
Peak Bone Mass Formation: Modern View of the Problem
 , ,
, ,
Abstract
:1. Introduction
2. Structural Properties of Bone
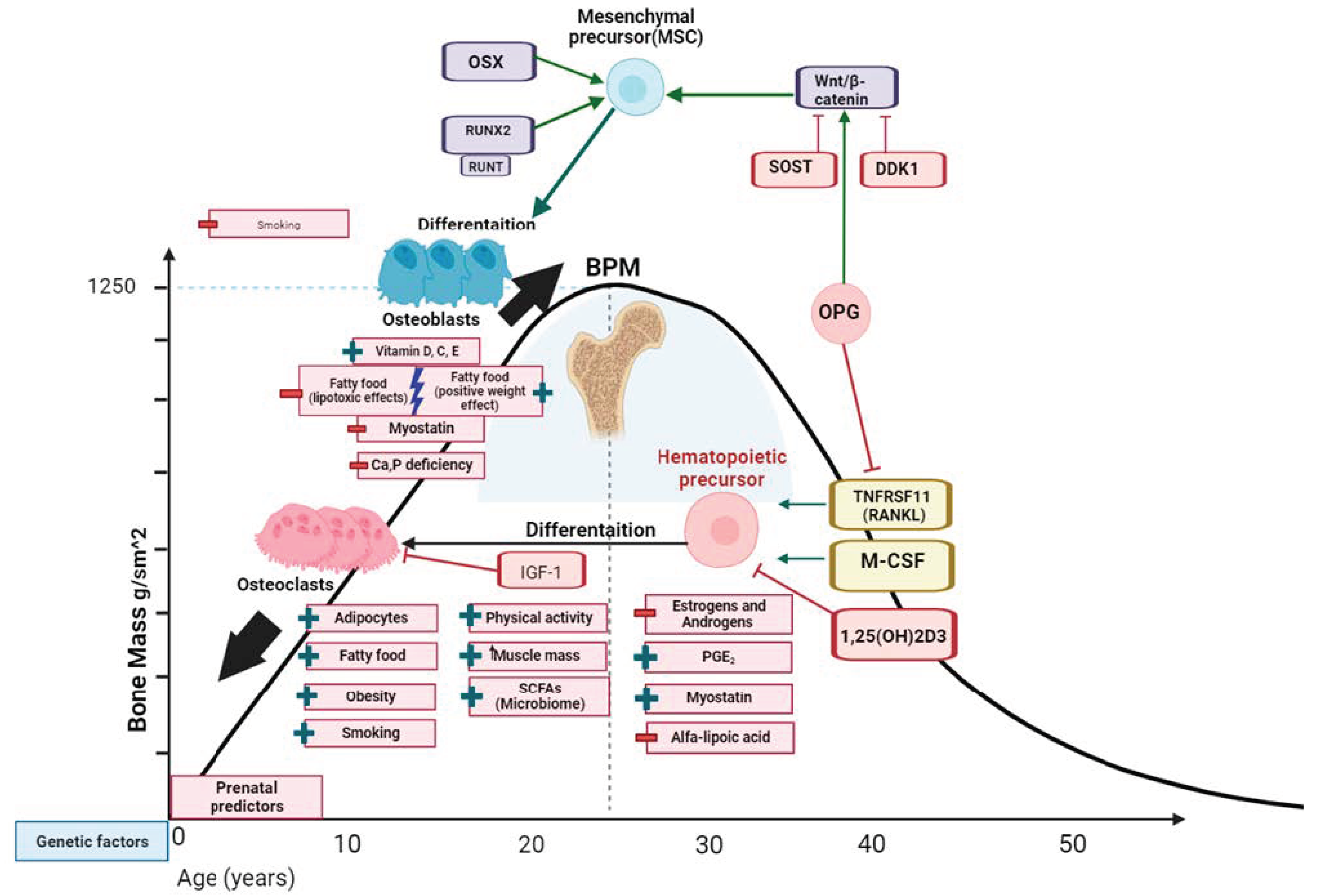
3. Peak Bone Mass and Its Absolute Values
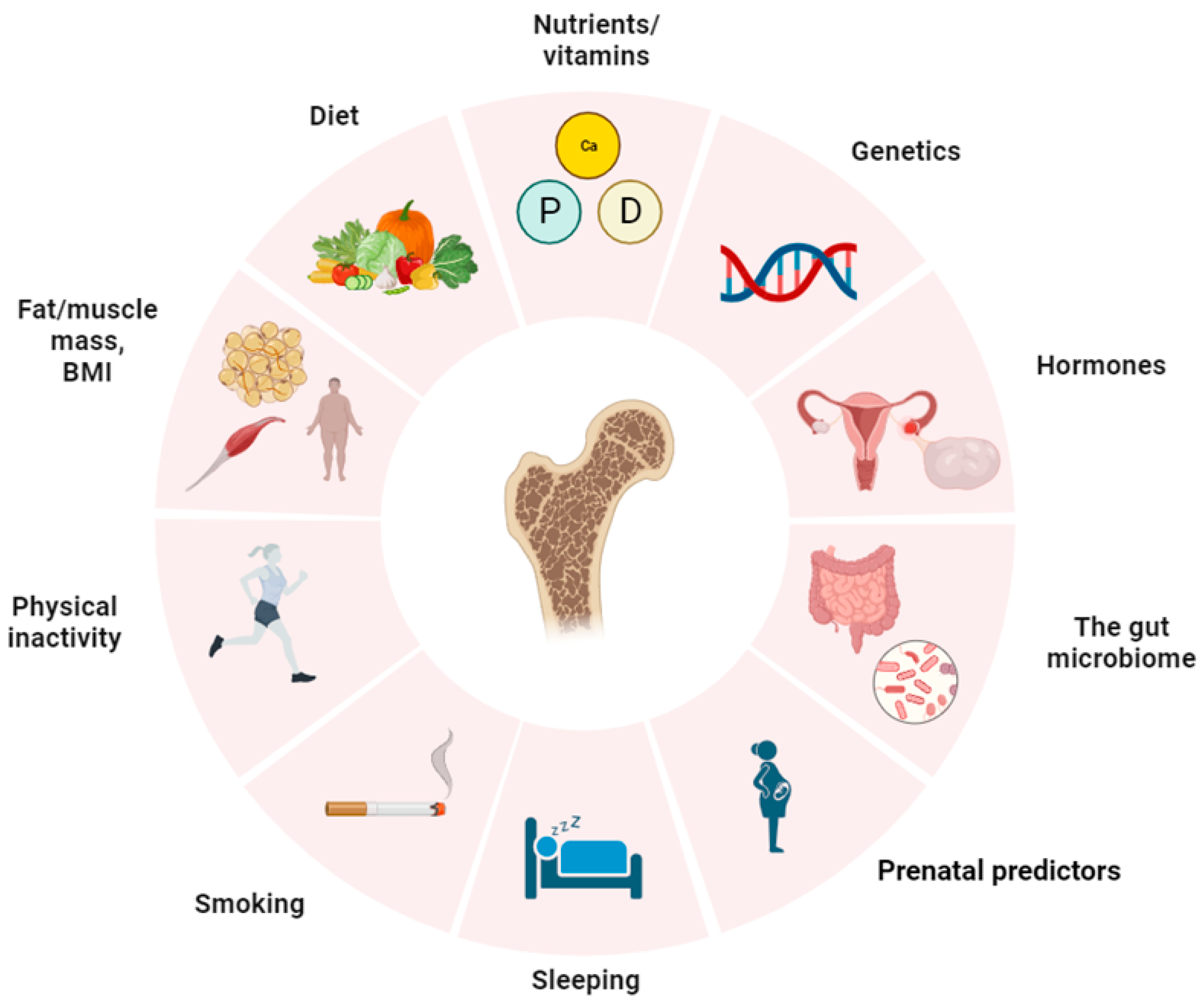
4. Factors Affecting PBM
4.1. Hormonal Background
4.2. Microbiome
4.3. Quality Body Composition
4.4. Smoking
4.5. Nutritional Deficiencies (Calcium, Vitamin D, Phosphorus), Lipids, and Food Character
4.6. Physical Activity, Lifestyle, Psychoemotional Status
4.7. Early Antropometric Characteristics
4.8. Heredity and Genetics
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| # | Authors | Sex | Mean Age (N) | aBMD | System | Country | |
|---|---|---|---|---|---|---|---|
| 1. | Paskova I.G. et al. (2015) [18] | m | 20 (3) | 1.16 ± 0.09 (LS) | Lunar | Russian Federation | |
| 21–25 (16) | 1.20 ± 0.15 (LS) | ||||||
| 26–30 (10) | 1.16 ± 0.1 (LS) | ||||||
| 31–35 (7) | 1.19 ± 0.13(LS) | ||||||
| 36–40 (10) | 1.17 ± 0.18(LS) | ||||||
| 2. | Zakharov I.S. et al. (2014) [19] | f | 16–19 (178) | 1.158 (LS) | Lunar-DPX-NT | Russian Federation | |
| 20–29 (178) | 1.219 (LS) | ||||||
| 30–39 (244) | 1.205 (LS) | ||||||
| 30–39 (244) | 1.068 (LS) | ||||||
| 3. | Looker A. C. et al. (1997) [19] | f | 20–29 (382) | 0.86 ± 0.12 (FN) | Hologic QDR | United States (NHANES III) | |
| m | 20–29 (409) | 0.93± 0.137 (FN) | |||||
| 4. | Henry M.Y. et al. (2004) [20] | f | 11–19 (69) | 0.970 (FN) | Hologic Acclaim 4500A | Great Britain | |
| m | 11–19 (63) | 1.05 (FN) | |||||
| 5. | Karlsson M.K. et al. (2001) [21] | m | 24.4 ± 0.6 (24) | 1.27 ± 0.03 (LS) 1.18 ± 0.03 (FN) | Lunar DPX-L | Sweden | |
| 6. | Bakker I. et al. (2003) [24] | m | 27 (83) | 1.170 ± 0.159 (LS) | Hologic QDR-2000 | Netherlands | |
| f | 27 (97) | 1.143 ± 0.138 (LS) | |||||
| m | 32 (195) | 1.147 ± 0.172 (LS) | |||||
| f | 32 (207) | 1.116 ± 0.129 (LS) | |||||
| m | 36 (170) | 1.110 ± 0.161 (LS) | |||||
| f | 36 (181) | 1.094 ± 0.120 (LS) | |||||
