
Particle Nanoarchitectonics for Nanomedicine and Nanotherapeutic Drugs with Special Emphasis on Nasal Drugs and Aging

 ,
,  , , , , and
, , , , and 
Abstract

1. Introduction
2. Biosynthesis and Bioavailability of Nanoparticles
3. Nanocarriers and Drug Delivery for CNS
4. Studies in Octogenarians for Genetic Analysis of Aging
4.1. Physiological Aging
4.2. Transcriptional Approach for Studies of Aging
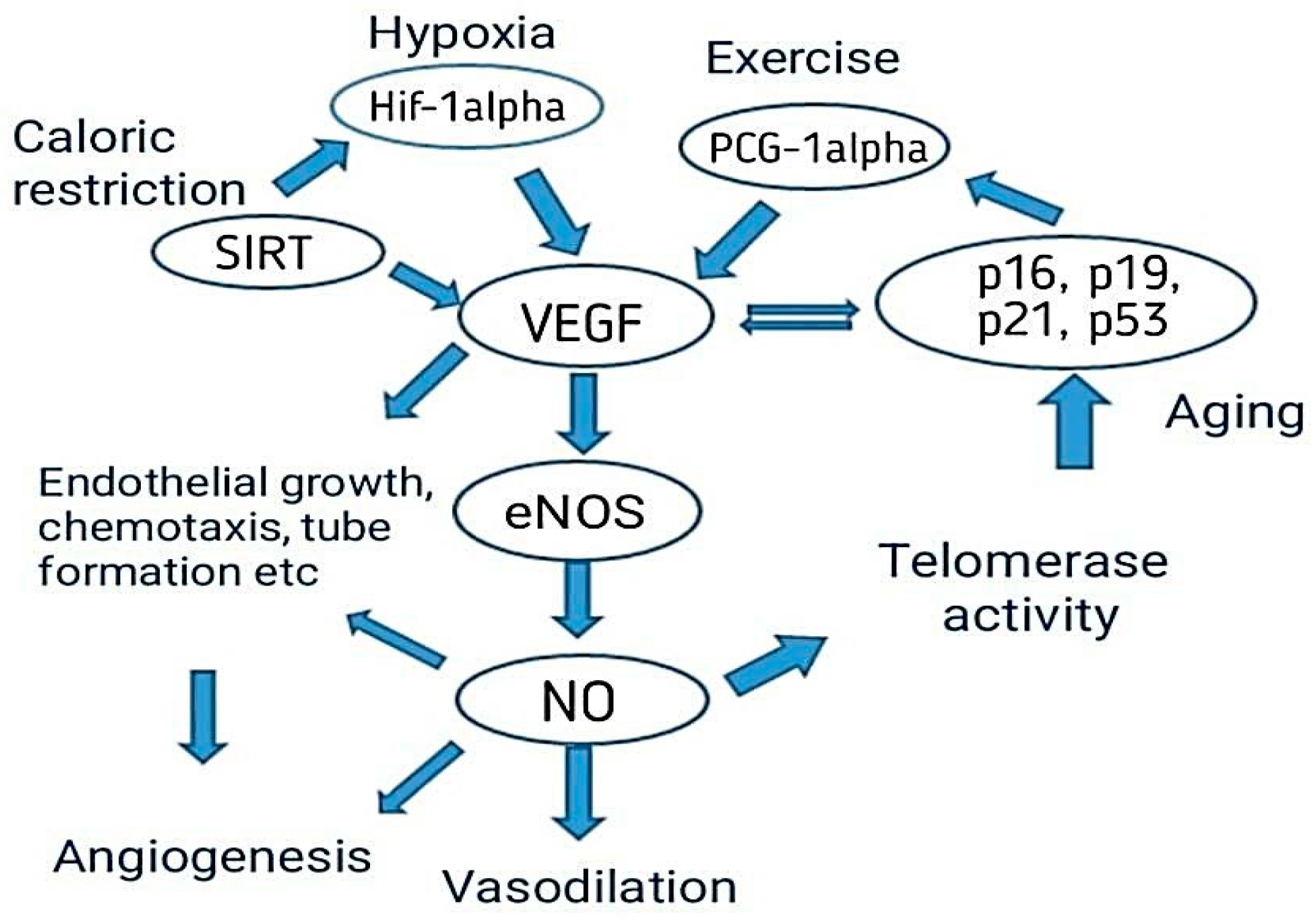
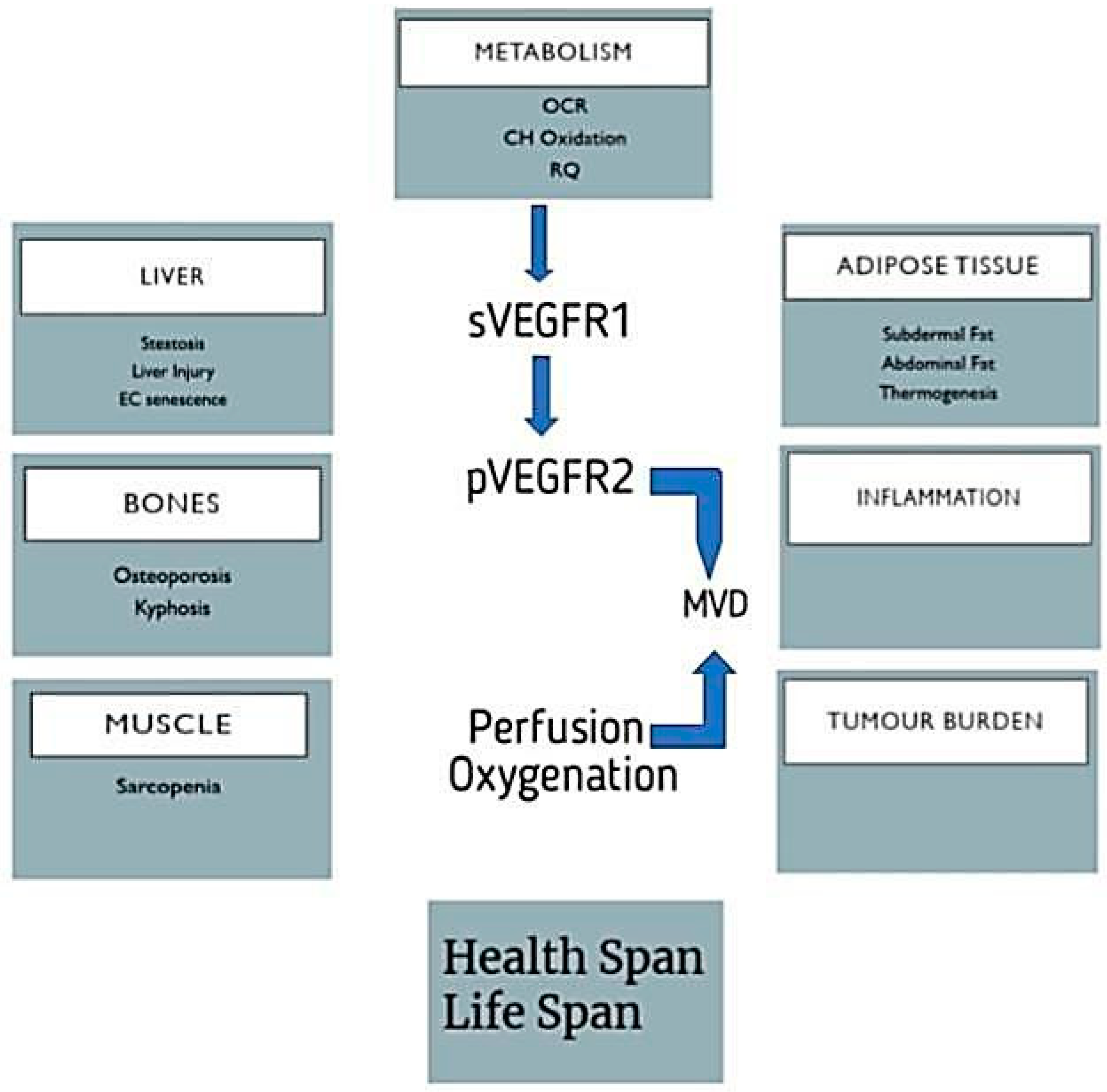
4.3. Aging Affects Angiogenesis
4.4. Role of VEGF to Delay Aging
4.5. Diet That Improves Vascular Function to Delay Aging
4.6. Role of FOLR1 in Aging
5. Intake of Folic Acid Leading to a Slow Aging Process
5.1. Effect of Deficiency of Folate in Elderly People
5.2. Dosage of Folic Acid by FDA
5.3. Folate-Containing Foods
6. Other Genes Affecting the Expressions of FOLR1 and VEGF
6.1. GATA6 Gene
- (a)
- direct activation of genes associated with maintaining stem cell characteristics (Oct4, Sox2, and Klf4 genes);
- (b)
- repression of miRNA biogenesis associated with these genes or their effector molecules, as demonstrated for Oct4 and its effector target Trim71.
