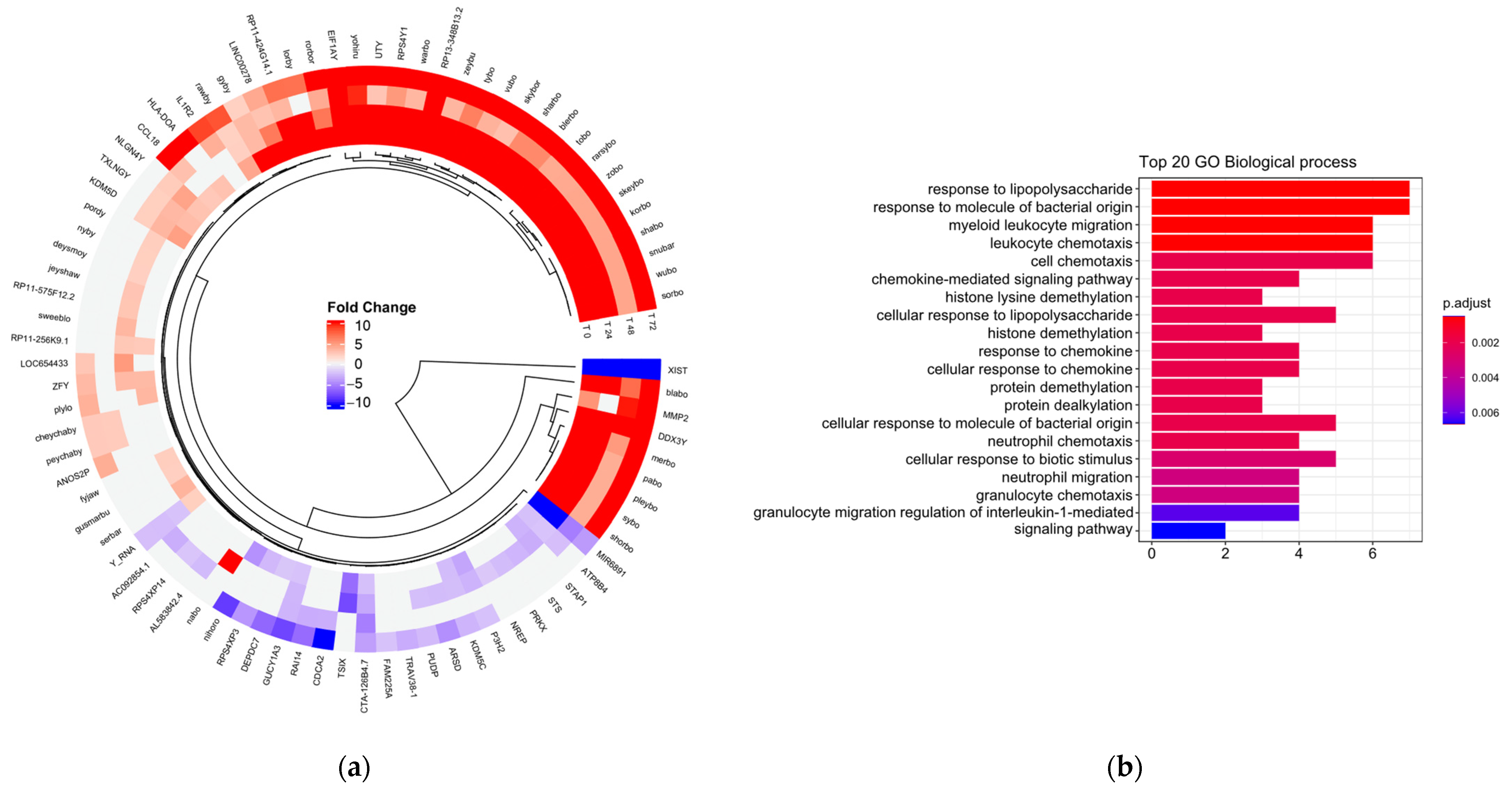
3.1. Transcriptomic Profile of M1 Macrophages Stimulated with LDL Associated with Sex
3.2. Pathways in M1 Macrophages Stimulated with LDL Associated with Sex and Time
