
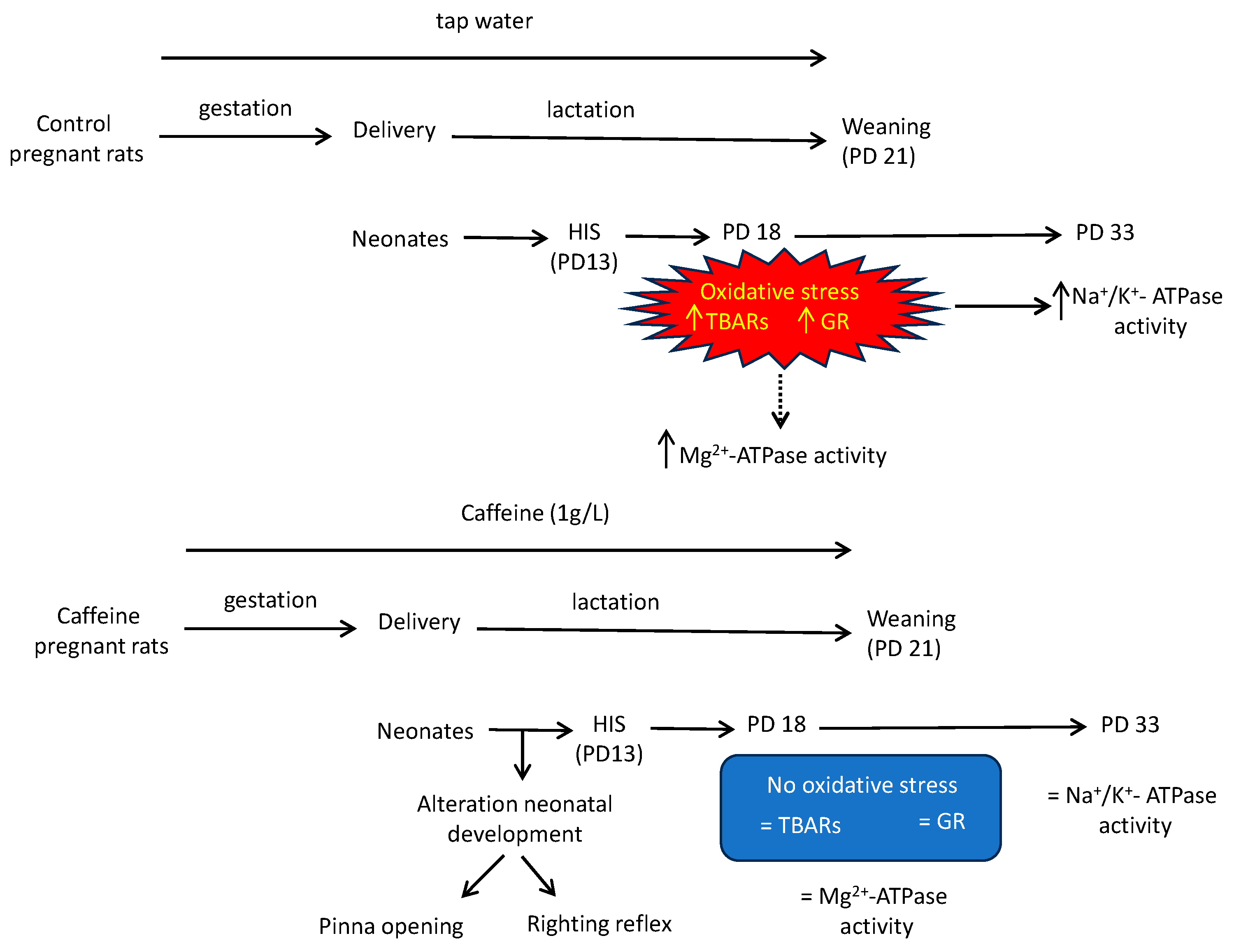
Maternal Caffeine Consumption during Gestation and Lactation Abolishes Cortical Oxidative Stress and Restores Na+/K+-ATPase Activity in Neonates Exposed to Hyperthermia-Induced Seizures


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animals
- (a)
- Water control rats: these animals were not exposed to HIS, and their mothers drank water during gestation and lactation;
- (b)
- Water HIS rats: these animals were exposed to hyperthermic seizures, and their mothers drank water during gestation and lactation;
- (c)
- Caffeine control rats: these animals were not exposed to HIS, and their mothers drank caffeine during gestation and lactation;
- (d)
- Caffeine HIS rats: these animals were exposed to HIS, and their mothers drank caffeine during gestation and lactation.