| 7. | Liberato S.C. (2015) [25] | m | 21.8 ± 2.18 (35) | 1.16 ± 0.15 (LS) | Lunar | Australia | |
| 8. | Tveit M. et al. (2014) [23] | m | 23.96 ± 3.85 (135) | 1.36 (0.11) (LS) | 1.27 (0.14) (FN) | Lunar DPX-L | Sweden |
| 35.16 ± 3.17 (113) | 1.27 (0.14) (LS) | 1.02 (0.15) (FN) | |||||
| 43.19 ± 6.61 (149) | 1.20 (0.16) (LS) | 0.98 (0.13) (FN) | |||||
| 9. | Lorentzon M. et al. (2005) [22] | m | 18.9 ± 0.6 (1068) | 1.24 ± 0.15 (LS) 1.17 ± 0.16 (FN) | Lunar Prodigy | Sweden | |
| 10. | Jones G. et al. (2000) [26] | f | 33.5 (278) | 1.07 (0.12) (LS) 0.84 (0.11) (FN) | Hologic QDR2000 | Australia | |
| 11. | Leppa¨la J. (1999) et al. [27] | f | 22 ± 5 (13) | 0.900 ± 0.132 (FN) | Norland XR-26 | Finland | |
| m | 26 ± 4 (28) | 0.952 ± 0.127 (FN) | |||||
| 12. | Ho S.C. et al. (1993) [28] | f | 21–24 (46) | 0.93 ± 0.10 (LS) | 0.79 ± 0.12 (FN) | Norland XR-26 | China |
| 25–28 (50) | 0.96 ± 0.11 (LS) | 0.82 ± 0.12 (FN) | |||||
| 29–32 (57) | 1.03 ± 0.12 (LS) | 0.86 ± 0.11 (FN) | |||||
| 33–36 (80) | 1.02 ± 0.11 (LS) | 0.83 ± 0.10 (FN) | |||||
| 37–40 (60) | 0.99 ± 0.14 (LS) | 0.82 ± 0.14 (FN) | |||||
| 13. | Orito S. et al. (2009) [30] | f | 12–30 (1322) | 0.961 ± 0.119 (LS) | Hologic QDR4500 | Japan | |
| 14. | Yanping D. et al. (2018) [29] | m | 21–30 (113) | 0.986 ± 0.150 (LS) | Hologic Delphi A | China | |
| m | 31–40 (141) | 0.967 ± 0.113 (LS) | |||||
| f | 21–30 (128) | 0.970 ± 0.100 (LS) | |||||
| f | 31–40 (101) | 0.995 ± 0.104 (LS) | |||||
References
- Chew, C.K.; Clarke, B.L. Causes of Low Peak Bone Mass in Women. Maturitas 2018, 111, 61–68. [Google Scholar] [CrossRef]
- Xue, S.; Kemal, O.; Lu, M.; Lix, L.M.; Leslie, W.D.; Yang, S. Age at Attainment of Peak Bone Mineral Density and Its Associated Factors: The National Health and Nutrition Examination Survey 2005–2014. Bone 2020, 131, 115163. [Google Scholar] [CrossRef]