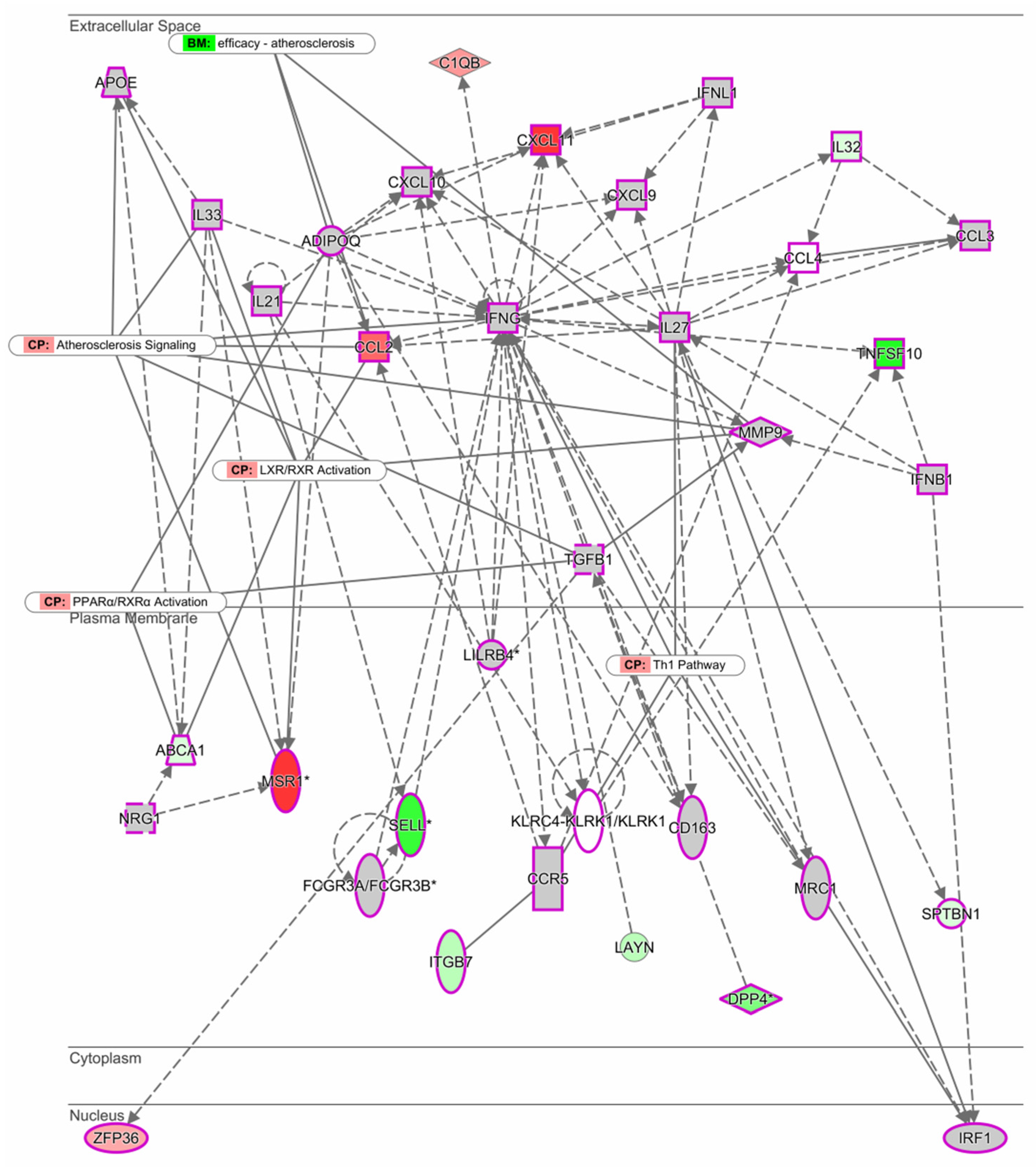
3.3. Gene Network between Men and Women in M1 Macrophages Stimulated with LDL
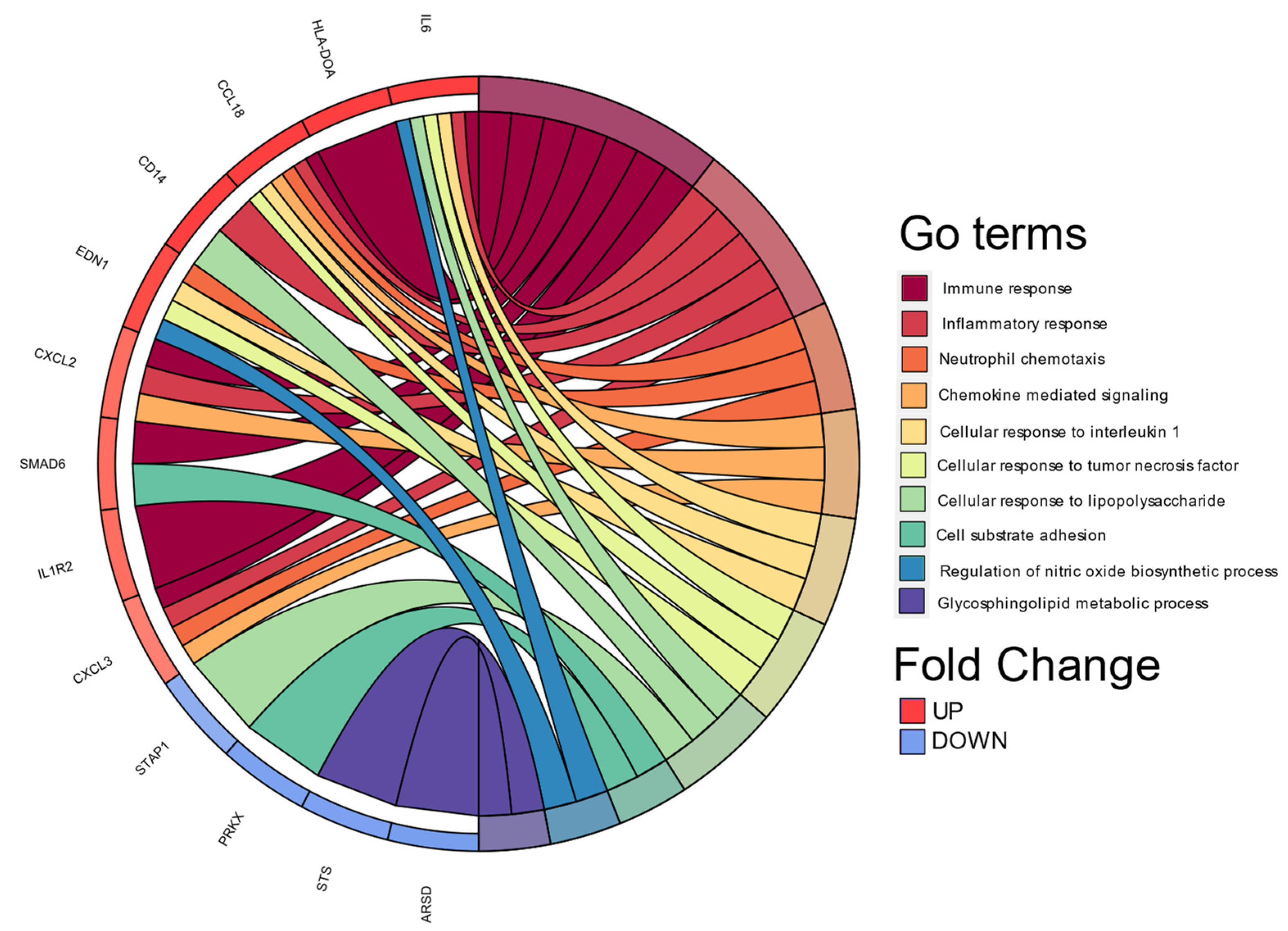
3.4. Biological Processes Associated with Cellular Functions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- INEGI—INdEyG. Características de las Defunciones Registradas en México Durante 2017; Instituto Nacional de Estadística y Geografía: Mexico City, Mexico, 2017.
