

Preclinical Efficacy of Peripheral Nerve Regeneration by Schwann Cell-like Cells Differentiated from Human Tonsil-Derived Mesenchymal Stem Cells in C22 Mice
 ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cultivation of TMSCs and NRPCs
2.2. Animal Experiments
2.2.1. C22 Mice
2.2.2. Transplantation
2.2.3. Gait Test
2.2.4. Rotarod Test
2.2.5. Nerve Conduction Study
2.3. Transmission Electron Microscopy
2.4. Immunohistochemistry
2.5. Fluorescence In Situ Hybridization
2.6. Real-Time Quantitative Polymerase Chain Reaction
2.7. Statistical Analysis
3. Results
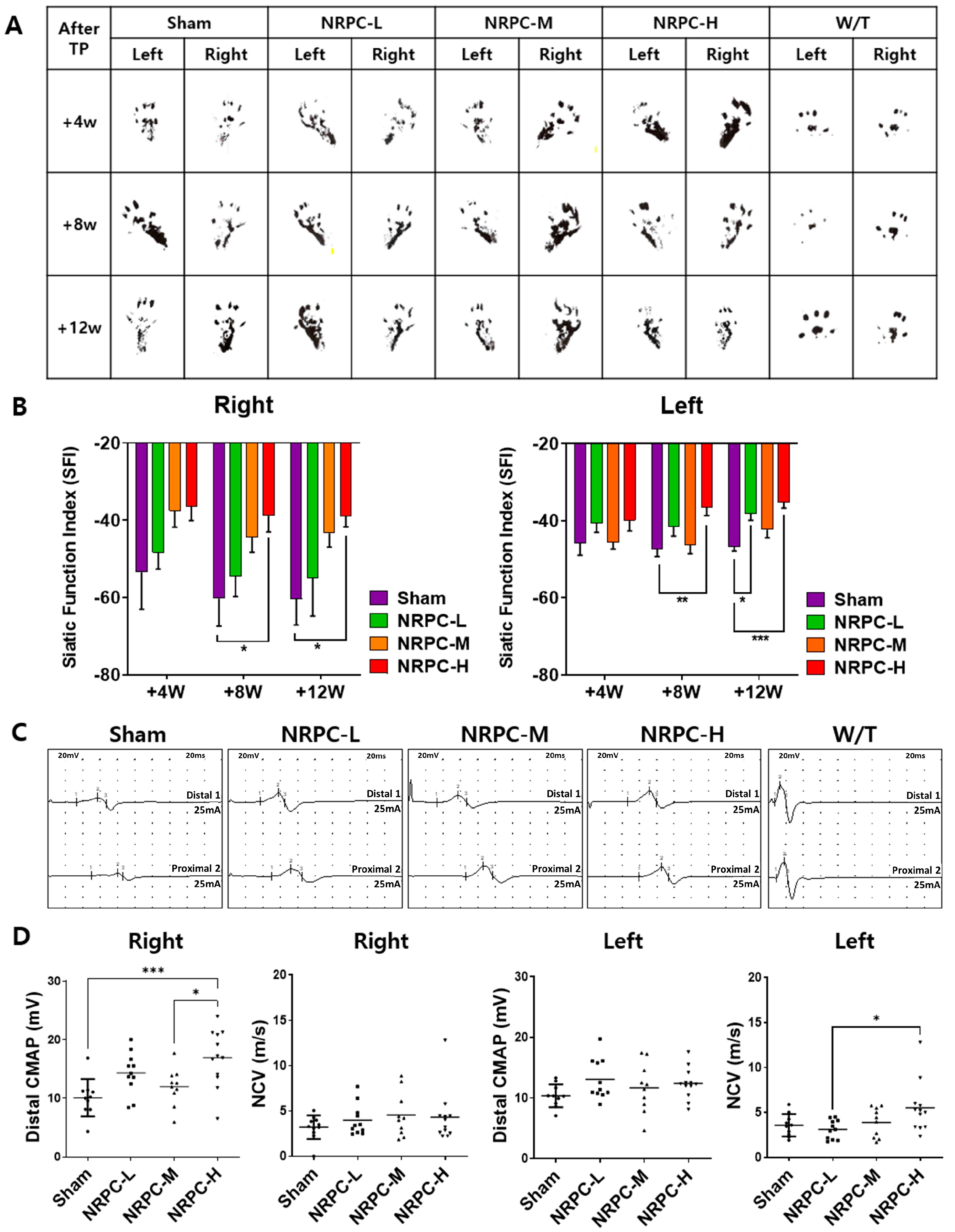
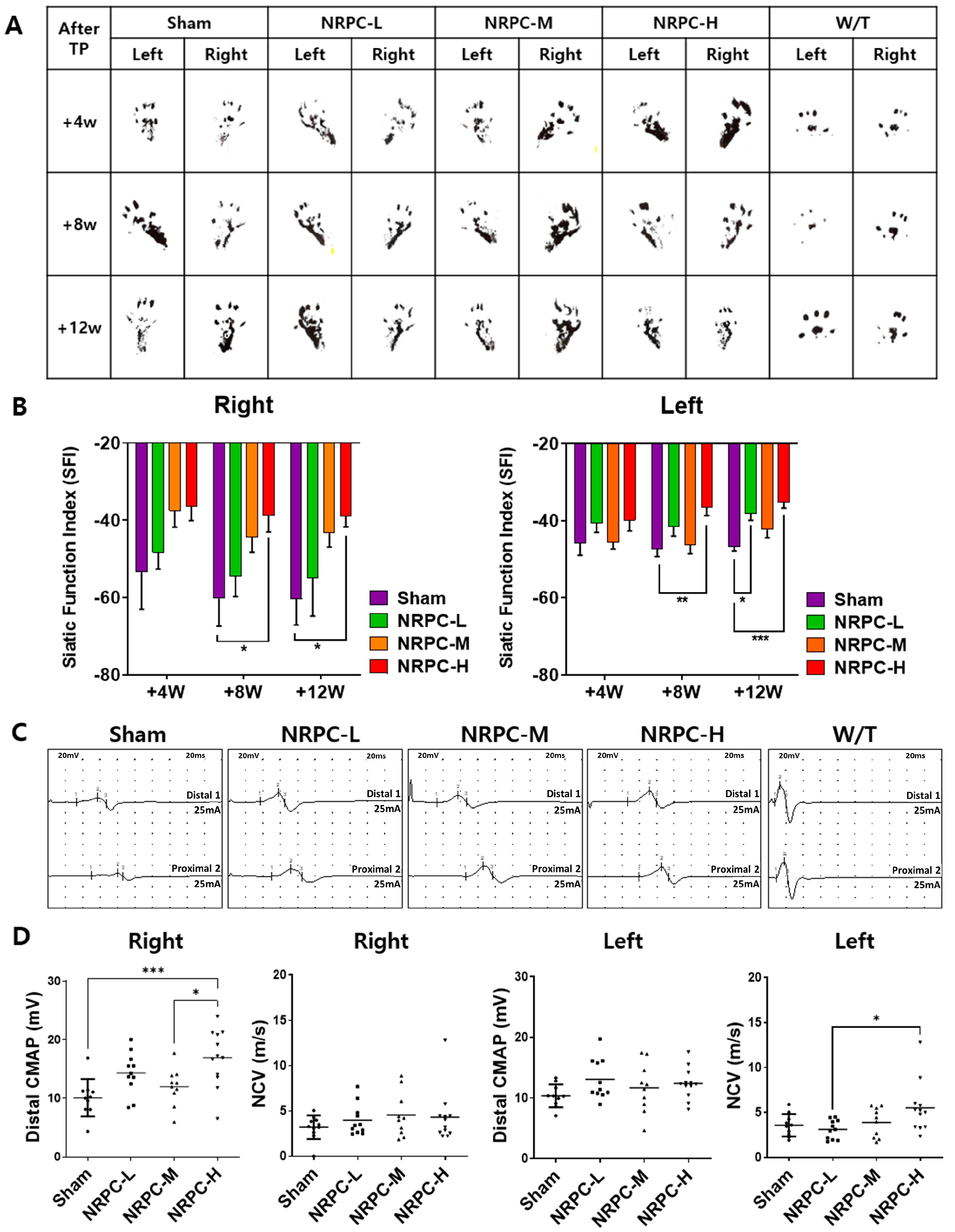
3.1. NRPCs Improved Behavioral Test Results in a Single-Dose Transplantation Experiment
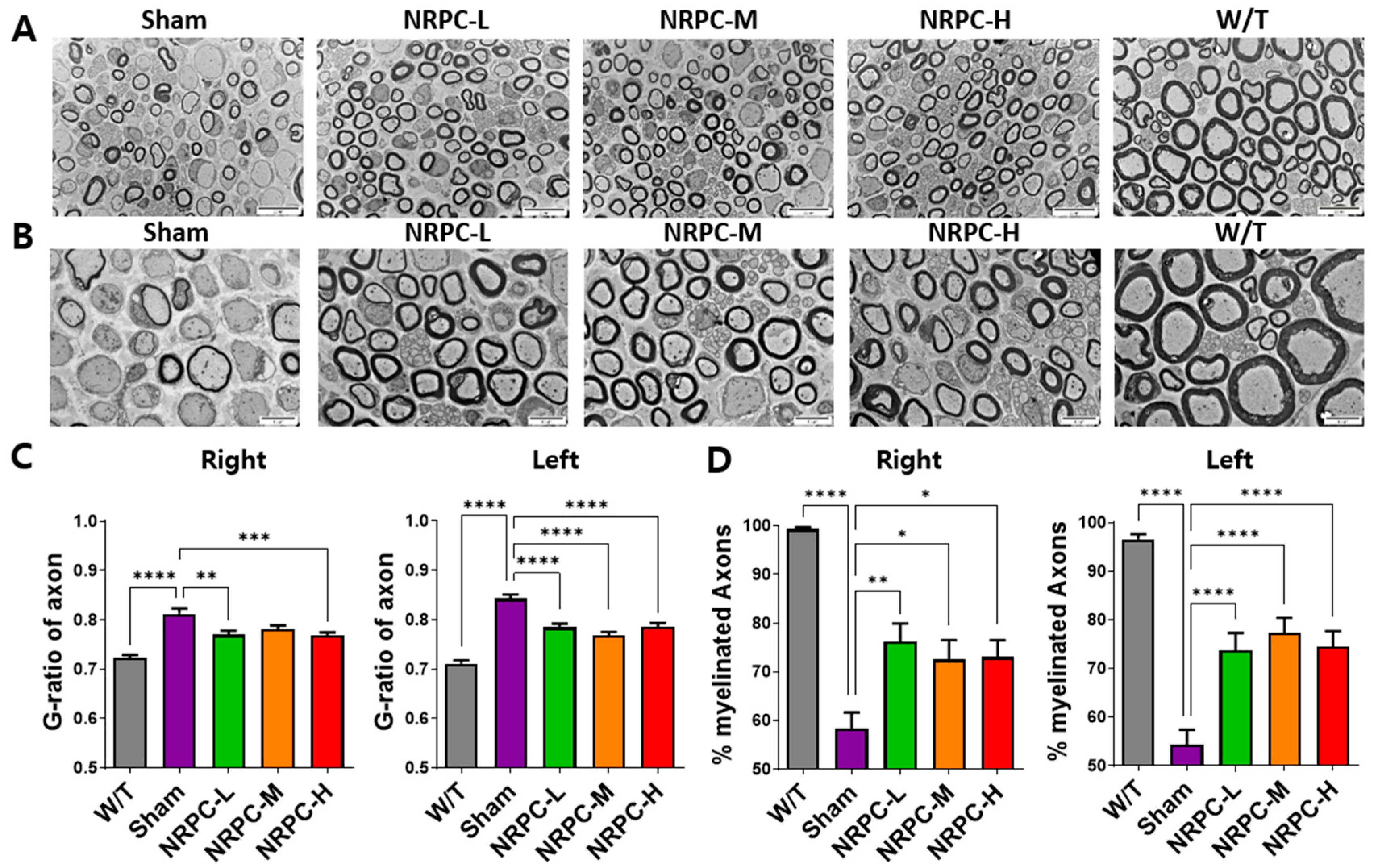
3.2. Restoration of Sciatic Nerve Structure in a Single-Dose Transplantation Experiment