6.2. CBS Gene
6.3. CISD2 Gene
6.4. SIRT1 and SIRT6 Genes
7. Associated Diseases
7.1. VEGF-Associated Diseases
7.2. FOLR1-Associated Diseases
8. Nanotherapeutics and Aging
9. Nanoparticles: As Drugs and Vaccine Delivery Systems via the Nasal Cavity
9.1. Nasal System

9.2. Zx+10a0 Nasal Vaccination: Why Nanoparticles?
9.3. Features of the Nose for the Delivery of Drugs and Vaccines
Activation of the Mucosal Immune System by Nanoparticles
- (1)
- effectors sites;
- (2)
- inductive sites.
- (1)
- barrier maintenance;
- (2)
- mucosal immune response initiation.
9.4. Nasal Drug and Vaccine Delivery and Nanoparticles
- (1)
- polysaccharide nanoparticles;
- (2)
- protein nanoparticles;
- (3)
- lipid nanoparticles;
- (4)
- polymers nanoparticles.
9.4.1. Polysaccharide-Based Nanoparticles
9.4.2. Chitosan Nanoparticles
9.4.3. Starch Nanoparticles
9.5. Polymer Nanoparticles
9.6. Lipid-Based Nanoparticles
Immune Stimulating Complexes (ISCOMs)
9.7. Protein-Based Nanoparticles
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Colombini, B.; Dinu, M.; Murgo, E.; Lotti, S.; Tarquini, R.; Sofi, F.; Mazzoccoli, G. Aging and low-level chronic inflammation: The role of the biological clock. Antioxidants 2022, 11, 2228. [Google Scholar] [CrossRef]
- Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; Della-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D. Oxidative stress, aging, and diseases. Clin. Interv. Aging 2018, 13, 757–772. [Google Scholar] [CrossRef] [PubMed]
- Adams, C.J.; Kopp, M.C.; Larburu, N.; Nowak, P.R.; Ali, M.M. structure and molecular mechanism of ER stress signaling by the unfolded protein response signal activator IRE1. Front. Mol. Biosci. 2019, 12, 11. [Google Scholar] [CrossRef] [PubMed]
- Szegezdi, E.; Logue, S.E.; Gorman, A.M.; Samali, A. Mediators of endoplasmic reticulum stress-induced apoptosis. EMBO Rep. 2006, 7, 880–885. [Google Scholar] [CrossRef]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef]
- Barbosa, M.C.; Grosso, R.A.; Fader, C.M. Hallmarks of aging: An autophagic perspective. Front. Endocrinol. 2019, 9, 790. [Google Scholar] [CrossRef] [PubMed]
- Oeppen, J.; Vaupel, J.W. Demography. Broken limits to life expectancy. Science 2002, 296, 1029–1031. [Google Scholar] [CrossRef]
- Bhatia, E.; Kumari, D.; Sharma, S.; Ahamad, N.; Banerjee, R. Nanoparticle platforms for dermal antiaging technologies: Insights in cellular and molecular mechanisms. WIREs Nanomed. Nanobiotech. 2021, 14, e1746. [Google Scholar] [CrossRef]
- Jasbi, N.E.; Dorranian, D. Effect of aging on the properties of TiO2 nanoparticle. J. Theor. Appl. Phys. 2016, 10, 157–161. [Google Scholar] [CrossRef]
- Wilmoth, J.R. The future of human longevity: A demographer’s perspective. Science 1998, 280, 395–397. [Google Scholar] [CrossRef]
- Park, T.J.; Lee, K.G.; Lee, S.Y. Advances in microbial biosynthesis of metal nanoparticles. Appl. Microbiol. Biotechnol. 2016, 100, 521–534. [Google Scholar] [CrossRef] [PubMed]
- Tosi, G.; Duskey, J.; Kreuter, J. Nanoparticles as carriers for drug delivery of macromolecules across the blood-brain barrier. Expert Opin. Drug Deliv. 2020, 17, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wu, B.; Wu, Y.; Song, X.; Zhang, S.; Liu, Z. Camouflaging nanoparticles with brain metastatic tumor cell membranes: A new strategy to traverse blood–brain barrier for imaging and therapy of brain tumors. Adv. Funct. Mater. 2020, 30, 1909369. [Google Scholar] [CrossRef]
- Jomehzadeh, N.; Koolivand, Z.; Dahdouh, E.; Akbari, A.; Zahedi, A.; Chamkouri, N. Investigating in-vitro antimicrobial activity, biosynthesis, and characterization of silver nanoparticles, zinc oxide nanoparticles, and silver-zinc oxide nanocomposites using Pistacia Atlantica Resin. Mater. Today Commun. 2021, 27, 102457. [Google Scholar] [CrossRef]
- Pedroso-Santana, S.; Fleitas-Salazar, N. Ionotropic gelation method in the synthesis of nanoparticles/microparticles for biomedical purposes. Polym. Int. 2020, 69, 443–447. [Google Scholar] [CrossRef]
- Rahman, M.; Khan, A.; Imtiyaz, Z.; Ali, S.; Makeen, H.A.; Rashid, S.; Arafah, A. Current nano-therapeutic approaches ameliorating inflammation in cancer progression. Semin. Cancer Biol. 2022, 86, 886–908. [Google Scholar] [CrossRef]
- Zhao, Z.; Wu, S.; Liu, Q.; Yang, C.; Zou, Y.; Wan, P. Feasibility assessment of CeO2 nanoparticles as aging-resistant agent of asphalt. Constr. Build. Mater. 2022, 330, 127245. [Google Scholar] [CrossRef]
- Wicki, A.; Witzigmann, D.; Balasubramanian, V.; Huwyler, J. Nanomedicine in cancer therapy: Challenges, opportunities, and clinical applications. J. Control. Release 2015, 200, 138–157. [Google Scholar] [CrossRef]
- Xu, H.; Li, S.; Liu, Y.S. Nanoparticles in the diagnosis and treatment of vascular aging and related diseases. Signal Transduct. Target. Ther. 2022, 7, 231. [Google Scholar] [CrossRef]
- Kubiak, J.M.; Macfarlane, R.J. Polymer-grafted nanoparticles as single-component, high filler content composites via simple transformative aging. Adv. Funct. Mater. 2022, 32, 2107139. [Google Scholar] [CrossRef]
- Moustafa, E.B.; Elsheikh, A.H.; Taha, M.A. The effect of TaC and NbC hybrid and mono-nanoparticles on AA2024 nanocomposites: Microstructure, strengthening, and artificial aging. Nanotechnol. Rev. 2022, 11, 2513–2525. [Google Scholar] [CrossRef]
- Xi, X.; Wang, L.; Zhou, T.; Yin, J.; Sun, H.; Yin, X.; Wang, N. Effects of physicochemical factors on the transport of aged polystyrene nanoparticles in saturated porous media. Chemosphere 2022, 289, 133239. [Google Scholar] [CrossRef] [PubMed]