- Loftus, A.; Cappariello, A.; George, C.; Ucci, A.; Shefferd, K.; Green, A.; Paone, R.; Ponzetti, M.; Delle Monache, S.; Muraca, M.; et al. Extracellular Vesicles from Osteotropic Breast Cancer Cells Affect Bone Resident Cells. J. Bone Miner. Res. 2020, 35, 396–412. [Google Scholar] [CrossRef]
- Farr, J.N.; Khosla, S. Skeletal Changes through the Lifespan—From Growth to Senescence. Nat. Rev. Endocrinol. 2015, 11, 513–521. [Google Scholar] [CrossRef] [PubMed]
- Hadji, P.; Colli, E.; Regidor, P.A. Bone Health in Estrogen-Free Contraception. Osteoporos. Int. 2019, 30, 2391–2400. [Google Scholar] [CrossRef]
- Golden, N.H. Bones and Birth Control in Adolescent Girls. J. Pediatr. Adolesc. Gynecol. 2020, 33, 249–254. [Google Scholar] [CrossRef]
- Hernandez, C.J.; Carter, D.R.; Beaupré, G.S. A Theoretical Analysis of the Relative Influences of Peak BMD, Age-Related Bone Loss and Menopause on the Development of Osteoporosis. Osteoporos. Int. 2003, 14, 843–847. [Google Scholar] [CrossRef] [PubMed]
- Ensrud, K.E.; Crandall, C.J. Osteoporosis. Ann. Intern Med. 2017, 167, ITC17–ITC32. [Google Scholar] [CrossRef] [PubMed]
- Burt, L.A.; Hanley, D.A.; Boyd, S.K. Cross-Sectional Versus Longitudinal Change in a Prospective HR-PQCT Study. J. Bone Miner. Res. 2017, 32, 1505–1513. [Google Scholar] [CrossRef] [PubMed]
- Gilsanz, V.; Wren, T.A.L.; Ponrartana, S.; Mora, S.; Rosen, C.J. Sexual Dimorphism and the Origins of Human Spinal Health. Endocr. Rev. 2018, 39, 221–239. [Google Scholar] [CrossRef]
- Chevalley, T.; Rizzoli, R. Acquisition of Peak Bone Mass. Best Pract. Res. Clin. Endocrinol. Metab. 2022, 36, 101616. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, Y.; Kang, H.; Liu, W.; Liu, P.; Zhang, J.; Harris, S.E.; Wu, D. Sclerostin Binds to LRP5/6 and Antagonizes Canonical Wnt Signaling. J. Biol. Chem. 2005, 280, 19883–19887. [Google Scholar] [CrossRef]
- Merrick, D.; Mistry, K.; Wu, J.; Gresko, N.; Baggs, J.E.; Hogenesch, J.B.; Sun, Z.; Caplan, M.J. Polycystin-1 Regulates Bone Development through an Interaction with the Transcriptional Coactivator TAZ. Hum. Mol. Genet. 2019, 28, 16–30. [Google Scholar] [CrossRef] [PubMed]