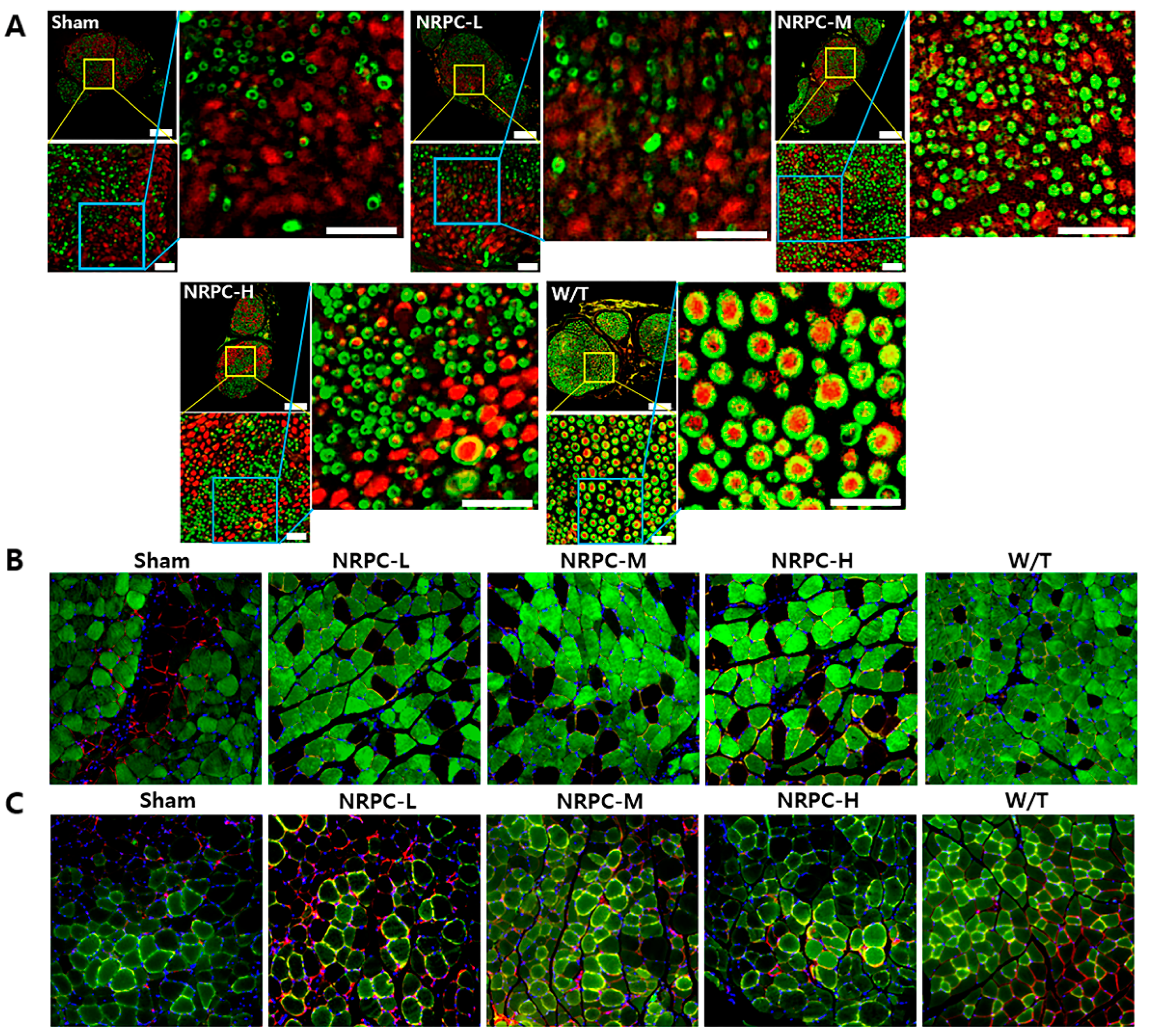
3.3. Regeneration of the Sciatic Nerve and Gastrocnemius and the Related Pathway in a Single-Dose Transplantation Experiment
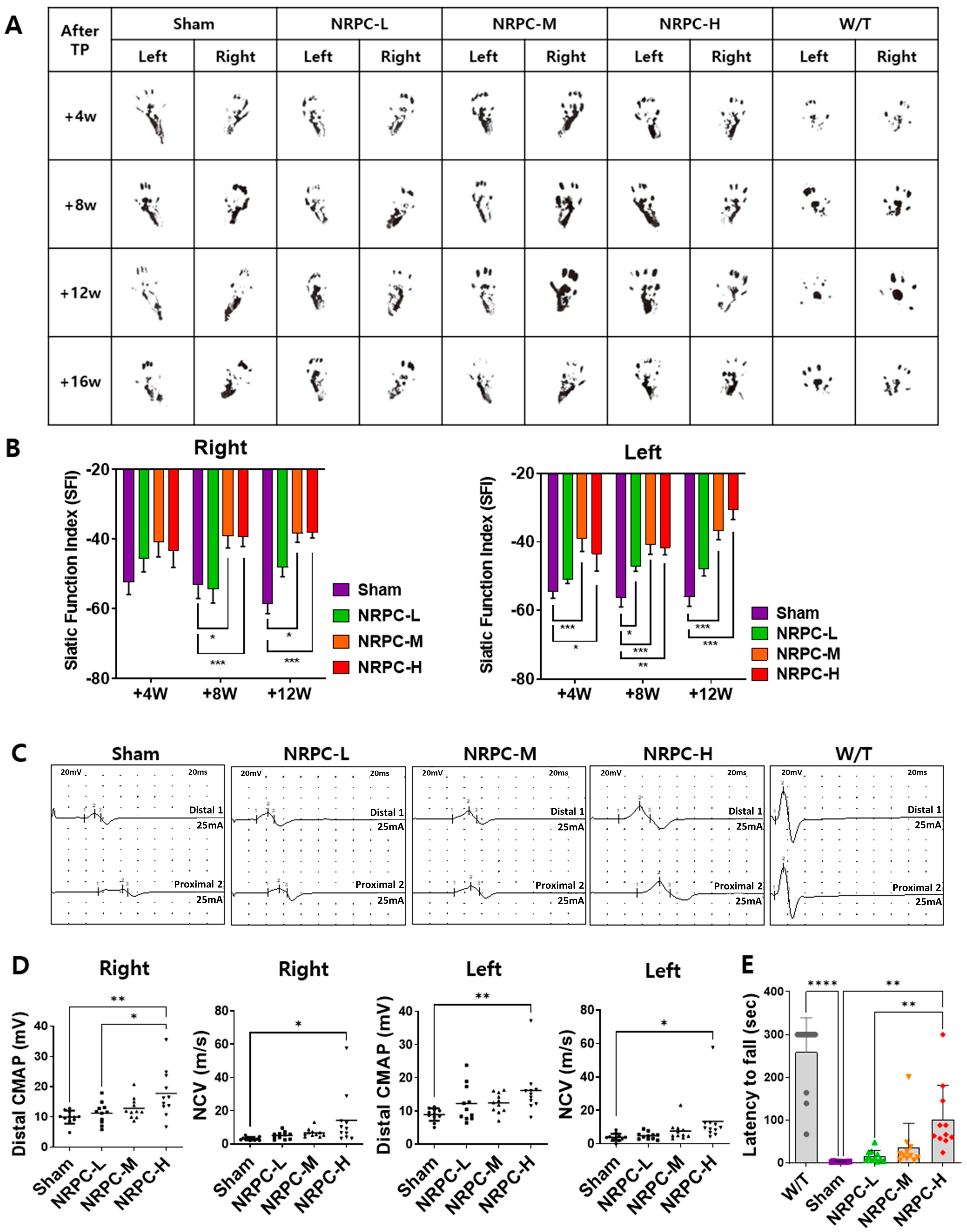
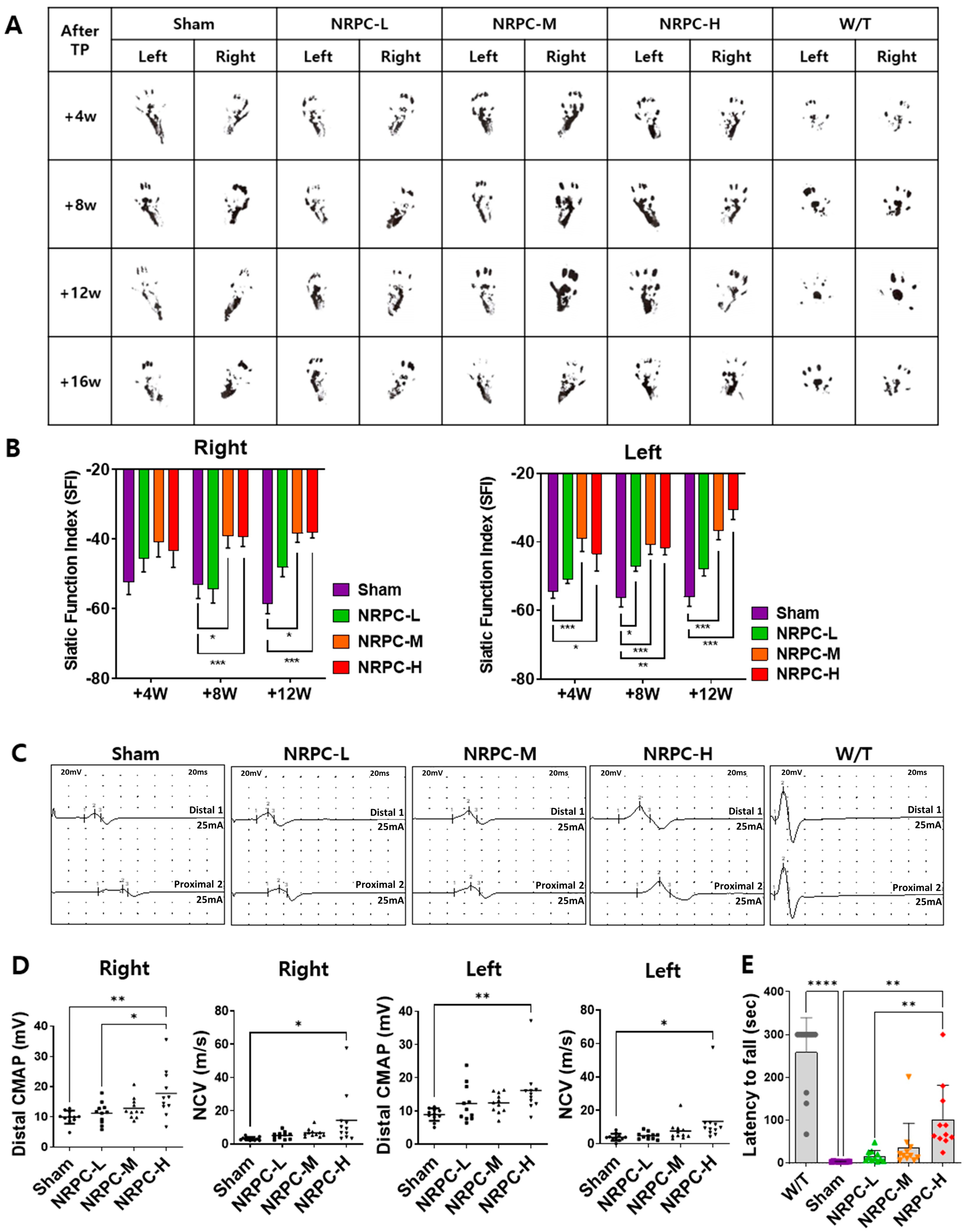
3.4. NRPC Improved Behavioral Test Results in the Repeated-Dose Transplantation Experiment
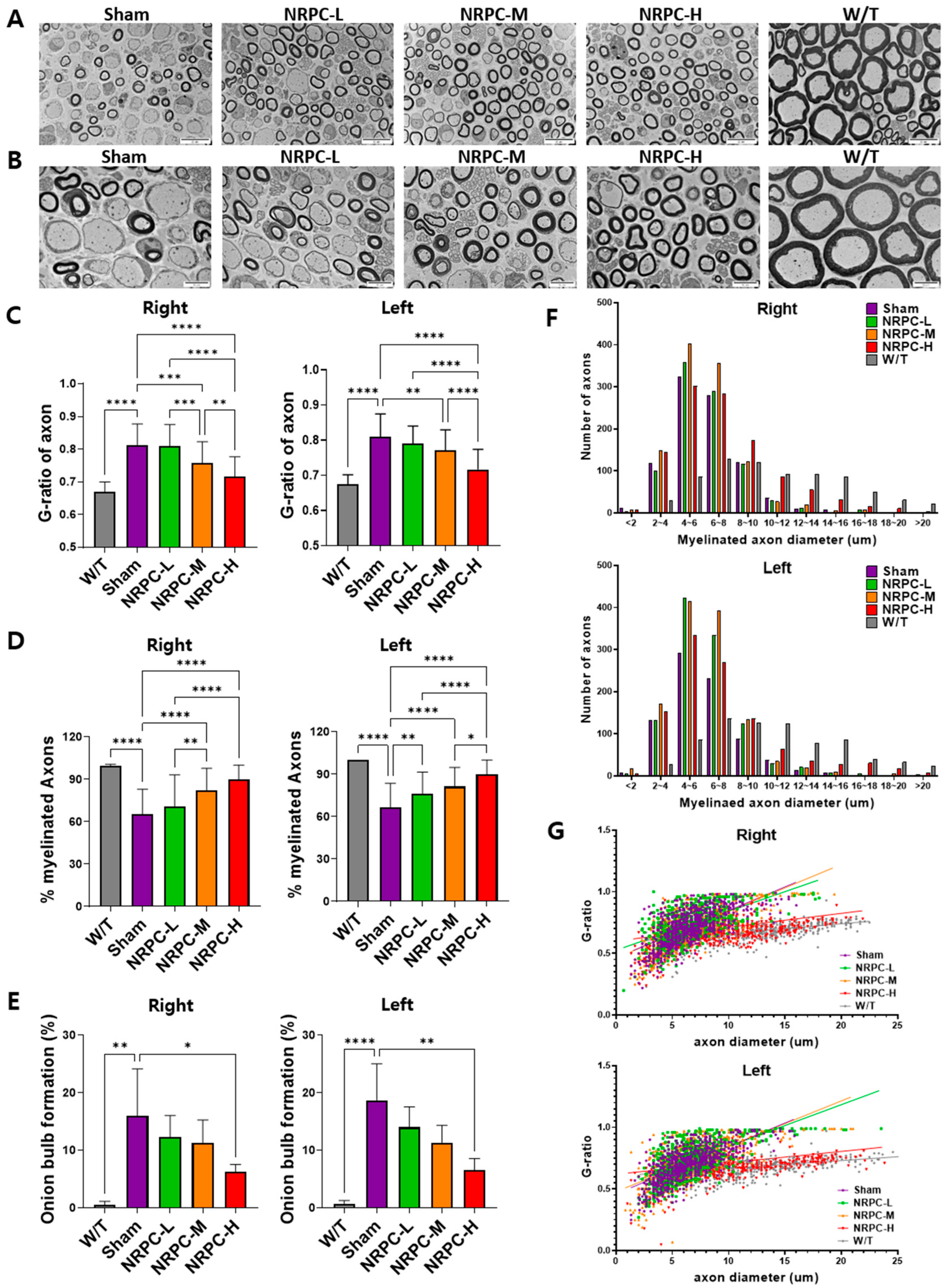