- Obrador, A.; González, D.; Almendros, P.; García-Gómez, C.; Fernández, M.D. Assessment of phytotoxicity and behavior of 1-year-aged Zn in soil from ZnO nanoparticles, bulk ZnO, and Zn sulfate in different soil-plant cropping systems: From biofortification to toxicity. J. Soil Sci. Plant Nutr. 2022, 22, 150–164. [Google Scholar] [CrossRef]
- Yang, H.; Li, X.; Zhu, L.; Wu, X.; Zhang, S.; Huang, F.; Feng, X.L. Heat shock protein inspired nanochaperones restore amyloid-β homeostasis for preventative therapy of Alzheimer’s disease. Adv. Sci. 2019, 6, 1901844. [Google Scholar] [CrossRef]
- Yang, P.; Sheng, D.; Guo, Q.; Wang, P.; Xu, S.; Qian, K.; Li, Y.; Cheng, Y.; Wang, L.; Lu, W.; et al. Neuronal mitochondria-targeted micelles relieving oxidative stress for delayed progression of Alzheimer’s disease. Biomaterials 2020, 238, 119844. [Google Scholar] [CrossRef] [PubMed]
- Mirzaie, Z.; Ansari, M.; Kordestani, S.S.; Rezaei, M.H.; Mozafari, M. Preparation and characterization of curcumin-loaded polymeric nanomicelles to interference with amyloidogenesis through glycation method. Biotechnol. Appl. Biochem. 2019, 66, 537–544. [Google Scholar] [CrossRef]
- Igartúa, D.; Martinez, C.; Temprana, C.; Alonso, S.; Prieto, M. PAMAM dendrimers as a carbamazepine delivery system for neurodegenerative diseases: A biophysical and nanotoxicological characterization. Int. J. Pharm. 2018, 544, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Aso, E.; Martinsson, I.; Appelhans, D.; Effenberg, C.; Benseny-Cases, N.; Cladera, J.; Gouras, G.; Ferrer, I.; Klementieva, O. Poly(propylene imine) dendrimers with histidine-maltose shell as novel type of nanoparticles for synapse and memory protection. Nanomedicine 2019, 17, 198–209. [Google Scholar] [CrossRef]
- Gothwal, A.; Kumar, H.; Nakhate, K.T.; Ajazuddin Dutta, A.; Borah, A.; Gupta, U. Lactoferrin coupled lower generation pamam dendrimers for brain targeted delivery of memantine in aluminum-chloride-induced Alzheimer’s disease in mice. Bioconjug. Chem. 2019, 16, 2573–2583. [Google Scholar] [CrossRef]
- Rahman, M.; Rahaman, M.; Islam, M.; Rahman, F.; Mithi, F.; Alqahtani, T.; Almikhlafi, M.A.; Alghamdi, S.; Alruwaili, A.; Hossain, M. Role of phenolic compounds in human disease. Curr. Knowl. Future Prospect. 2021, 27, 233. [Google Scholar]
- Saleem, A.; Afzal, M.; Naveed, M.; Makhdoom, S.I.; Mazhar, M.; Aziz, T.; Khan, A.A.; Kamal, Z.; Shahzad, M.; Alharbi, M.; et al. HPLC, FTIR and GC-MS analyses of thymus vulgaris phytochemicals executing in vitro and in vivo biological activities and effects on COX-1, COX-2 and gastric cancer genes computationally. Molecules 2022, 27, 8512. [Google Scholar] [CrossRef] [PubMed]
- Naveed, M.; Bukhari, B.; Aziz, T.; Zaib, S.; Mansoor, M.A.; Khan, A.A.; Shahzad, M.; Dablool, A.S.; Alruways, M.W.; Almalki, A.A.; et al. Green synthesis of silver nanoparticles using the plant extract of Acer oblongifolium and study of its antibacterial and antiproliferative activity via mathematical approaches. Molecules 2022, 27, 4226. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; Islam, M.R.; Shohag, S.; Hossain, M.E.; Rahaman, M.S.; Islam, F.; Ahmed, M.; Mitra, S.; Khandaker, M.U.; Idris, A.M.; et al. The multifunctional role of herbal products in the management of diabetes and obesity: A comprehensive review. Molecules 2022, 6, 1713. [Google Scholar] [CrossRef] [PubMed]
- Naveed, M.; Batool, H.; Rehman, S.U.; Javed, A.; Makhdoom, S.I.; Aziz, T.; Mohamed, A.A.; Sameeh, M.Y.; Alruways, M.W.; Dablool, A.S.; et al. Characterization and evaluation of the antioxidant, antidiabetic, anti-inflammatory, and cytotoxic activities of silver nanoparticles synthesized using Brachychiton populneus leaf extract. Processes 2022, 10, 1521. [Google Scholar] [CrossRef]
- Rahman, M.M.; Rahaman, M.S.; Islam, M.R.; Hossain, M.E.; Mannan Mithi, F.; Ahmed, M.; Saldías, M.; Akkol, E.K.; Sobarzo-Sánchez, E. Multifunctional therapeutic potential of phytocomplexes and natural extracts for antimicrobial properties. Antibiotics 2021, 6, 1076. [Google Scholar] [CrossRef]
- Pandareesh, M.D.; Mythri, R.B.; Srinivas Bharath, M.M. Bioavailability of dietary polyphenols: Factors contributing to their clinical application in CNS diseases. Neurochem. Int. 2015, 89, 198–208. [Google Scholar] [CrossRef]
- Bengmark, S. Curcumin, An atoxic antioxidant and natural NFkappaB, cyclooxygenase-2, lipooxygenase, and inducible nitric oxide synthase inhibitor: A shield against acute and chronic diseases. J. Parenter. Enter. Nutr. 2006, 30, 45–51. [Google Scholar] [CrossRef]
- Wang, S.L.; Li, Y.; Wen, Y.; Chen, Y.F.; Na, L.X.; Li, S.T.; Sun, C.H. Curcumin, a potential inhibitor of up-regulation of TNF-alpha and IL-6 induced by palmitate in 3T3-L1 adipocytes through NF-kappaB and JNK pathway. Biomed. Env. Sci. 2009, 22, 32–39. [Google Scholar] [CrossRef]
- Guo, Y.Z.; He, P.; Feng, A.M. Effect of curcumin on expressions of NF-κBp65, TNF-α and IL-8 in placental tissue of premature birth of infected mice. Asian Pac. J. Trop. Med. 2017, 10, 175–178. [Google Scholar] [CrossRef]
- Taghipour, Y.D.; Hajialyani, M.; Naseri, R.; Hesari, M.; Mohammadi, P.; Stefanucci, A.; Mollica, A.; Farzaei, M.H.; Abdollahi, M. Nanoformulations of natural products for management of metabolic syndrome. Int. J. Nanomed. 2019, 16, 5303–5321. [Google Scholar] [CrossRef]
- Naqvi, S.; Panghal, A.; Flora, S.J.S. Nanotechnology: A Promising Approach for Delivery of Neuroprotective Drugs. Front. Neurosci. 2020, 14, 494. [Google Scholar] [CrossRef] [PubMed]