2.3. Hyperthermia-Induced Seizures
2.4. Preparation of Plasma Membranes and Cytosolic Fractions from Rat Cortex
2.5. Neonatal Development
- (a)
- Auricle opening: the postnatal day when the auricles were fully opened was recorded;
- (b)
- Righting reflex on the surface: pups were placed face up on a smooth surface, and the time it took for them to assume a normal position was noted. The animals were allowed 15 s to turn over;
- (c)
- Geotaxis reflex: pups were positioned on an inclined surface with their heads facing downward, and the time required for them to reorient and face upward was recorded. The animals were given 30 s to perform the task;
- (d)
- Eye opening: the day when the pups first opened their eyes was recorded.
2.6. Lipid Peroxidation Levels Determinations in the Cortex of Rats Submitted to Hyperthermia-Induced Seizures
- (a)
- Five days after HIS: control water (n = 3); HIS water (n = 4); control caffeine (n = 5); and HIS caffeine (n = 3);
- (b)
- Twenty days after HIS: control water (n = 5); HIS water (n = 4); control caffeine (n = 3); and HIS caffeine (n = 5).
2.7. Glutathione Reductase Activity
- (a)
- Five days after HIS: control water (n = 5); HIS water (n = 5); control caffeine (n = 5); and HIS caffeine (n = 4);
- (b)
- Twenty days after HIS: control water (n = 5); and HIS water (n = 5).
2.8. Glutathione S-Transferase Activity
- (a)
- Five days after HIS: control water (n = 5); HIS water (n = 5); control caffeine (n = 5); and HIS caffeine (n = 4);
- (b)
- Twenty days after HIS: control water (n = 4); and HIS water (n = 5).
2.9. Na+/K+-ATPase and Mg2+-ATPase Activities Assay
- (a)
- Five days after HIS: control water (n = 5); HIS water (n = 5); control caffeine (n = 4); and HIS caffeine (n = 4);
- (b)
- Twenty days after HIS: control water (n = 7); HIS water (n = 7); control caffeine (n = 5); and HIS caffeine (n = 4).
- (a)
- Five days after HIS: control water (n = 4); HIS water (n = 4); control caffeine (n = 5); and HIS caffeine (n = 4);
- (b)
- Twenty days after HIS: control water (n = 7); HIS water (n = 7); control caffeine (n = 5); and HIS caffeine (n = 3).
2.10. Statistical and Data Analysis
3. Results
3.1. Effect of Chronic Maternal Caffeine Intake on Neonatal Development
3.2. Effect of HIS and Maternal Caffeine Intake during Gestation and Lactation on TBARS Level in Cortex Brain
3.3. Effect of HIS and Maternal Caffeine Intake during Gestation and Lactation on Glutathione Reductase Activity in Cortex Brain
3.4. Effect of HIS and Maternal Caffeine Intake during Gestation and Lactation on Na+/K+-ATPase Activity in Cortex Brain
3.5. Effect of HIS and Maternal Caffeine Intake during Gestation and Lactation on Mg2+-ATPase Activity in Cortex Brain
4. Discussion
4.1. HIS Induce Oxidative Stress in the Medium Term in Cortex Brain
4.2. Maternal Caffeine Intake Promotes Beneficial Effects in Neonates Submitted to HIS
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Frary, C.D.; Johnson, R.K.; Wang, M.Q. Food Sources and Intakes of Caffeine in the Diets of Persons in the United States. J. Am. Diet. Assoc. 2005, 105, 110–113. [Google Scholar] [CrossRef]
- Brazier, J.L.; Ritter, J.; Berland, M.; Khenfer, D.; Faucon, G. Pharmacokinetics of Caffeine during and after Pregnancy. Dev. Pharmacol. Ther. 1983, 6, 315–322. [Google Scholar] [CrossRef]
- Fredholm, B.B.; Bättig, K.; Holmén, J.; Nehlig, A.; Zvartau, E.E. Actions of Caffeine in the Brain with Special Reference to Factors That Contribute to Its Widespread Use. Pharmacol. Rev. 1999, 51, 83–133. [Google Scholar]