- Tabas, I.; Bornfeldt, K.E. Macrophage Phenotype and Function in Different Stages of Atherosclerosis. Circ. Res. 2016, 118, 653–667. [Google Scholar] [CrossRef] [PubMed]
- Zimmer, S.; Grebe, A.; Latz, E. Danger signaling in atherosclerosis. Circ. Res. 2015, 116, 323–340. [Google Scholar] [PubMed]
- Groh, L.; Keating, S.T.; Joosten, L.A.B.; Netea, M.G.; Riksen, N.P. Monocyte and macrophage immunometabolism in atherosclerosis. Semin. Immunopathol. 2018, 40, 203–214. [Google Scholar] [PubMed]
- Fairweather, D. Sex Differences in Inflammation during Atherosclerosis. Clin. Med. Insights Cardiol. 2014, 8, 49–59. [Google Scholar]
- Yuan, X.M.; Ward, L.J.; Forssell, C.; Siraj, N.; Li, W. Carotid Atheroma from Men Has Significantly Higher Levels of Inflammation and Iron Metabolism Enabled by Macrophages. Stroke 2018, 49, 419–425. [Google Scholar] [CrossRef]
- Klar, A.S.; Michalak-Micka, K.; Biedermann, T.; Simmen-Meuli, C.; Reichmann, E.; Meuli, M. Characterization of M1 and M2 polarization of macrophages in vascularized human dermo-epidermal skin substitutes in vivo. Pediatr. Surg. Int. 2018, 34, 129–135. [Google Scholar] [CrossRef]
- Ueland, T.; Laugsand, L.E.; Vatten, L.J.; Janszky, I.; Platou, C.; Michelsen, A.E.; Damås, J.K.; Aukrust, P.; Åsvold, B.O. Monocyte/macrophage and T cell activation markers are not independently associated with MI risk in healthy individuals––Results from the HUNT Study. Int. J. Cardiol. 2017, 243, 502–504. [Google Scholar]
- Gistera, A.; Hansson, G.K. The immunology of atherosclerosis. Nat. Rev. Nephrol. 2017, 13, 368–380. [Google Scholar]
- De Paoli, F.; Staels, B.; Chinetti-Gbaguidi, G. Macrophage phenotypes and their modulation in atherosclerosis. Circ. J. 2014, 78, 1775–1781. [Google Scholar] [PubMed]
- Tabas, I.; Lichtman, A.H. Monocyte-Macrophages and T Cells in Atherosclerosis. Immunity 2017, 47, 621–634. [Google Scholar]
- Lahn, B.T.; Page, D.C. Functional coherence of the human Y chromosome. Science 1997, 278, 675–680. [Google Scholar]
- Gershoni, M.; Pietrokovski, S. Reduced selection and accumulation of deleterious mutations in genes exclusively expressed in men. Nat. Commun. 2014, 5, 4438. [Google Scholar] [PubMed]
- Su, A.I.; Wiltshire, T.; Batalov, S.; Lapp, H.; Ching, K.A.; Block, D.; Zhang, J.; Soden, R.; Hayakawa, M.; Kreiman, G.; et al. A gene atlas of the mouse and human protein-encoding transcriptomes. Proc. Natl. Acad. Sci. USA 2004, 101, 6062–6067. [Google Scholar]
- Khodabandehlou, K.; Masehi-Lano, J.J.; Poon, C.; Wang, J.; Chung, E.J. Targeting cell adhesion molecules with nanoparticles using in vivo and flow-based in vitro models of atherosclerosis. Exp. Biol. Med. 2017, 242, 799–812. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Jiang, Q.; Ding, Z.; Liu, G.; Yu, P.; Jiang, G.; Yu, Z.; Yang, C.; Qian, J.; Jiang, H.; et al. Identification of a Common Different Gene Expression Signature in Ischemic Cardiomyopathy. Genes 2018, 9, 56. [Google Scholar] [PubMed]
- Case, L.K.; Wall, E.H.; Dragon, J.A.; Saligrama, N.; Krementsov, D.N.; Moussawi, M.; Zachary, J.F.; Huber, S.A.; Blankenhorn, E.P.; Teuscher, C. The Y chromosome as a regulatory element shaping immune cell transcriptomes and susceptibility to autoimmune disease. Genome Res. 2013, 23, 1474–1485. [Google Scholar]
- Bertone-Johnson, E.R.; Virtanen, J.K.; Nurmi, T.; Niskanen, L.; Mursu, J.; Voutilainen, S.; Ronkainen, K.; Kauhanen, J.; Tuomainen, T.P. Follicle-Stimulating Hormone Levels and Subclinical Atherosclerosis in Older Postmenopausal Women. Am. J. Epidemiol. 2018, 187, 16–26. [Google Scholar] [CrossRef]