- Huwyler, J.; Wu, D.; Pardridge, W. Brain drug delivery of small molecules using immunoliposomes. Proc. Natl. Acad. Sci. USA 1996, 93, 14164–14169. [Google Scholar] [CrossRef]
- Kozubek, A.; Gubernator, J.; Przeworska, E.; Stasiuk, M. Liposomal drug delivery, a novel approach: PLARosomes. Acta Biochim. Pol. 2000, 47, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Voinea, M.; Simionescu, M. Designing of ‘intelligent’ liposomes for efficient delivery of drugs. J. Cell. Mol. Med. 2002, 6, 465–474. [Google Scholar] [CrossRef] [PubMed]
- Alyaudtin, R.N.; Reichel, A.; Löbenberg, R.; Ramge, P.; Kreuter, J.; Begley, D.J. Interaction of poly (butylcyanoacrylate) nanoparticles with the blood-brain barrier in vivo and in vitro. J. Drug Target 2001, 9, 209–221. [Google Scholar] [CrossRef]
- Roney, C.; Kulkarni, P.; Arora, V.; Antich, P.; Bonte, F.; Wu, A.; Mallikarjuana, N.; Manohar, S.; Liang, H.-F.; Kulkarni, A. Targeted nanoparticles for drug delivery through the blood–brain barrier for Alzheimer’s disease. J. Control. Release 2005, 108, 193–214. [Google Scholar] [CrossRef]
- Geesaman, B.J.; Benson, E.; Brewster, S.J.; Kunkel, L.M.; Blanche, H.; Thomas, G.; Perls, T.T.; Daly, M.J.; Puca, A.A. Haplotype-based identification of a microsomal transfer protein marker associated with the human lifespan. Proc. Natl. Acad. Sci. USA 2003, 100, 115–120. [Google Scholar] [CrossRef]
- Corder, E.H.; Saunders, A.M.; Strittmatter, W.J.; Schmechel, D.E.; Gaskell, P.C.; Small, G.W.; Roses, A.D.; Haines, J.L.; Pericak-Vance, M.A. Gene dose of apolipoprotein E type 4 allele and the risk of Alzheimer’s disease in late onset families. Science 1993, 261, 921–923. [Google Scholar] [CrossRef]
- Liu, X.; Chen, C. Strategies to optimize brain penetration in drug discovery. Curr. Opin. Drug Discov. Dev. 2005, 8, 505–512. [Google Scholar]
- Vinogradov, S.; Batrakova, E.; Kabanov, A. Poly (ethylene glycol)– polyethyleneimine NanoGelTM particles: Novel drug delivery systems for antisense oligonucleotides. Colloids Surf. B Biointerfaces 1999, 16, 291–304. [Google Scholar] [CrossRef]
- Moghimi, S.; Illum, L.; Davis, S. Physiopathological and physicochemical considerations in targeting of colloids and drug carriers to the bone marrow. Crit. Rev. Ther. Drug Carr. Syst. 1990, 7, 187–209. [Google Scholar]
- Wu, Y.Z.; Tsai, Y.Y.; Chang, L.S.; Chen, Y.J. Evaluation of Gallic Acid-Coated Gold Nanoparticles as an Anti-Aging Ingredient. Pharmaceuticals 2021, 14, 1071. [Google Scholar] [CrossRef]
- Zhang, W.; Chen, X.; Ma, Y.; Xu z Wu, L.; Yang, Y.; Tsang, S.; Chen, S. Positive Aging Effect of ZnO Nanoparticles Induced by Surface Stabilization. J. Phys. Chem. Lett. 2020, 11, 5863–5870. [Google Scholar] [CrossRef]
- Zang, S.; Duan, E. Fighting against skin aging. Cell Transplant. Beijing 2018, 27, 729–738. [Google Scholar] [CrossRef]
- Wlaschek, M.; Tantcheva-Poór, I.; Naderi, L.; Ma, W.; Schneider, L.A.; Razi-Wolf, Z.; Scharffetter-Kochanek, K. Solar UV irradiation and dermal photoaging. J. Photochem. Photobiol. B Biol. 2001, 63, 41–51. [Google Scholar] [CrossRef]
- Atzmon, G.; Rincon, M.; Schechter, C.B.; Shuldiner, A.R.; Lipton, R.B.; Bergman, A.; Barzilai, N. Lipoprotein genotype and conserved pathway for exceptionallongevity in humans. PLoS Biol. 2006, 4, e113. [Google Scholar] [CrossRef] [PubMed]
- Yaşar, P.; Ayaz, G.; User, S.D.; Güpür, G.; Muyan, M. Molecular mechanism of estrogen–estrogen receptor signaling. Reprod. Med. Biol. 2017, 16, 4–20. [Google Scholar] [CrossRef]
- McGue, M.; Vaupel, J.W.; Holm, N.; Harvald, B. Longevity is moderately a heritable in a sample of Danish twins born. J. Gerontol. 1993, 48, 1870–1880. [Google Scholar] [CrossRef]
- Corder, E.H.; Saunders, N.J.; Risch, W.J.; Strittmatter, D.E.; Schmechel, P.C.; Gaskell, J.B.; Rimmler, J.B.; Locke, P.A.; Conneally, P.M.; Schmader, K.E.; et al. Protective effect of apolipoprotein E type 2 allele for late onset Alzheimer disease. Nat. Gen. 1994, 7, 180–184. [Google Scholar] [CrossRef] [PubMed]
- Kervinen, K.; Savolainen, M.J.; Salokannel, J.; Hynninen, A.; Heikkinen, J.; Ehnholm, C.; Koistinen, M.J.; Kesäniemi, Y.A. Apolipoprotein E and B polymorphisms–longevity factors assessed in nonagenarians. Atherosclerosis 1994, 105, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Schachter, F.; Faure-Delanef, L.; Guenot, F.; Rouger, H.; Froguel, P.; Lesueur-Ginot, L.; Daniel, C. Genetic associations with human longevity at the APOE and ACEloci. Nat. Genet. 1994, 6, 29–32. [Google Scholar] [CrossRef]
- Jiang, Q.; Lee, C.D.; Mandrekar, S.; Wilkinson, B.; Cramer, P.; Zelcer, N.; Mann, K.; Lamb, B.; Willson, T.M.; Collins, J.L.; et al. ApoE promotes the proteolytic degradation of Aβ. Neuron 2008, 58, 681–693. [Google Scholar] [CrossRef]
- Kenyon, C.; Chang, J.; Gensch, E.; Rudner, A.; Tabtiang, R. Elegans mutant that lives twice as long as wild type. Nature 1993, 366, 461–464. [Google Scholar] [CrossRef] [PubMed]
- Clancy, D.J.; Gems, D.; Harshman, L.G.; Oldham, S.; Stocker, H.; Hafen, E.; Leevers, S.J.; Partridge, L. Extension of lifespan by loss of CHICO, a Drosophila insulin receptor substrate protein. Science 2001, 292, 104–106. [Google Scholar] [CrossRef]
- Bluher, M.; Kahn, B.B.; Kahn, C.R. Extended longevity in mice lacking the insulin receptor in adipose tissue. Science 2003, 299, 572–574. [Google Scholar] [CrossRef]
- Karasik, D.; Demissie, S.; Cupples, L.A.; Kiel, D.P. Disentangling the genetic determinants of human aging: Biological age as an alternative to the use of survival measures. J. Gerontol. A Biol. Sci. Med. Sci. 2005, 60, 574–587. [Google Scholar] [CrossRef]