3.5. Restoration of the Sciatic Nerve Structure in the Repeated-Dose Transplantation Experiment
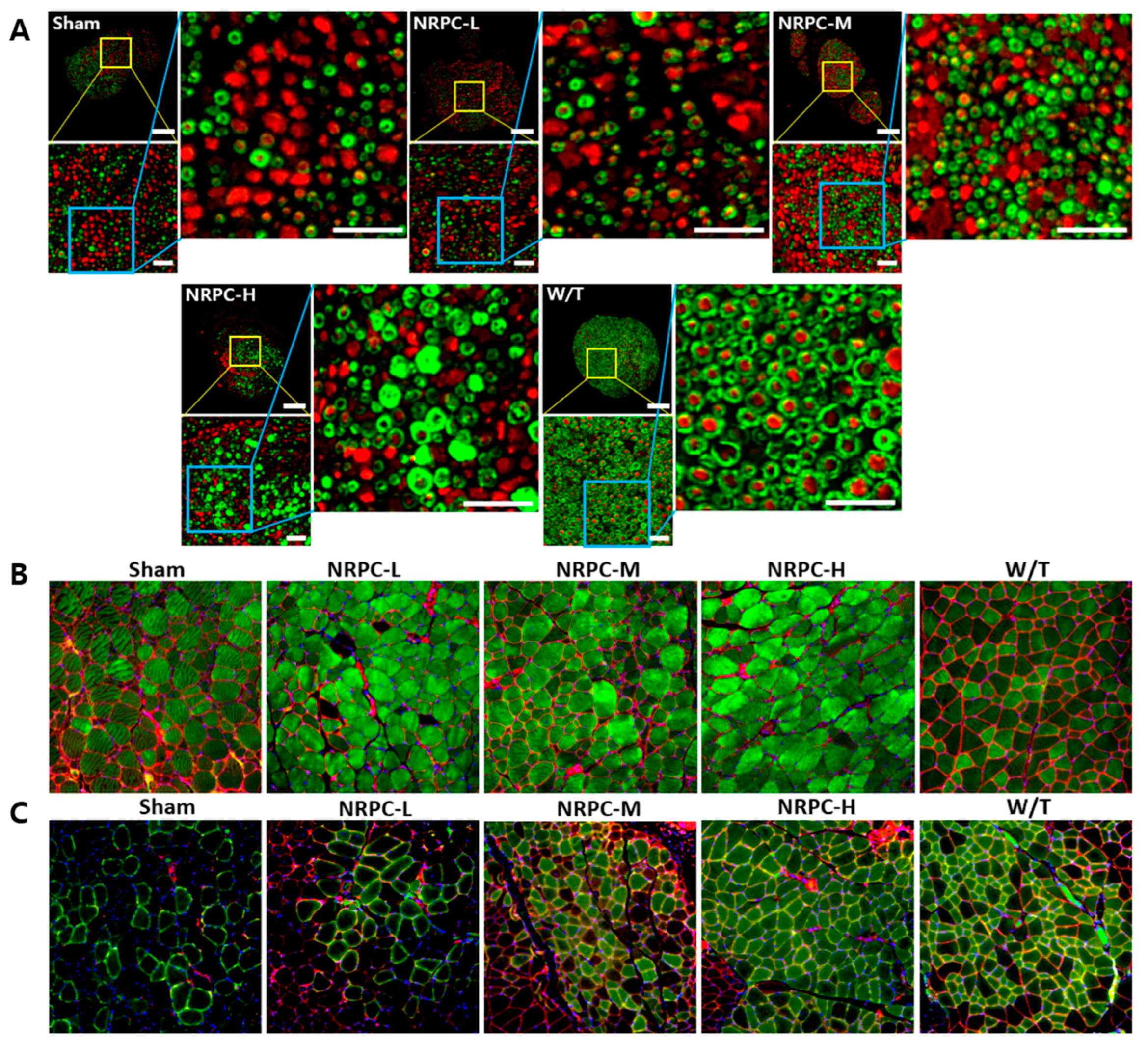
3.6. Restoration of the Sciatic Nerve and Gastrocnemius in the Repeated-Dose Transplantation Experiment
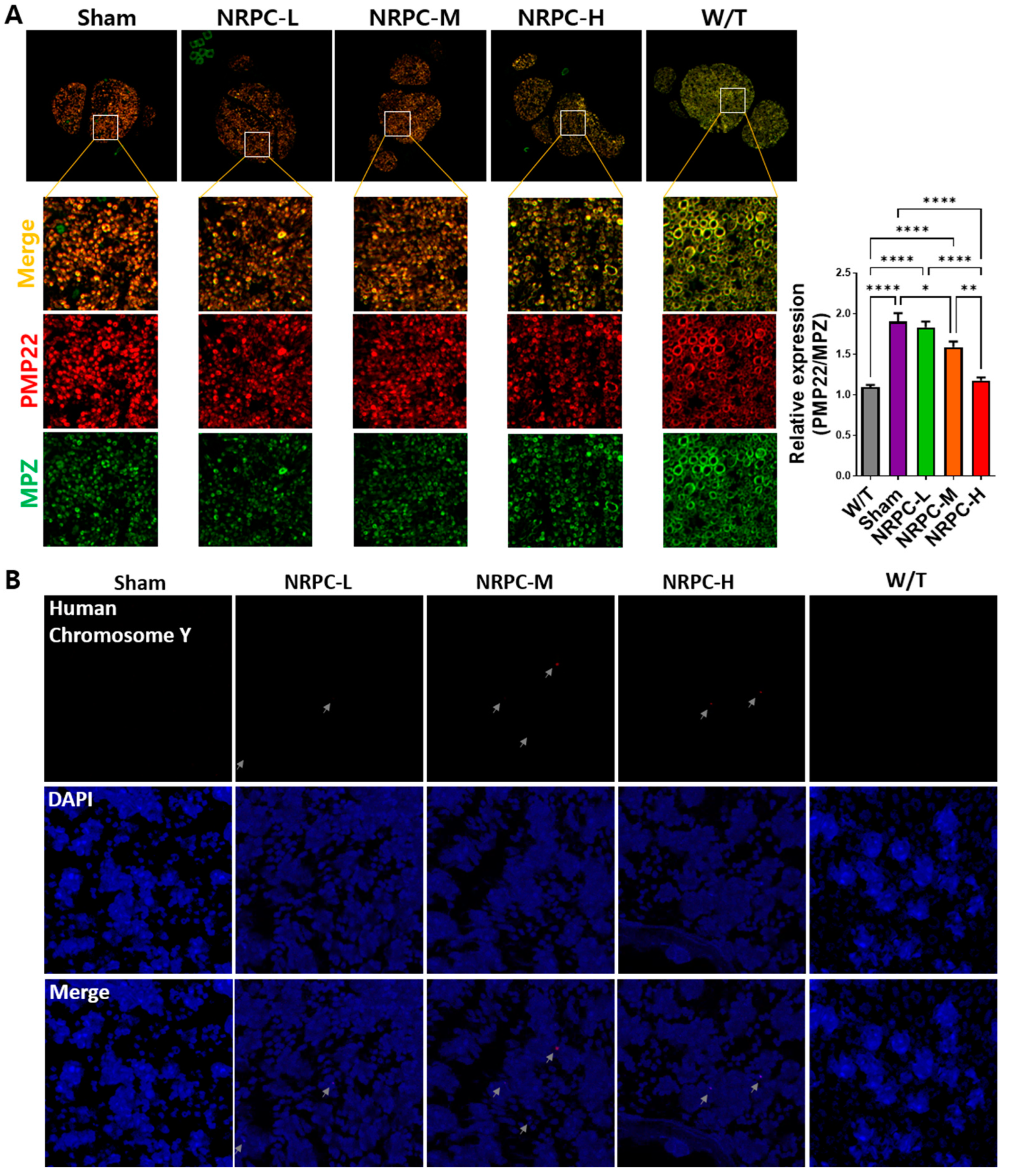
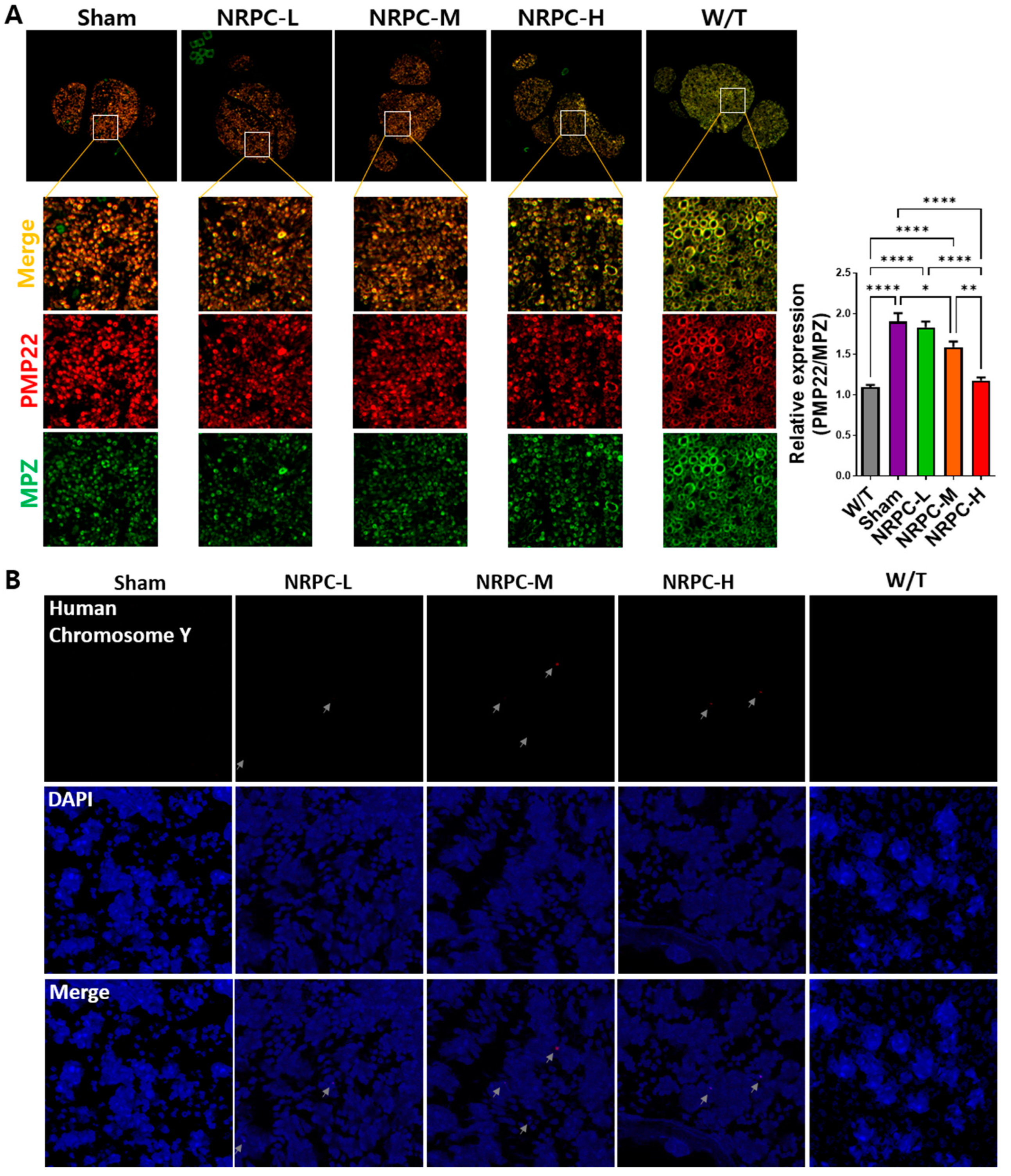
3.7. Downregulation of PMP22 and the Presence of NRPCs 12 Weeks after Repeated-Dose Transplantation in the Sciatic Nerve