- Santos, L.; Elliott-Sale, K.J.; Sale, C. Exercise and Bone Health across the Lifespan. Biogerontology 2017, 18, 931–946. [Google Scholar] [CrossRef] [PubMed]
- Lindgren, E.; Rosengren, B.E.; Karlsson, M.K. Does Peak Bone Mass Correlate with Peak Bone Strength? Cross-Sectional Normative Dual Energy X-ray Absorptiometry Data in 1052 Men Aged 18–28 Years. BMC Musculoskelet. Disord. 2019, 20, 404. [Google Scholar] [CrossRef]
- Kröger, H.; Vainio, P.; Nieminen, J.; Kotaniemi, A. Comparison of Different Models for Interpreting Bone Mineral Density Measurements Using DXA and MRI Technology. Bone 1995, 17, 157–159. [Google Scholar] [CrossRef]
- Zakharov, I.S.; Kolpinsky, G.I.; Ushakova, G.A. Referential indices of bone mineral density of lumbar vertebrae for women of kuzbass. Politravma 2014, 3, 3–14. [Google Scholar]
- Pashkova, I.G.; Gaivoronsky, I.V.; Gaivoronskaya, M.G. Age features of bone mineral density lumbar spine in adult males. Bull. Russ. Mil. Med. Acad. 2015, 1, 111–114. [Google Scholar]
- Looker, A.C.; Orwoll, E.S.; Johnston, C.C.; Lindsay, R.L.; Wahner, H.W.; Dunn, W.L.; Calvo, M.S.; Harris, T.B.; Heyse, S.P. Prevalence of Low Femoral Bone Density in Older U.S. Adults from NHANES III. J. Bone Miner. Res. 1997, 12, 1761–1768. [Google Scholar] [CrossRef]
- Henry, Y.M.; Fatayerji, D.; Eastell, R. Attainment of Peak Bone Mass at the Lumbar Spine, Femoral Neck and Radius in Men and Women: Relative Contributions of Bone Size and Volumetric Bone Mineral Density. Osteoporos. Int. 2004, 15, 263–273. [Google Scholar] [CrossRef]
- Karlsson, M.; Magnusson, H.; Karlsson, C.; Seeman, E. The Duration of Exercise as a Regulator of Bone Mass. Bone 2001, 28, 128–132. [Google Scholar] [CrossRef]
- Lorentzon, M.; Mellström, D.; Ohlsson, C. Age of Attainment of Peak Bone Mass Is Site Specific in Swedish Men—The GOOD Study. J. Bone Miner. Res. 2005, 20, 1223–1227. [Google Scholar] [CrossRef] [PubMed]