- Lähteenvuo, J.; Rosenzweig, A. Effects of aging on angiogenesis. Circ. Res. 2012, 27, 1252–1264. [Google Scholar] [CrossRef]
- Longchamp, A.; Mirabella, T.; Arduini, A.; MacArthur, M.R.; Das, A.; Treviño-Villarreal, J.H.; Hine, C.; Ben-Sahra, I.; Knudsen, N.H.; Brace, L.E.; et al. Amino Acid Restriction Triggers Angiogenesis via GCN2/ATF4 Regulation of VEGF and H2S Production. Cell 2018, 173, 117–129. [Google Scholar] [CrossRef] [PubMed]
- Shooshtari, M.K.; Moazedi, A.A.; Parham, G.A. Memory and motor coordination improvement by folic Acid supplementation in healthy adult male rats. Iran J. Basic Med. Sci. 2012, 15, 1173–1179. [Google Scholar]
- Bailey, L.B.; Stover, P.J.; McNulty, H. Biomarkers of nutrition for development-folate review. J. Nutr. 2015, 145, 1636S–1680S. [Google Scholar] [CrossRef] [PubMed]
- Kuang, J.; Chen, L.; Tang, Q.; Zhang, J.; Li, Y.; He, J. The Role of Sirt6 in Obesity and Diabetes. Front. Physiol. 2018, 9, 135. [Google Scholar] [CrossRef]
- Jiao, H.; Walczak, B.E.; Lee, M.S.; Lemieux, M.E.; Li, W.J. GATA6 regulates aging of human mesenchymal stem/stromal cells. Stem Cells. 2021, 39, 62–77. [Google Scholar] [CrossRef]
- Browder, K.C.; Pradeep, R.; Yamamoto, M.; Haghani, A.; Guillen, I.G.; Sahu, S.; Wang, C.; Luque, Y.; Prieto, J.; Shi, L.; et al. In vivo partial reprogramming alters age-associated molecular changes during physiological aging in mice. Nat. Aging 2022, 2, 243–253. [Google Scholar] [CrossRef]
- Mohanty, V.; Shah, A.; Allender, E.; Siddiqui, M.R.; Monick, S.; Ichi, S.; Mania-Farnell, B.; GMcLone, D.; Tomita, T.; Mayanil, C.S. Folate receptor alpha upregulates Oct4, Sox2 and Klf4 and downregulates miR-138 andmiR-let-7 in cranial neural crest cells. Stem Cells 2016, 34, 2721–2732. [Google Scholar] [CrossRef]
- Zhao, Q.; Zhang, C.; Li, D.; Huang, X.; Ren, B.; Yue, L.; Du, B.; Godfrey, O.; Zhang, W. CBS gene polymorphism and promoter methylation-mediating effects on the efficacy of folate therapy in patients with hyperhomocysteinemia. J. Gene Med. 2020, 22, e3156. [Google Scholar] [CrossRef]
- Hall, J.A.; Dominy, J.E.; Lee, Y.; Puigserver, P. The sirtuin family’s role in aging and age-associated pathologies. J. Clin. Investig. 2013, 123, 973–979. [Google Scholar] [CrossRef]
- Michishita, E.; McCord, R.A.; Boxer, L.D.; Barber, M.F.; Hong, T.; Gozani, O. Cell cycle-dependent deacetylation of telomeric histone H3 lysine K56 by human SIRT6. Cell Cycle 2009, 8, 2664–2666. [Google Scholar] [CrossRef]
- Yang, S.J.; Choi, J.M.; Chae, S.W.; Kim, W.J.; Park, S.E.; Rhee, E.J. Activation of peroxisome proliferator-activated receptor gamma by rosiglitazone increases sirt6 expression and ameliorates hepatic steatosis in rats. PLoS ONE 2011, 6, e17057. [Google Scholar] [CrossRef]
- Vitiello, M.; Zullo, A.; Servillo, L.; Mancini, F.P.; Borriello, A.; Giovane, A. Multiple pathways of SIRT6 at the crossroads in the control of longevity, cancer, and cardiovascular diseases. Aging Res. Rev. 2017, 35, 301–311. [Google Scholar] [CrossRef]
- Oh, J.H.; Karadeniz, F.; Lee, J.I.; Park, S.Y.; Seo, Y.; Kong, C.S. Anticatabolic and anti-inflammatory effects of myricetin 3-O-β-d-galactopyranoside in UVA-irradiated dermal cells via repression of MAPK/AP-1 and activation of TGFβ/Smad. Molecules 2020, 25, 1331. [Google Scholar] [CrossRef]
- Polito, F.; Marini, H.; Bitto, A.; Irrera, N.; Vaccaro, M.; Adamo, E.B.; Altavilla, D. Genistein aglycone, a soy-derived isoflavone, improves skin changes induced by ovariectomy in rats. Br. J. Pharmacol. 2012, 165, 994–1005. [Google Scholar] [CrossRef]
- Zhao, X.; Liu, Y.; Jia, P.; Cheng, H.; Wang, C.; Chen, S.; Li, Z. Chitosan hydrogel-loaded MSC-derived extracellular vesicles promote skin rejuvenation by ameliorating the senescence of dermal fibroblasts. Stem Cell Res. Ther. 2021, 12, 196. [Google Scholar] [CrossRef]
- Karthivashan, G.; Ganesan, P.; Park, S.Y.; Kim, J.S.; Choi, D.K. Therapeutic strategies and nano-drug delivery applications in management of aging Alzheimer’s disease. Drug Deliv. 2018, 25, 307–320. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo-Veiga, B.; Alvarez-Lorenzo, C.; Loftsson, T.; Sigurdsson, H.H. Age-related ocular conditions: Current treatments and role of cyclodextrin-based nanotherapies. Int. J. Pharm. 2021, 603, 120707. [Google Scholar] [CrossRef]
- Kut, C.; Mac Gabhann, F.; Popel, A.S. Where is VEGF in the body? A meta-analysis of VEGF distribution in cancer. Br. J. Cancer 2007, 97, 978–985. [Google Scholar] [CrossRef] [PubMed]
- Holmes, D.I.; Zachary, I. The vascular endothelial growth factor (VEGF) family: Angiogenic factors in health and disease. Genome Biol. 2005, 6, 209. [Google Scholar] [CrossRef]
- Safran, M.; Rosen, N.; Twik, M.; BarShir, R.; Iny Stein, T.; Dahary, D.; Fishilevich, S. The GeneCards Suite Chapter, Practical Guide to Life Science Databases. Lancet 2021, 48, 27–56. [Google Scholar]
- Jalal, S.; Younis, M.Z. Aging and Elderly in Pakistan. Aging Int. 2014, 39, 4–12. [Google Scholar] [CrossRef]
- Maßberg, D.; Hatt, H. Human Olfactory Receptors: Novel Cellular Functions Outside of the Nose. Physiol. Rev. 2018, 98, 1739–1763. [Google Scholar] [CrossRef]
- Weinhold, I.; Mlynski, G. Numerical simulation of airflow in the human nose. Eur. Arch. Otorhinolaryngol. 2004, 261, 452–455. [Google Scholar] [CrossRef] [PubMed]
- Berta, A.; Ekdale, E.G.; Cranford, T.W. Review of the cetacean nose: Form, function, and evolution. Anat. Rec. 2014, 297, 2205–2215. [Google Scholar] [CrossRef]