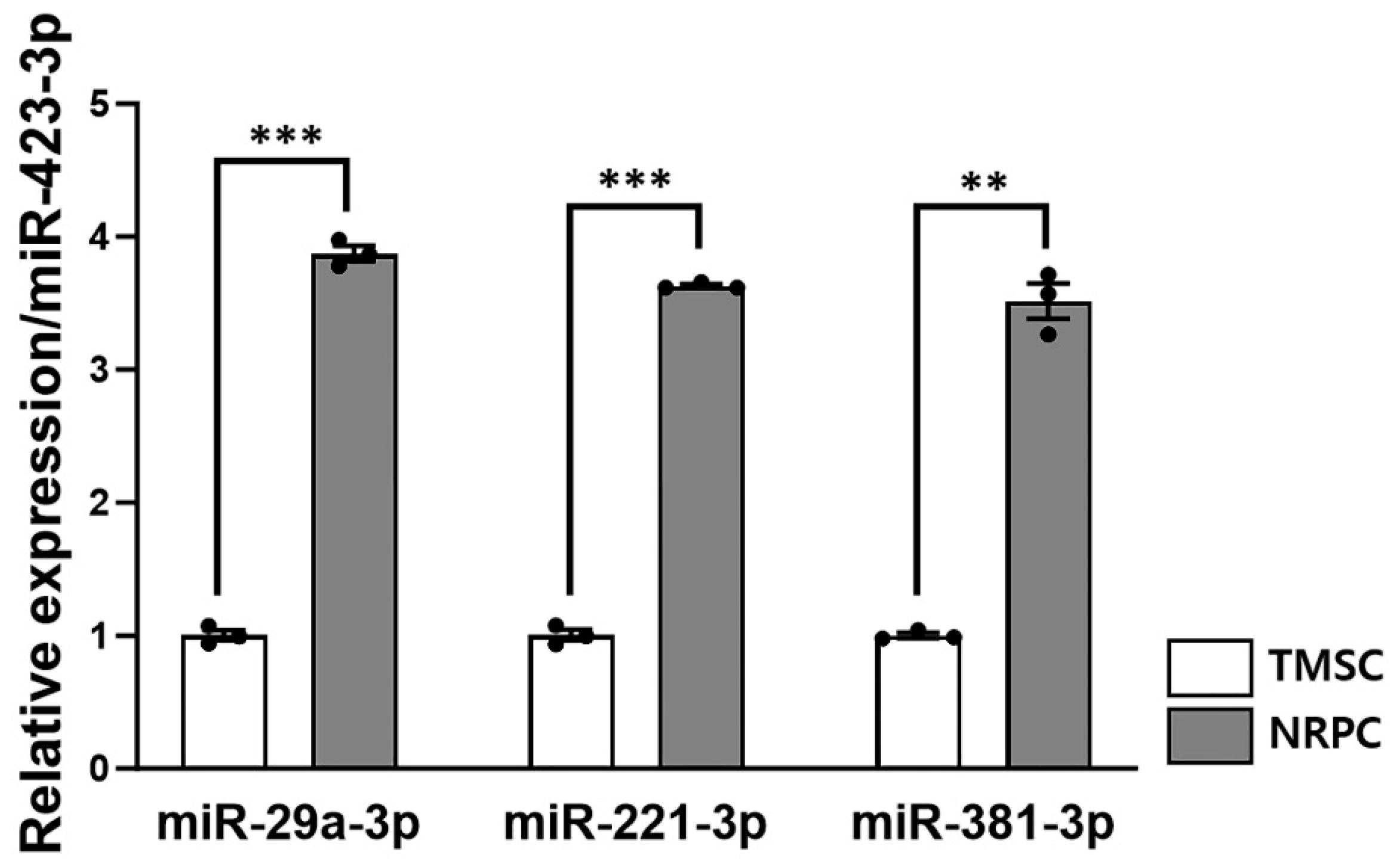
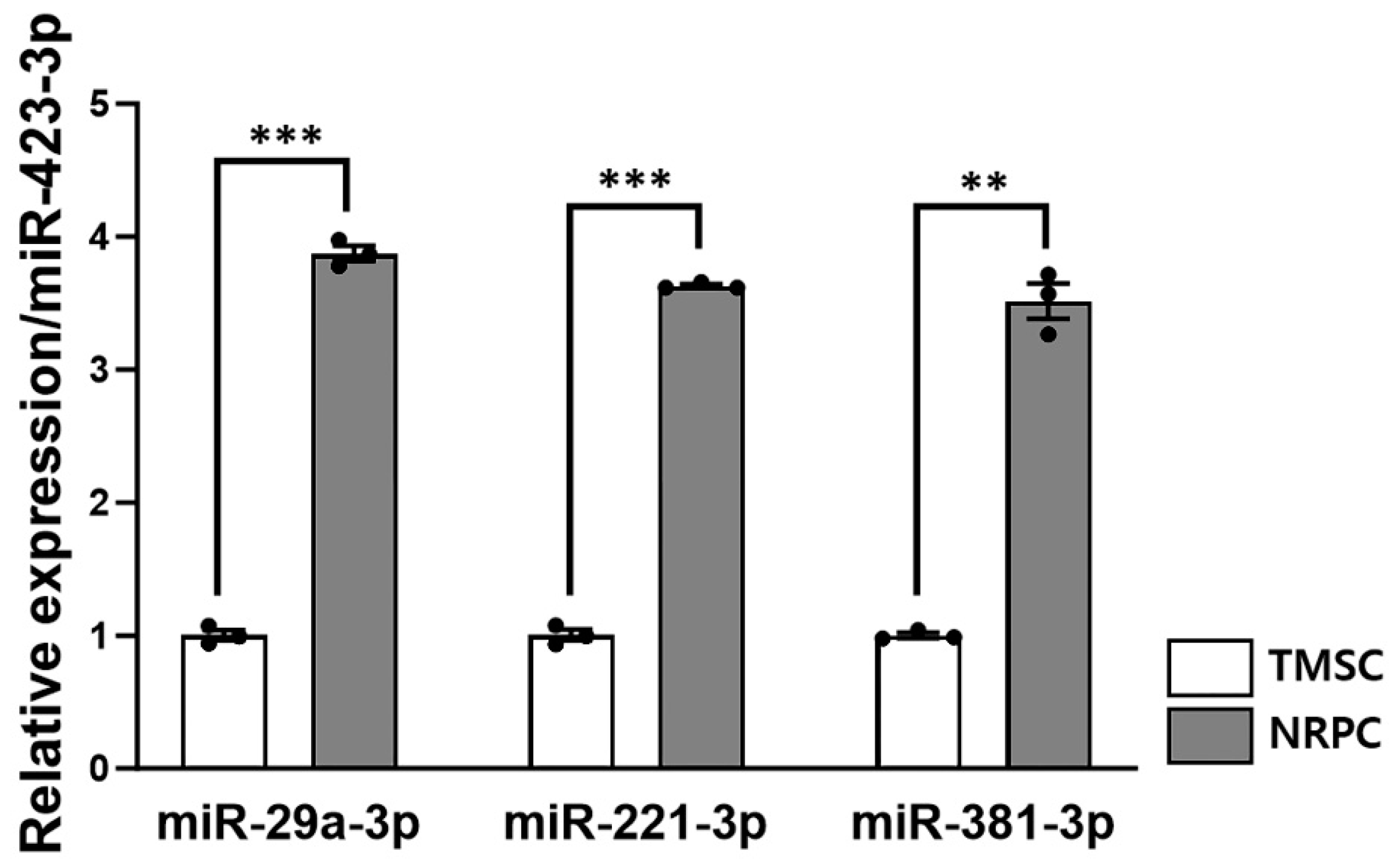
3.8. Expression of Peripheral Nerve Regeneration-Related microRNA in NRPCs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Okamoto, Y.; Takashima, H. The Current State of Charcot-Marie-Tooth Disease Treatment. Genes 2023, 14, 1391. [Google Scholar] [CrossRef] [PubMed]
- Stavrou, M.; Kagiava, A.; Sargiannidou, I.; Georgiou, E.; Kleopa, K.A. Charcot-Marie-Tooth neuropathies: Current gene therapy advances and the route toward translation. J. Peripher. Nerv. Syst. 2023, 28, 150–168. [Google Scholar] [CrossRef] [PubMed]
- Health Information on National Institute of Neurological Disorders and Stroke Website. Available online: https://www.ninds.nih.gov/health-information/disorders/charcot-marie-tooth-disease (accessed on 5 October 2023).