- Bloomer, L.D.; Nelson, C.P.; Eales, J.; Denniff, M.; Christofidou, P.; Debiec, R.; Moore, J.; Consortium, C.; Zukowska-Szczechowska, E.; Goodall, A.H.; et al. Male-specific region of the Y chromosome and cardiovascular risk: Phylogenetic analysis and gene expression studies. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 1722–1727. [Google Scholar] [CrossRef]
- Miklos, D.B.; Kim, H.T.; Zorn, E.; Hochberg, E.P.; Guo, L.; Mattes-Ritz, A.; Viatte, S.; Soiffer, R.J.; Antin, J.H.; Ritz, J. Antibody response to DBY minor histocompatibility antigen is induced after allogeneic stem cell transplantation and in healthy female donors. Blood 2004, 103, 353–359. [Google Scholar] [CrossRef]
- Mokou, M.; Klein, J.; Makridakis, M.; Bitsika, V.; Bascands, J.L.; Saulnier-Blache, J.S.; Mullen, W.; Sacherer, M.; Zoidakis, J.; Pieske, B.; et al. Proteomics based identification of KDM5 histone demethylases associated with cardiovascular disease. EBioMedicine 2019, 41, 91–104. [Google Scholar] [CrossRef]
- Yao, X.; Yan, C.; Zhang, L.; Li, Y.; Wan, Q. LncRNA ENST00113 promotes proliferation, survival, and migration by activating PI3K/Akt/mTOR signaling pathway in atherosclerosis. Medicine 2018, 97, e0473. [Google Scholar] [CrossRef]
- Huang, S.; Tao, W.; Guo, Z.; Cao, J.; Huang, X. Suppression of long noncoding RNA TTTY15 attenuates hypoxia-induced cardiomyocytes injury by targeting miR-455-5p. Gene 2019, 701, 1–8. [Google Scholar] [CrossRef]
- Xu, X.; Ma, C.; Liu, C.; Duan, Z.; Zhang, L. Knockdown of long noncoding RNA XIST alleviates oxidative low-density lipoprotein-mediated endothelial cells injury through modulation of miR-320/NOD2 axis. Biochem. Biophys. Res. Commun. 2018, 503, 586–592. [Google Scholar] [CrossRef] [PubMed]
- Wan, W.; Murphy, P.M. Regulation of atherogenesis by chemokines and chemokine receptors. Arch. Immunol. Ther. Exp. 2013, 61, 1–14. [Google Scholar]
- Szentes, V.; Gazdag, M.; Szokodi, I.; Dezsi, C.A. The Role of CXCR3 and Associated Chemokines in the Development of Atherosclerosis and During Myocardial Infarction. Front. Immunol. 2018, 9, 1932. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Medley, S.C.; Hu, T.; Hinsdale, M.E.; Lupu, F.; Virmani, R.; Olson, L.E. PDGFRbeta signalling regulates local inflammation and synergizes with hypercholesterolaemia to promote atherosclerosis. Nat. Commun. 2015, 6, 7770. [Google Scholar] [CrossRef]
- Watt, V.; Chamberlain, J.; Steiner, T.; Francis, S.; Crossman, D. TRAIL attenuates the development of atherosclerosis in apolipoprotein E deficient mice. Atherosclerosis 2011, 215, 348–354. [Google Scholar]
- Rozenberg, I.; Sluka, S.H.; Mocharla, P.; Hallenberg, A.; Rotzius, P.; Boren, J.; Kränkel, N.; Landmesser, U.; Borsig, L.; Lüscher, T.F.; et al. Deletion of L-selectin increases atherosclerosis development in ApoE−/− mice. PLoS ONE 2011, 6, e21675. [Google Scholar] [CrossRef] [PubMed]
- Duan, L.; Rao, X.; Xia, C.; Rajagopalan, S.; Zhong, J. The regulatory role of DPP4 in atherosclerotic disease. Cardiovasc. Diabetol. 2017, 16, 76. [Google Scholar] [CrossRef]
- de Winther, M.P.; van Dijk, K.W.; Havekes, L.M.; Hofker, M.H. Macrophage scavenger receptor class A: A multifunctional receptor in atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 290–297. [Google Scholar]
- Zhang, N.; Zhang, Y.; Zhao, S.; Sun, Y. Septin4 as a novel binding partner of PARP1 contributes to oxidative stress induced human umbilical vein endothelial cells injure. Biochem. Biophys. Res. Commun. 2018, 496, 621–627. [Google Scholar] [PubMed]

 ), and other (◯). Graphs were plotted using IPA software by QIAGEN.