- Jenkins, E.K.; DeChant, M.T.; Perry, E.B. When the nose doesn’t know: Canine olfactory function associated with health, management, and potential links to microbiota. Front. Vet. Sci. 2018, 56, 1–16. [Google Scholar] [CrossRef]
- Csaba, N.; Garcia-Fuentes, M.; Alonso, M.J. Nanoparticles for nasal vaccination. Adv. Drug Deliv. Rev. 2009, 61, 140–157. [Google Scholar] [CrossRef] [PubMed]
- Mara Mainardes, R.; Cristina Cocenza Urban, M.; Oliveira Cinto, P.; Vinicius Chaud, M.; Cesar Evangelista, R.; Palmira Daflon Gremiao, M. Liposomes and micro/nanoparticles as colloidal carriers for nasal drug delivery. Curr. Drug Deliv. 2006, 3, 275–285. [Google Scholar] [CrossRef]
- Kim, B.G.; Kang, I.J. Evaluation of the effects of biodegradable nanoparticles on a vaccine delivery system using AFM, SEM, and TEM. Ultramicroscopy 2008, 108, 1168–1173. [Google Scholar] [CrossRef]
- Salatin, S.; Barar, J.; Barzegar-Jalali, M.; Adibkia, K.; Milani, M.A.; Jelvehgari, M. Hydrogel nanoparticles and nanocomposites for nasal drug/vaccine delivery. Arch. Pharmacal. Res. 2016, 39, 1181–1192. [Google Scholar] [CrossRef]
- Ali, J.; Ali, M.; Baboota, S.; Kaur Sahni, J.; Ramassamy, C.; Dao, L. Potential of nanoparticulate drug delivery systems by intranasal administration. Curr. Pharm. Des. 2010, 16, 1644–1653. [Google Scholar] [CrossRef]
- Ho, W.; Gao, M.; Li, F.; Li, Z.; Zhang, X.Q.; Xu, X. Next-Generation Vaccines: Nanoparticle-Mediated DNA and mRNA Delivery. Adv. Healthc. Mater. 2021, 10, 2001812. [Google Scholar] [CrossRef] [PubMed]
- Naveed, M.; Sheraz, M.; Amin, A.; Waseem, M.; Aziz, T.; Khan, A.A.; Ghani, M.; Shahzad, M.; Alruways, M.W.; Dablool, A.S.; et al. Designing a novel peptide-based multi-epitope vaccine to evoke a robust immune response against pathogenic multidrug-resistant Providencia heimbachae. Vaccines 2022, 10, 1300. [Google Scholar] [CrossRef]
- Naveed, M.; Makhdoom, S.I.; Ali, U.; Jabeen, K.; Aziz, T.; Khan, A.A.; Jamil, S.; Shahzad, M.; Alharbi, M.; Alshammari, A. Immunoinformatics approach to design multi-epitope-based vaccine against machupo virus taking viral nucleocapsid as a potential candidate. Vaccines 2022, 10, 1732. [Google Scholar] [CrossRef] [PubMed]
- Naveed, M.; Mughal, M.S.; Jabeen, K.; Aziz, T.; Naz, S.; Nazir, N.; Shahzad, M.; Alharbi, M.; Alshammari, A.; Sadhu, S.S. Evaluation of the whole proteome to design a novel mRNA-based vaccine against multidrug-resistant Serrati marcescens. Front. Microbiol. 2022, 13, 960285. [Google Scholar] [CrossRef] [PubMed]
- Cardoso VM, D.O.; Moreira, B.J.; Comparetti, E.J.; Sampaio, I.; Ferreira LM, B.; Lins PM, P.; Zucolotto, V. Is nanotechnology helping in the fight against COVID-19? Front Nanotechnol 2020, 4, 1–30. [Google Scholar] [CrossRef]
- Shim, S.; Yoo, H.S. The application of mucoadhesive chitosan nanoparticles in nasal drug delivery. Mar. Drugs 2020, 18, 605. [Google Scholar] [CrossRef]
- Rabiee, N.; Ahmadi, S.; Afshari, R.; Khalaji, S.; Rabiee, M.; Bagherzadeh, M.; Fatahi, Y.; Dinarvand, R.; Tahriri, M.; Tayebi, L. Polymeric nanoparticles for nasal drug delivery to the brain: Relevance to Alzheimer’s disease. Adv. Ther. 2021, 4, 2000076. [Google Scholar] [CrossRef]
- Clementino, A.R.; Pellegrini, G.; Banella, S.; Colombo, G.; Cantù, L.; Sonvico, F.; Del Favero, E. Structure and fate of nanoparticles designed for the nasal delivery of poorly soluble drugs. Mol. Pharm. 2021, 18, 3132–3146. [Google Scholar] [CrossRef]
- Chatzitaki, A.-T.; Jesus, S.; Karavasili, C.; Andreadis, D.; Fatouros, D.G.; Borges, O. Chitosan-coated PLGA nanoparticles for the nasal delivery of ropinirole hydrochloride: In vitro and ex vivo evaluation of efficacy and safety. Int. J. Pharm. 2020, 589, 119776. [Google Scholar] [CrossRef] [PubMed]
- Hao, R.; Sun, B.; Yang, L.; Ma, C.; Li, S. RVG29-modified microRNA-loaded nanoparticles improve ischemic brain injury by nasal delivery. Drug Deliv. 2020, 27, 772–781. [Google Scholar] [CrossRef]
- Mahallawi, W.H.; Aljeraisi, T.M. Infection with SARS-CoV-2 primes immunological memory in human nasal-associated lymphoid tissue. Clin. Immunol. 2021, 231, 108850. [Google Scholar] [CrossRef]
- Serfling, S.; Zhi, Y.; Schirbel, A.; Lindner, T.; Meyer, T.; Gerhard-Hartmann, E.; Lapa, C.; Hagen, R.; Hackenberg, S.; Buck, A. Improved cancer detection in Waldeyer’s tonsillar ring by 68Ga-FAPI PET/CT imaging. Eur. J. Nucl. Med. Mol. Imaging 2021, 48, 1178–1187. [Google Scholar] [CrossRef]
- Mahallawi, W.H.; Aljeraisi, T.M. In vitro cell culture model of human nasal associated lymphoid tissue (NALT) to evaluate the humoral immune response to SARS-CoV 2 spike proteins. Saudi J. Biol. Sci. 2021, 28, 4516–4521. [Google Scholar] [CrossRef] [PubMed]
- Aljurayyan, A.; Puksuriwong, S.; Ahmed, M.; Sharma, R.; Krishnan, M.; Sood, S.; Davies, K.; Rajashekar, D.; Leong, S.; McNamara, P.S. Activation and induction of antigen specific T follicular helper cells play a critical role in live-attenuated influenza vaccine induced human mucosal anti-influenza antibody response. J. Virol. 2018, 92, e00114–e00118. [Google Scholar] [CrossRef]
- Hussain, N.; Mumtaz, M.; Adil, M.; Waseem, M. ACE I/D polymorphism and In-Silico screening of potential bioactive phytochemicals against COVID-19. Bioinform. Biol. Insights. 2022, 2, 23–45. [Google Scholar]
- Hussain, N.; Falek Sher, S.; Ming, L.; Adil, M. Association of VEGF gene polymorphism (rs699947) with Glaucoma and In-silico study of antiglaucoma bioactive chemicals. Appl. Biochem. Biotechnol. 2022, 194, 5185–5195. [Google Scholar] [CrossRef]