- Roa, B.B.; Dyck, P.J.; Marks, H.G.; Chance, P.F.; Lupski, J.R. Dejerine–Sottas Syndrome Associated with Point Mutation in the Peripheral Myelin Protein 22 (PMP22) Gene. Nat. Genet. 1993, 5, 269–273. [Google Scholar] [CrossRef] [PubMed]
- Boutary, S.; Echaniz-Laguna, A.; Adams, D.; Loisel-Duwattez, J.; Schumacher, M.; Massaad, C.; Massaad-Massade, L. Treating PMP22 Gene Duplication-Related Charcot-Marie-Tooth Disease: The Past, the Present and the Future. Transl. Res. 2021, 227, 100–111. [Google Scholar] [CrossRef]
- Mittendorf, K.F.; Marinko, J.T.; Hampton, C.M.; Ke, Z.; Hadziselimovic, A.; Schlebach, J.P.; Law, C.L.; Li, J.; Wright, E.R.; Sanders, C.R.; et al. Peripheral myelin protein 22 alters membrane architecture. Sci. Adv. 2017, 3, e1700220. [Google Scholar] [CrossRef]
- Prukop, T.; Stenzel, J.; Wernick, S.; Kungl, T.; Mroczek, M.; Adam, J.; Ewers, D.; Nabirotchkin, S.; Nave, K.-A.; Hajj, R.; et al. Early Short-Term PXT3003 Combinational Therapy Delays Disease Onset in a Transgenic Rat Model of Charcot-Marie-Tooth Disease 1A (CMT1A). PLoS ONE 2019, 14, e0209752. [Google Scholar] [CrossRef]
- Ha, N.; Choi, Y.I.; Jung, N.; Song, J.Y.; Bae, D.K.; Kim, M.C.; Lee, Y.J.; Song, H.; Kwak, G.; Jeong, S.; et al. A Novel Histone Deacetylase 6 Inhibitor Improves Myelination of Schwann Cells in a Model of Charcot–Marie–Tooth Disease Type 1A. Br. J. Pharmacol. 2020, 177, 5096–5113. [Google Scholar] [CrossRef]
- Jeon, H.; Kim, H.J.; Doo, H.M.; Chang, E.H.; Kwak, G.; Mo, W.M.; Jang, S.Y.; Lee, M.W.; Choi, B.-O.; Hong, Y.B. Cytokines Secreted by Mesenchymal Stem Cells Reduce Demyelination in an Animal Model of Charcot-Marie-Tooth Disease. Biochem. Biophys. Res. Commun. 2022, 597, 1–7. [Google Scholar] [CrossRef]
- Ozes, B.; Myers, M.; Moss, K.; Mckinney, J.; Ridgley, A.; Chen, L.; Bai, S.; Abrams, C.K.; Freidin, M.M.; Mendell, J.R.; et al. AAV1.NT-3 Gene Therapy for X-Linked Charcot–Marie–Tooth Neuropathy Type 1. Gene Ther. 2022, 29, 127–137. [Google Scholar] [CrossRef]
- Zhao, H.T.; Damle, S.; Ikeda-Lee, K.; Kuntz, S.; Li, J.; Mohan, A.; Kim, A.; Hung, G.; Scheideler, M.A.; Scherer, S.S.; et al. PMP22 Antisense Oligonucleotides Reverse Charcot-Marie-Tooth Disease Type 1A Features in Rodent Models. J. Clin. Investig. 2018, 128, 359–368. [Google Scholar] [CrossRef]
- Gautier, B.; Hajjar, H.; Soares, S.; Berthelot, J.; Deck, M.; Abbou, S.; Campbell, G.; Ceprian, M.; Gonzalez, S.; Fovet, C.-M.; et al. AAV2/9-Mediated Silencing of PMP22 Prevents the Development of Pathological Features in a Rat Model of Charcot-Marie-Tooth Disease 1 A. Nat. Commun. 2021, 12, 2356. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-S.; Lee, J.Y.; Song, D.W.; Bae, H.S.; Doo, H.M.; Yu, H.S.; Lee, K.J.; Kim, H.K.; Hwang, H.; Kwak, G.; et al. Targeted PMP22 TATA-Box Editing by CRISPR/Cas9 Reduces Demyelinating Neuropathy of Charcot-Marie-Tooth Disease Type 1A in Mice. Nucleic Acids Res. 2020, 48, 130–140. [Google Scholar] [CrossRef]
- Fernández-Garza, L.E.; Barrera-Barrera, S.A.; Barrera-Saldaña, H.A. Mesenchymal Stem Cell Therapies Approved by Regulatory Agencies around the World. Pharmaceuticals 2023, 16, 1334. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.Y.; Choi, Y.M.; Kim, H.Y.; Park, Y.S.; Jung, S.C.; Park, J.W.; Woo, S.Y.; Ryu, K.H.; Kim, H.S.; Jo, I. Application of tonsil-derived mesenchymal stem cells in tissue regeneration: Concise review. Stem. Cells 2019, 37, 1252–1260. [Google Scholar] [CrossRef] [PubMed]
- Ryu, K.-H.; Cho, K.-A.; Park, H.S.; Kim, J.-Y.; Woo, S.-Y.; Jo, I.; Choi, Y.H.; Park, Y.M.; Jung, S.-C.; Chung, S.M.; et al. Tonsil-Derived Mesenchymal Stromal Cells: Evaluation of Biologic, Immunologic and Genetic Factors for Successful Banking. Cytotherapy 2012, 14, 1193–1202. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Jeong, S.; Nam, Y.H.; Yum, Y.; Jung, S.-C. Transplantation of Differentiated Tonsil-Derived Mesenchymal Stem Cells Ameliorates Murine Duchenne Muscular Dystrophy via Autophagy Activation. Tissue Eng. Regen. Med. 2022, 19, 1283–1294. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Park, S.; Oh, S.-Y.; Nam, Y.H.; Choi, Y.M.; Choi, Y.; Kim, H.Y.; Jung, S.Y.; Kim, H.S.; Jo, I.; et al. Density-Dependent Differentiation of Tonsil-Derived Mesenchymal Stem Cells into Parathyroid-Hormone-Releasing Cells. Int. J. Mol. Sci. 2022, 23, 715. [Google Scholar] [CrossRef]
- Park, S.; Kim, J.Y.; Myung, S.; Jung, N.; Choi, Y.; Jung, S.C. Differentiation of Motor Neuron-Like Cells from Tonsil-Derived Mesenchymal Stem Cells and Their Possible Application to Neuromuscular Junction Formation. Int. J. Mol. Sci. 2019, 20, 2702. [Google Scholar] [CrossRef]
- Park, S.; Jung, N.; Myung, S.; Choi, Y.; Chung, K.W.; Choi, B.-O.; Jung, S.-C. Differentiation of Human Tonsil-Derived Mesenchymal Stem Cells into Schwann-Like Cells Improves Neuromuscular Function in a Mouse Model of Charcot-Marie-Tooth Disease Type 1A. Int. J. Mol. Sci. 2018, 19, 2393. [Google Scholar] [CrossRef]
- Balakrishnan, A.; Belfiore, L.; Chu, T.H.; Fleming, T.; Midha, R.; Biernaskie, J.; Schuurmans, C. Insights into the Role and Potential of Schwann Cells for Peripheral Nerve Repair from Studies of Development and Injury. Front Mol. Neurosci. 2021, 13, 608442. [Google Scholar] [CrossRef]
- Bolívar, S.; Navarro, X.; Udina, E. Schwann Cell Role in Selectivity of Nerve Regeneration. Cells 2020, 9, 2131. [Google Scholar] [CrossRef] [PubMed]
- Hopf, A.; Schaefer, D.J.; Kalbermatten, D.F.; Guzman, R.; Madduri, S. Schwann Cell-like Cells: Origin and Usability for Repair and Regeneration of the Peripheral and Central Nervous System. Cells 2020, 9, 1990. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Men, Y.; Dong, P. Schwann Cells Promote the Capability of Neural Stem Cells to Differentiate into Neurons and Secret Neurotrophic Factors. Exp. Ther. Med. 2017, 13, 2029–2035. [Google Scholar] [CrossRef] [PubMed]
- Jung, N.; Park, S.; Choi, Y.; Park, J.W.; Hong, Y.B.; Park, H.H.; Yu, Y.; Kwak, G.; Kim, H.S.; Ryu, K.H.; et al. Tonsil-Derived Mesenchymal Stem Cells Differentiate into a Schwann Cell Phenotype and Promote Peripheral Nerve Regeneration. Int. J. Mol. Sci. 2016, 17, 1867. [Google Scholar] [CrossRef] [PubMed]