- Iglesias, I.; León, D.; Ruiz, M.A.; Albasanz, J.L.; Martín, M. Chronic Intake of Caffeine during Gestation down Regulates Metabotropic Glutamate Receptors in Maternal and Fetal Rat Heart. Amino Acids 2006, 30, 257–266. [Google Scholar] [CrossRef]
- Lorenzo, A.M.; León, D.; Castillo, C.A.; Ruiz, M.A.; Albasanz, J.L.; Martín, M. Maternal Caffeine Intake during Gestation and Lactation Down-Regulates Adenosine A1 Receptor in Rat Brain from Mothers and Neonates. J. Neurosci. Res. 2010, 88, 1252–1261. [Google Scholar] [CrossRef]
- Ősz, B.-E.; Jîtcă, G.; Ștefănescu, R.-E.; Pușcaș, A.; Tero-Vescan, A.; Vari, C.-E. Caffeine and Its Antioxidant Properties-It Is All about Dose and Source. Int. J. Mol. Sci. 2022, 23, 13074. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, V.; Mishra, A.; Singh, S.; Shukla, S. Caffeine Improves Memory and Cognition via Modulating Neural Progenitor Cell Survival and Decreasing Oxidative Stress in Alzheimer’s Rat Model. Curr. Alzheimer Res. 2023, 20, 175–189. [Google Scholar] [CrossRef] [PubMed]
- Souza, M.A.; Mota, B.C.; Gerbatin, R.R.; Rodrigues, F.S.; Castro, M.; Fighera, M.R.; Royes, L.F.F. Antioxidant Activity Elicited by Low Dose of Caffeine Attenuates Pentylenetetrazol-Induced Seizures and Oxidative Damage in Rats. Neurochem. Int. 2013, 62, 821–830. [Google Scholar] [CrossRef]
- Endesfelder, S.; Weichelt, U.; Strauß, E.; Schlör, A.; Sifringer, M.; Scheuer, T.; Bührer, C.; Schmitz, T. Neuroprotection by Caffeine in Hyperoxia-Induced Neonatal Brain Injury. Int. J. Mol. Sci. 2017, 18, 187. [Google Scholar] [CrossRef] [PubMed]
- Shinnar, S.; Glauser, T.A. Febrile Seizures. J. Child Neurol. 2002, 17 (Suppl. 1), S44–S52. [Google Scholar] [CrossRef]
- Stafstrom, C.E. Assessing the Behavioral and Cognitive Effects of Seizures on the Developing Brain. Prog. Brain Res. 2002, 135, 377–390. [Google Scholar] [CrossRef]
- McClelland, S.; Dubé, C.M.; Yang, J.; Baram, T.Z. Epileptogenesis after Prolonged Febrile Seizures: Mechanisms, Biomarkers and Therapeutic Opportunities. Neurosci. Lett. 2011, 497, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Dubé, C.M.; Brewster, A.L.; Baram, T.Z. Febrile Seizures: Mechanisms and Relationship to Epilepsy. Brain Dev. 2009, 31, 366–371. [Google Scholar] [CrossRef] [PubMed]
- Baram, T.Z.; Gerth, A.; Schultz, L. Febrile Seizures: An Appropriate-Aged Model Suitable for Long-Term Studies. Brain Res. Dev. Brain Res. 1997, 98, 265–270. [Google Scholar] [CrossRef]
- Crespo, M.; León-Navarro, D.A.; Martín, M. Cerebellar Oxidative Stress and Fine Motor Impairment in Adolescent Rats Exposed to Hyperthermia-Induced Seizures Is Prevented by Maternal Caffeine Intake during Gestation and Lactation. Eur. J. Pharmacol. 2018, 822, 186–198. [Google Scholar] [CrossRef]
- Crespo, M.; León-Navarro, D.A.; Martín, M. Na+/K+- and Mg2+-ATPases and Their Interaction with AMPA, NMDA and D2 Dopamine Receptors in an Animal Model of Febrile Seizures. Int. J. Mol. Sci. 2022, 23, 14638. [Google Scholar] [CrossRef]
- Novaes, L.S.; Dos Santos, N.B.; Dragunas, G.; Perfetto, J.G.; Leza, J.C.; Scavone, C.; Munhoz, C.D. Repeated Restraint Stress Decreases Na,K-ATPase Activity via Oxidative and Nitrosative Damage in the Frontal Cortex of Rats. Neuroscience 2018, 393, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Aguilar Diaz De Leon, J.; Borges, C.R. Evaluation of Oxidative Stress in Biological Samples Using the Thiobarbituric Acid Reactive Substances Assay. J. Vis. Exp. 2020, 159, e61122. [Google Scholar] [CrossRef]