- Tveit, M.; Rosengren, B.E.; Nilsson, J.A.; Karlsson, M.K. Exercise in Youth: High Bone Mass, Large Bone Size, and Low Fracture Risk in Old Age. Scand. J. Med. Sci. Sports 2015, 25, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Bakker, I.; Twisk, J.W.R.; Van Mechelen, W.; Roos, J.C.; Kemper, H.C.G. Ten-Year Longitudinal Relationship between Physical Activity and Lumbar Bone Mass in (Young) Adults. J. Bone Miner. Res. 2003, 18, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Liberato, S.C.; Maple-Brown, L.; Bressan, J. Association between Bone Mineralization, Body Composition, and Cardiorespiratory Fitness Level in Young Australian Men. J. Clin. Densitom. 2015, 18, 187–191. [Google Scholar] [CrossRef]
- Jones, G.; Nguyen, T.V. Associations between Maternal Peak Bone Mass and Bone Mass in Prepubertal Male and Female Children. J. Bone Miner. Res. 2000, 15, 1998–2004. [Google Scholar] [CrossRef]
- Leppälä, J.; Kannus, P.; Niemi, S.; Sievänen, H.; Vuori, I.; Järvinen, M. An Early-Life Femoral Shaft Fracture and Bone Mineral Density at Adulthood. Osteoporos. Int. 1999, 10, 337–342. [Google Scholar] [CrossRef]
- Ho, S.C.; Hsu, S.Y.; Leung, P.C.; Chan, C.; Swaminathan, R.; Fan, Y.K.; Chan, S.S. A Longitudinal Study of the Determinants of Bone Mass in Chinese Women Aged 21 to 40 I. Baseline Association of Anthropometric Measurements with Bone Mineral Density. Ann. Epidemiol. 1993, 3, 256–263. [Google Scholar] [CrossRef]
- Du, Y.; Zhu, H.; Zheng, S.; Zhu, X.; Zhang, X.; Xue, S.; Li, H.; Hong, W.; Tang, W.; Chen, M.; et al. Age and Sex Effects on the Relationship between Body Composition and Hip Geometric Structure in Males and Females from East China. Arch. Osteoporos. 2018, 13, 79. [Google Scholar] [CrossRef]
- Orito, S.; Kuroda, T.; Onoe, Y.; Sato, Y.; Ohta, H. Age-Related Distribution of Bone and Skeletal Parameters in 1322 Japanese Young Women. J. Bone Miner. Metab. 2009, 27, 698–704. [Google Scholar] [CrossRef]