), and other (◯). Graphs were plotted using IPA software by QIAGEN.
), and other (◯). Graphs were plotted using IPA software by QIAGEN.
), and other (◯). Graphs were plotted using IPA software by QIAGEN.


{kind=link}
{kind=link}
{kind=link}
| Function | Genes | Fold Change |
|---|---|---|
| Histone demethylase and inductive activity of INF-α | DDX3Y | 254.51 |
| Cellular apoptosis and translation factors | EIF1AY | 108.43 |
| Ribosomal protein translation | RPS4Y1 | 44.91 |
| Unknown | ANOS2P, merbo, RP11-424G14.1, sybo, yohiru, blabo, nabo, tobo, zeybu, blerbo, Serbar, nyby, Shabo, tybo, ZFY-AS1; AC006157.4, pabo, Sharbo, zobo, gyby, pleybo, Shorbo, TC0Y00006490.hg.1, rarsybo, skeybo, vubo, TC0Y00007072.hg.1, korbo, rawby, skybor, warbo, TC0Y00007073.hg.1, rorbor, Snubar, wubo, TC0Y00007286.hg.1, lorby, RP11256K9.1, Sorbo, TC0Y00007293.hg.1, TC0Y00007306.hg.1, Y_RNA. | 38.99 |
| Chromatin organization | UTY, KDM5D, | 26.3 |
| Transcription regulator | UTY, ZFY | 13.26 |
| Long noncoding RNA | TTTY15, LINC00278 | 6.5 |
| Prevention of protein degradation | USP9Y | 4.73 |
| Cellular exocytosis | TXLNGY | 2.92 |
| Adhesion molecules | NLGN4Y (Neuroligin 4 Y-Linked) | 2.24 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nambo-Venegas, R.; Palacios-González, B.; Mas-Oliva, J.; Aurioles-Amozurrutia, A.K.; Cruz-Rangel, A.; Moreno, A.; Hidalgo-Miranda, A.; Rodríguez-Dorantes, M.; Vadillo-Ortega, F.; Xicohtencatl-Cortes, J.; et al. Conversion of M1 Macrophages to Foam Cells: Transcriptome Differences Determined by Sex. Biomedicines 2023, 11, 490. https://doi.org/10.3390/biomedicines11020490
Nambo-Venegas R, Palacios-González B, Mas-Oliva J, Aurioles-Amozurrutia AK, Cruz-Rangel A, Moreno A, Hidalgo-Miranda A, Rodríguez-Dorantes M, Vadillo-Ortega F, Xicohtencatl-Cortes J, et al. Conversion of M1 Macrophages to Foam Cells: Transcriptome Differences Determined by Sex. Biomedicines. 2023; 11(2):490. https://doi.org/10.3390/biomedicines11020490
Chicago/Turabian StyleNambo-Venegas, Rafael, Berenice Palacios-González, Jaime Mas-Oliva, Ana Karen Aurioles-Amozurrutia, Armando Cruz-Rangel, Abel Moreno, Alfredo Hidalgo-Miranda, Mauricio Rodríguez-Dorantes, Felipe Vadillo-Ortega, Juan Xicohtencatl-Cortes, and et al. 2023. "Conversion of M1 Macrophages to Foam Cells: Transcriptome Differences Determined by Sex" Biomedicines 11, no. 2: 490. https://doi.org/10.3390/biomedicines11020490
APA StyleNambo-Venegas, R., Palacios-González, B., Mas-Oliva, J., Aurioles-Amozurrutia, A. K., Cruz-Rangel, A., Moreno, A., Hidalgo-Miranda, A., Rodríguez-Dorantes, M., Vadillo-Ortega, F., Xicohtencatl-Cortes, J., Ruiz-Olmedo, M. I., & Reyes-Grajeda, J. P. (2023). Conversion of M1 Macrophages to Foam Cells: Transcriptome Differences Determined by Sex. Biomedicines, 11(2), 490. https://doi.org/10.3390/biomedicines11020490