- Panda, S.K.; Colonna, M. Innate lymphoid cells in mucosal immunity. Front. Immunol. 2019, 10, 861. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Jin, L.; Chen, T. The effects of secretory IgA in the mucosal immune system. BioMed Res. Int. 2020, 2020, 2032057. [Google Scholar] [CrossRef]
- Dillon, A.; Lo, D.D. M cells: Intelligent engineering of mucosal immune surveillance. Front. Immunol. 2019, 10, 1499. [Google Scholar] [CrossRef] [PubMed]
- Kortekaas Krohn, I.; Seys, S.F.; Lund, G.; Jonckheere, A.C.; Dierckx de Casterlé, I.; Ceuppens, J.L.; Steelant, B.; Hellings, P.W. Nasal epithelial barrier dysfunction increases sensitization and mast cell degranulation in the absence of allergic inflammation. Allergy 2020, 75, 1155–1164. [Google Scholar] [CrossRef]
- Zhao, R.; Guo, Z.; Zhang, R.; Deng, C.; Xu, J.; Dong, W.; Hong, Z.; Yu, H.; Situ, H.; Liu, C. Nasal epithelial barrier disruption by particulate matter ≤2.5 μm via tight junction protein degradation. J. Appl. Toxicol. 2018, 38, 678–687. [Google Scholar] [CrossRef]
- Bernocchi, B.; Carpentier, R.; Betbeder, D. Nasal nanovaccines. Int. J. Pharm. 2017, 530, 128–138. [Google Scholar] [CrossRef] [PubMed]
- Bento, D.; Staats, H.F.; Borges, O. Effect of particulate adjuvant on the anthrax protective antigen dose required for effective nasal vaccination. Vaccine 2015, 33, 3609–3613. [Google Scholar] [CrossRef]
- Pawar, D.; Jaganathan, K. Mucoadhesive glycol chitosan nanoparticles for intranasal delivery of hepatitis B vaccine: Enhancement of mucosal and systemic immune response. Drug Deliv. 2016, 23, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Debin, A.; Kravtzoff, R.; Santiago, J.V.; Cazales, L.; Sperandio, S.; Melber, K.; Janowicz, Z.; Betbeder, D.; Moynier, M. Intranasal immunization with recombinant antigens associated with new cationic particles induces strong mucosal as well as systemic antibody and CTL responses. Vaccine 2002, 20, 2752–2763. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Zhang, Z.; Lv, J.; Zhou, P.; Hu, W.; Fang, Y.; Chen, H.; Liu, X.; Shao, J.V.; Zhao, F. Induction of mucosal immune responses and protection of cattle against direct contact challenge by intranasal delivery with foot-and-mouth disease virus antigen mediated by nanoparticles. Int. J. Nanomed. 2014, 9, 5603. [Google Scholar] [CrossRef] [PubMed]
- Ninomiya, A.; Ogasawara, K.; Kajino, K.; Takada, A.V.; Kida, H. Intranasal administration of a synthetic peptide vaccine encapsulated in liposome together with an anti CD40 antibody induces protective immunity against influenza A virus in mice. Vaccine 2002, 20, 3123–3129. [Google Scholar] [CrossRef] [PubMed]
- Cibulski, S.P.; Mourglia-Ettlin, G.; Teixeira, T.F.; Quirici, L.; Roehe, P.M.; Ferreira, F.; Silveira, F. Novel ISCOMs from Quillaja brasiliensis saponins induce mucosal and systemic antibody production, T-cell responses and improved antigen uptake. Vaccine 2016, 34, 1162–1171. [Google Scholar] [CrossRef] [PubMed]
- Misstear, K.; McNeela, E.A.; Murphy, A.G.; Geoghegan, J.A.; O’Keeffe, K.M.; Fox, J.; Chan, K.; Heuking, S.; Collin, N.; Foster, T.J. Targeted nasal vaccination provides antibody-independent protection against Staphylococcus aureus. J. Infect. Dis. 2014, 209, 1479–1484. [Google Scholar] [CrossRef] [PubMed]
- Rice-Ficht, A.C.; Arenas-Gamboa, A.M.; Kahl-McDonagh, M.M.; Ficht, T.A. Polymeric particles in vaccine delivery. Curr. Opin. Microbiol. 2010, 13, 106–112. [Google Scholar] [CrossRef]
- Zaman, M.; Chandrudu, S.; Giddam, A.K.; Reiman, J.; Skwarczynski, M.; McPhun, V.; Moyle, P.M.; Batzloff, M.R.; Good, M.F.; Toth, I. Group A Streptococcal vaccine candidate: Contribution of epitope to size, antigen presenting cell interaction and immunogenicity. Nanomedicine 2014, 9, 2613–2624. [Google Scholar] [CrossRef]
- Marasini, N.; Giddam, A.K.; Khalil, Z.G.; Hussein, W.M.; Capon, R.J.; Batzloff, M.R.; Good, M.F.; Toth, I.; Skwarczynski, M. Double adjuvanting strategy for peptide-based vaccines: Trimethyl chitosan nanoparticles for lipopeptide delivery. Nanomedicine 2016, 11, 3223–3235. [Google Scholar] [CrossRef]
- Doavi, T.; Mousavi, S.L.; Kamali, M.; Amani, J.; Ramandi, M.F. Chitosan-based intranasal vaccine against Escherichia coli O157, H7. Iran. Biomed. J. 2016, 20, 97. [Google Scholar]
- Dhakal, S.; Renu, S.; Ghimire, S.; Shaan Lakshmanappa, Y.; Hogshead, B.T.; Feliciano Ruiz, N.; Lu, F.; HogenEsch, H.; Krakowka, S.; Lee, C.W. Mucosal immunity and protective efficacy of intranasal inactivated influenza vaccine is improved by chitosan nanoparticle delivery in pigs. Front. Immunol. 2018, 9, 934. [Google Scholar] [CrossRef]
- Renu, S.; Renukaradhya, G.J. Chitosan nanoparticle based mucosal vaccines delivered against infectious diseases of poultry and pigs. Front. Bioeng. Biotechnol. 2020, 8, 558349. [Google Scholar] [CrossRef] [PubMed]
- Vila, A.; Sánchez, A.; Pérez, C.; Alonso, M.J. PLA-PEG nanospheres: New carriers for transmucosal delivery of proteins and plasmid DNA. Polym. Adv. Technol. 2002, 13, 851–858. [Google Scholar]
- Yan, X.; Diao, M.; Yu, Y.; Gao, F.; Wang, E.; Wang, Z.; Zhang, T. Influence of esterification and ultrasound treatment on formation and properties of starch nanoparticles and their impact as a filler on chitosan-based films characteristics. Int. J. Biol. Macromol. 2021, 179, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Akel, H.; Ismail, R.; Katona, G.; Sabir, F.; Ambrus, R.; Csóka, I. A comparison study of lipid and polymeric nanoparticles in the nasal delivery of meloxicam: Formulation, characterization, and in vitro evaluation. Int. J. Pharm. 2021, 604, 120724. [Google Scholar] [CrossRef] [PubMed]