- Fricker, L.; Penna, V.; Lampert, F.; Stark, G.B.; Witzel, C.; Koulaxouzidis, G. A Self-Made, Low-Cost Infrared System for Evaluating the Sciatic Functional Index in Mice. Neural Regen. Res. 2016, 11, 829–834. [Google Scholar] [CrossRef]
- Pisciotta, C.; Saveri, P.; Pareyson, D. Challenges in Treating Charcot-Marie-Tooth Disease and Related Neuropathies: Current Management and Future Perspectives. Brain Sci. 2021, 11, 1447. [Google Scholar] [CrossRef]
- Boutary, S.; Caillaud, M.; El Madani, M.; Vallat, J.-M.; Loisel-Duwattez, J.; Rouyer, A.; Richard, L.; Gracia, C.; Urbinati, G.; Desmaële, D.; et al. Squalenoyl SiRNA PMP22 Nanoparticles Are Effective in Treating Mouse Models of Charcot-Marie-Tooth Disease Type 1 A. Commun. Biol. 2021, 4, 317. [Google Scholar] [CrossRef]
- Stavrou, M.; Kagiava, A.; Choudury, S.G.; Jennings, M.J.; Wallace, L.M.; Fowler, A.M.; Heslegrave, A.; Richter, J.; Tryfonos, C.; Christodoulou, C.; et al. A Translatable RNAi-Driven Gene Therapy Silences PMP22/Pmp22 Genes and Improves Neuropathy in CMT1A Mice. J. Clin. Investig. 2022, 132, e159814. [Google Scholar] [CrossRef]
- Robertson, A.M.; Perea, J.; McGuigan, A.; King, R.H.M.; Muddle, J.R.; Gabreëls-Festen, A.A.; Thomas, P.K.; Huxley, C. Comparison of a New Pmp22 Transgenic Mouse Line with Other Mouse Models and Human Patients with CMT1A*. J. Anat. 2002, 200, 377–390. [Google Scholar] [CrossRef]
- Fledrich, R.; Stassart, R.M.; Sereda, M.W. Murine Therapeutic Models for Charcot-Marie-Tooth (CMT) Disease. Br. Med. Bull. 2012, 102, 89–113. [Google Scholar] [CrossRef]
- Robertson, A.M.; Huxley, C.; King, R.H.M.; Thomas, P.K. Development of Early Postnatal Peripheral Nerve Abnormalities in Trembler-J and PMP22 Transgenic Mice. J. Anat. 1999, 195, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Duman, M.; Jaggi, S.; Enz, L.S.; Jacob, C.; Schaeren-Wiemers, N. Theophylline Induces Remyelination and Functional Recovery in a Mouse Model of Peripheral Neuropathy. Biomedicines 2022, 10, 1418. [Google Scholar] [CrossRef] [PubMed]
- Walsh, S.; Midha, R. Practical Considerations Concerning the Use of Stem Cells for Peripheral Nerve Repair. Neurosurg. Focus 2009, 26, E2. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-S.; Kwak, G.; Kim, H.J.; Park, H.-T.; Choi, B.-O.; Hong, Y.B. MiR-381 Attenuates Peripheral Neuropathic Phenotype Caused by Overexpression of PMP22. Exp. Neurobiol. 2019, 28, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Serfecz, J.; Bazick, H.; Al Salihi, M.O.; Turner, P.; Fields, C.; Cruz, P.; Renne, R.; Notterpek, L. Downregulation of the Human Peripheral Myelin Protein 22 Gene by MiR-29a in Cellular Models of Charcot–Marie–Tooth Disease. Gene Ther. 2019, 26, 455–464. [Google Scholar] [CrossRef] [PubMed]
- Verrier, J.D.; Lau, P.; Hudson, L.; Murashov, A.K.; Renne, R.; Notterpek, L. Peripheral Myelin Protein 22 Is Regulated Post-Transcriptionally by MiRNA-29a. Glia 2009, 57, 1265–1279. [Google Scholar] [CrossRef]
- Wen, L.-L.; Yu, T.-H.; Ma, Y.-Z.; Mao, X.-Y.; Ao, T.-R.; Javed, R.; Ten, H.; Matsuno, A.; Ao, Q. Sequential Expression of MiR-221-3p and MiR-338-3p in Schwann Cells as a Therapeutic Strategy to Promote Nerve Regeneration and Functional Recovery. Neural Regen. Res. 2022, 18, 671–682. [Google Scholar] [CrossRef]
- Ma, C.H.E.; Omura, T.; Cobos, E.J.; Latrémolière, A.; Ghasemlou, N.; Brenner, G.J.; van Veen, E.; Barrett, L.; Sawada, T.; Gao, F.; et al. Accelerating Axonal GroW/Th Promotes Motor Recovery after Peripheral Nerve Injury in Mice. J. Clin. Investig. 2011, 121, 4332–4347. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Value/Specification |
|---|---|
| Viability | >90% |
| Morphology | Specific form for neural precursor cell of the shape of a bipolar or tripolar |
| Positive markers | CD73, CD90, CD105 |
| Negative markers | CD14, CD34, CD45 |
| Infectious agent test | SCB (p4), WCB (p9), F/P (p14) |
| Other quality tests | HIV RNA, HCV RNA, HBV DNA, CMV RNA, anti-HIV, anti-HCV, anti-HBsAg, anti-CMV IgM/IgG, anti-HTLV-1/2, Syphilis |
| Cryoprotectant | Cryostor CS10 |
| Experiment | Group | Number of Cells for Transplantation | Mouse | Number of Animals (Male) | Route of Administration | Dosing Frequency |
|---|---|---|---|---|---|---|
| Single administration | NRPC-low | 2.5 × 104 cells/ 100 μL/site × 2 sites | C22 mice | 11 |
| Single |
| NRPC-med | 2.5 × 105 cells/ 100 μL/site × 2 sites | 11 | ||||
| NRPC-high | 5.0 × 105 cells/ 100 μL/site × 2 sites | 12 | ||||
| Sham | CS10/100 μL/ site × 2 sites | 11 | ||||
| W/T | - | Wild-type mice | 12 | |||
| Repeated administration | NRPC-low | 2.5 × 104 cells/ 100 μL/site × 2 sites | C22 mice | 11 |
| Two repetitions 4 weeks apart |
| NRPC-med | 2.5 × 105 cells/ 100 μL/site × 2 sites | 11 | ||||
| NRPC-high | 5.0 × 105 cells/ 100 μL/site × 2 sites | 11 | ||||
| Sham | CS10/100 μL/ site × 2 sites | 11 | ||||
| W/T | - | Wild-type mice | 13 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nam, Y.H.; Park, S.; Yum, Y.; Jeong, S.; Park, H.E.; Kim, H.J.; Lim, J.; Choi, B.-O.; Jung, S.-C. Preclinical Efficacy of Peripheral Nerve Regeneration by Schwann Cell-like Cells Differentiated from Human Tonsil-Derived Mesenchymal Stem Cells in C22 Mice. Biomedicines 2023, 11, 3334. https://doi.org/10.3390/biomedicines11123334
Nam YH, Park S, Yum Y, Jeong S, Park HE, Kim HJ, Lim J, Choi B-O, Jung S-C. Preclinical Efficacy of Peripheral Nerve Regeneration by Schwann Cell-like Cells Differentiated from Human Tonsil-Derived Mesenchymal Stem Cells in C22 Mice. Biomedicines. 2023; 11(12):3334. https://doi.org/10.3390/biomedicines11123334
Chicago/Turabian StyleNam, Yu Hwa, Saeyoung Park, Yoonji Yum, Soyeon Jeong, Hyo Eun Park, Ho Jin Kim, Jaeseung Lim, Byung-Ok Choi, and Sung-Chul Jung. 2023. "Preclinical Efficacy of Peripheral Nerve Regeneration by Schwann Cell-like Cells Differentiated from Human Tonsil-Derived Mesenchymal Stem Cells in C22 Mice" Biomedicines 11, no. 12: 3334. https://doi.org/10.3390/biomedicines11123334
APA StyleNam, Y. H., Park, S., Yum, Y., Jeong, S., Park, H. E., Kim, H. J., Lim, J., Choi, B.-O., & Jung, S.-C. (2023). Preclinical Efficacy of Peripheral Nerve Regeneration by Schwann Cell-like Cells Differentiated from Human Tonsil-Derived Mesenchymal Stem Cells in C22 Mice. Biomedicines, 11(12), 3334. https://doi.org/10.3390/biomedicines11123334