- León Navarro, D.A.; Crespo, M.; Martín, M. Oxidative Stress in Epileptogenesis: Febrile Seizures, Chemoconvulsant Pilocarpine, and Electrical Stimulation. In Oxidative Stress and Dietary Antioxidants in Neurological Diseases; Elsevier: Amsterdam, The Netherlands, 2020; pp. 81–94. [Google Scholar]
- Rodríguez-Rodríguez, A.; Egea-Guerrero, J.J.; Murillo-Cabezas, F.; Carrillo-Vico, A. Oxidative Stress in Traumatic Brain Injury. Curr. Med. Chem. 2014, 21, 1201–1211. [Google Scholar] [CrossRef]
- Łukawski, K.; Czuczwar, S.J. Oxidative Stress and Neurodegeneration in Animal Models of Seizures and Epilepsy. Antioxidants 2023, 12, 1049. [Google Scholar] [CrossRef]
- Zaniani, N.R.; Roohbakhsh, A.; Moghimi, A.; Mehri, S. Protective Effect of Toll-like Receptor 4 Antagonist on Inflammation, EEG, and Memory Changes Following Febrile Seizure in Wistar Rats. Behav. Brain Res. 2022, 420, 113723. [Google Scholar] [CrossRef] [PubMed]
- Güneş, S.; Dirik, E.; Yiş, U.; Seçkin, E.; Kuralay, F.; Köse, S.; Unalp, A. Oxidant Status in Children after Febrile Seizures. Pediatr. Neurol. 2009, 40, 47–49. [Google Scholar] [CrossRef] [PubMed]
- El-Masry, H.M.A.; Sadek, A.A.; Hassan, M.H.; Ameen, H.H.; Ahmed, H.A. Metabolic Profile of Oxidative Stress and Trace Elements in Febrile Seizures among Children. Metab. Brain Dis. 2018, 33, 1509–1515. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.F.A.; Hoffmann, M.S.; Rambo, L.M.; Ribeiro, L.R.; Lima, F.D.; Furian, A.F.; Oliveira, M.S.; Fighera, M.R.; Royes, L.F.F. The Involvement of Na+, K+-ATPase Activity and Free Radical Generation in the Susceptibility to Pentylenetetrazol-Induced Seizures after Experimental Traumatic Brain Injury. J. Neurol. Sci. 2011, 308, 35–40. [Google Scholar] [CrossRef]
- Bogdanova, A.; Petrushanko, I.Y.; Hernansanz-Agustín, P.; Martínez-Ruiz, A. “Oxygen Sensing” by Na,K-ATPase: These Miraculous Thiols. Front. Physiol. 2016, 7, 314. [Google Scholar] [CrossRef] [PubMed]
- Gibb, R.; Kolb, B. Neonatal Handling Alters Brain Organization but Does Not Influence Recovery from Perinatal Cortical Injury. Behav. Neurosci. 2005, 119, 1375–1383. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Crespo, M.; León-Navarro, D.A.; Martín, M. Maternal Caffeine Consumption during Gestation and Lactation Abolishes Cortical Oxidative Stress and Restores Na+/K+-ATPase Activity in Neonates Exposed to Hyperthermia-Induced Seizures. Biomedicines 2023, 11, 3292. https://doi.org/10.3390/biomedicines11123292
Crespo M, León-Navarro DA, Martín M. Maternal Caffeine Consumption during Gestation and Lactation Abolishes Cortical Oxidative Stress and Restores Na+/K+-ATPase Activity in Neonates Exposed to Hyperthermia-Induced Seizures. Biomedicines. 2023; 11(12):3292. https://doi.org/10.3390/biomedicines11123292
Chicago/Turabian StyleCrespo, María, David Agustín León-Navarro, and Mairena Martín. 2023. "Maternal Caffeine Consumption during Gestation and Lactation Abolishes Cortical Oxidative Stress and Restores Na+/K+-ATPase Activity in Neonates Exposed to Hyperthermia-Induced Seizures" Biomedicines 11, no. 12: 3292. https://doi.org/10.3390/biomedicines11123292
APA StyleCrespo, M., León-Navarro, D. A., & Martín, M. (2023). Maternal Caffeine Consumption during Gestation and Lactation Abolishes Cortical Oxidative Stress and Restores Na+/K+-ATPase Activity in Neonates Exposed to Hyperthermia-Induced Seizures. Biomedicines, 11(12), 3292. https://doi.org/10.3390/biomedicines11123292