- Mwema, A.; Bottemanne, P.; Paquot, A.; Ucakar, B.; Vanvarenberg, K.; Alhouayek, M.; Muccioli, G.G.; des Rieux, A. Lipid nanocapsules for the nose-to-brain delivery of the anti-inflammatory bioactive lipid PGD2-G. Nanotechnol. Biol. Med. 2022, 48, 102633. [Google Scholar] [CrossRef] [PubMed]
- Ramvikas, M.; Arumugam, M.; Chakrabarti, S.; Jaganathan, K. Nasal vaccine delivery. In Micro & Nanotechnology in Vaccine Development; Elsevier: Amsterdam, The Netherlands, 2017; pp. 279–301. [Google Scholar]
- Pacini, M.F.; González, F.B.; Dinatale, B.; Balbi, C.B.; Villar, S.R.; Farré, C.; Lupi, G.; Espariz, M.; Blancato, V.S.; Magni, C. Nasal immunization with a L. lactis-derived trans-sialidase antigen plus c-di-AMP protects against acute oral T. cruzi infection. Vaccine 2022, 40, 2311–2323. [Google Scholar] [CrossRef]
- López-Sagaseta, J.; Malito, E.; Rappuoli, R.; Bottomley, M.J. Self-assembling protein nanoparticles in the design of vaccines. Comput. Struct. Biotechnol. 2016, 14, 58–68. [Google Scholar] [CrossRef]
- Witthöft, C.M.; Forssén, K.; Johannesson, L.; Jägerstad, M. Folates—Food sources, analyses, retention and bioavailability. J. Food Nutr. Res. 1999, 43, 138–146. [Google Scholar] [CrossRef]
- De Carvalho, M.M.; Stamford, T.C.; Dos Santos, E.P.; Tenorio, P.; Sampaio, F. Chitosan as an oral antimicrobial agent. In Science against Microbial Pathogens: Communicating Current Research and Technological Advances; Formatex Research Center: Badajoz, Spain, 2011; pp. 542–551. [Google Scholar]
- Hayat, P.; Khan, I.; Rehman, A.; Jamil, T.; Hayat, A.; Rehman, M.U.; Ullah, N.; Sarwar, A.; Alharbi, A.A.; Dablool, A.S.; et al. Myogenesis and Analysis of Antimicrobial Potential of Silver Nanoparticles (AgNPs) against Pathogenic Bacteria. Molecules 2023, 28, 637. [Google Scholar] [CrossRef]
- Kaczmarek, J.C.; Kowalski, P.S.; Anderson, D.G. Advances in the delivery of RNA therapeutics: From concept to clinical reality. Gen. Med. 2017, 9, 60. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Features | Changes with Aging | Effects on Nanotherapeutics (NPs) |
|---|---|---|
| Absorption | ||
| Gastric acid | Decreased secretion | Reduced degradation |
| Microbiota | Reduced diversity and leaky gut | Increased absorption time |
| Gut motility | Delayed gastric emptying | Potential increase in toxicity |
| Injectable permeability | Injection absorption unaffected | Injectable NPs are unaffected |
| Distribution | ||
| Plasma proteins | Reduced circulating albumin | May reduce protein corona |
| Body composition | Decreased water and increased fat contents | May reduce the distribution of water-soluble NPs and increase the distribution of fat-soluble NPs. |
| Cellular uptake | ||
| Phagocytosis | Reduced clearance and high circulating inflammatory markers | Unaffected NPs uptake |
| Endocytosis | By LSECs is slightly reduced | Largely unaffected |
| Transcytosis | By LSECs is unaffected | Unaffected |
| Passive uptake | Decreased due to pseudocappilarization | Reduced NP uptake |
| Metabolism | ||
| Phase I | Reduced due to hepatic blood flow | Prolonged circulation time |
| Phase II | Reduced | Unaffected |
| Type of Particles | Particles Characteristics | Model | Immunization Parameters | Immunity Response | Reference | |||
|---|---|---|---|---|---|---|---|---|
| Size (nm) | Z (mV) | Admi. | Ag Dose (µg) | Anes. | ||||
| Chitosan | 300–680 | 26 | Mice | 3 × 15 µL | 2.5 | Yes | h, m | (Bento et al.) [120] |
| 80 | 14 | Mice | 2 × 10 µL | 10 | Yes | h, m | (Pawar et al.) [121] | |
| Maltodextrin | 70 | 38 | Mice | 3 × 12 µL | 10 | No | h, c, m | (Debin et al.) [122] |
| Chitosan PLGA | 500 nm–2 µm | - | Cattle | 1/2/3 mL | 10–15 | - | h, m | (Pan et al.) [123] |
| Liposomes | 30–100 | - | Mice | 3 × 50 µL | - | - | c | (Ninomiya et al.) [124] |
| ISCOMs | 40–50 | - | Mice | 2 µL | 2 | Yes | h, m | (Cibulski et al.) [125] |
| Polystyrene | 300–390 | - | Mice | 3 × 20 µL | 10 | - | c | (Misstear et al.) [126] |
| PLA microparticle | - | - | Mice | 10 × 50 µL | - | Yes | h | (Rice-Ficht et al.) [127] |
| Lipopeptide | 150–1000 | - | Mice | 3 × 10 µL | 40 | - | h, m | (Zaman et al.) [128] |
| Dextran | 140–310 | −38:39 | Mice | 3 × 10 µL | 10 | No | h, m | (Marasini et al.) [129] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aziz, T.; Nadeem, A.A.; Sarwar, A.; Perveen, I.; Hussain, N.; Khan, A.A.; Daudzai, Z.; Cui, H.; Lin, L. Particle Nanoarchitectonics for Nanomedicine and Nanotherapeutic Drugs with Special Emphasis on Nasal Drugs and Aging. Biomedicines 2023, 11, 354. https://doi.org/10.3390/biomedicines11020354
Aziz T, Nadeem AA, Sarwar A, Perveen I, Hussain N, Khan AA, Daudzai Z, Cui H, Lin L. Particle Nanoarchitectonics for Nanomedicine and Nanotherapeutic Drugs with Special Emphasis on Nasal Drugs and Aging. Biomedicines. 2023; 11(2):354. https://doi.org/10.3390/biomedicines11020354
Chicago/Turabian StyleAziz, Tariq, Abad Ali Nadeem, Abid Sarwar, Ishrat Perveen, Nageen Hussain, Ayaz Ali Khan, Zubaida Daudzai, Haiying Cui, and Lin Lin. 2023. "Particle Nanoarchitectonics for Nanomedicine and Nanotherapeutic Drugs with Special Emphasis on Nasal Drugs and Aging" Biomedicines 11, no. 2: 354. https://doi.org/10.3390/biomedicines11020354
APA StyleAziz, T., Nadeem, A. A., Sarwar, A., Perveen, I., Hussain, N., Khan, A. A., Daudzai, Z., Cui, H., & Lin, L. (2023). Particle Nanoarchitectonics for Nanomedicine and Nanotherapeutic Drugs with Special Emphasis on Nasal Drugs and Aging. Biomedicines, 11(2), 354. https://doi.org/10.3390/biomedicines11020354