- Zhu, X.; Zheng, H. Factors Influencing Peak Bone Mass Gain. Front. Med. 2021, 15, 53–69. [Google Scholar] [CrossRef] [PubMed]
- Almeida, M.; Laurent, M.R.; Dubois, V.; Claessens, F.; O’Brien, C.A.; Bouillon, R.; Vanderschueren, D.; Manolagas, S.C. Estrogens and Androgens in Skeletal Physiology and Pathophysiology. Physiol. Rev. 2017, 97, 135–187. [Google Scholar] [CrossRef] [PubMed]
- Carson, J.A.; Manolagas, S.C. Effects of Sex Steroids on Bones and Muscles: Similarities, Parallels, and Putative Interactions in Health and Disease. Bone 2015, 80, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Manolagas, S.C. From Estrogen-Centric to Aging and Oxidative Stress: A Revised Perspective of the Pathogenesis of Osteo-porosis. Endocr. Rev. 2010, 31, 266–300. [Google Scholar] [CrossRef]
- Park-Min, K.-H. Mechanisms Involved in Normal and Pathological Osteoclastogenesis. Cell. Mol. Life Sci. 2018, 75, 2519–2528. [Google Scholar] [CrossRef]
- Yasuda, H. Discovery of the RANKL/RANK/OPG System. J. Bone Miner. Metab. 2021, 39, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Szeliga, A.; Maciejewska-Jeske, M.; Męczekalski, B. Bone Health and Evaluation of Bone Mineral Density in Patients with Premature Ovarian Insufficiency. Menopausal Rev. 2018, 17, 112–116. [Google Scholar] [CrossRef]
- Vlot, M.C.; Wiepjes, C.M.; de Jongh, R.T.; T’Sjoen, G.; Heijboer, A.C.; den Heijer, M. Gender-Affirming Hormone Treatment Decreases Bone Turnover in Transwomen and Older Transmen. J. Bone Miner. Res. 2019, 34, 1862–1872. [Google Scholar] [CrossRef]
- Yan, J.; Charles, J.F. Gut Microbiome and Bone: To Build, Destroy, or Both? Curr. Osteoporos. Rep. 2017, 15, 376–384. [Google Scholar] [CrossRef]
- Lucas, S.; Omata, Y.; Hofmann, J.; Böttcher, M.; Iljazovic, A.; Sarter, K.; Albrecht, O.; Schulz, O.; Krishnacoumar, B.; Krönke, G.; et al. Short-Chain Fatty Acids Regulate Systemic Bone Mass and Protect from Pathological Bone Loss. Nat. Commun. 2018, 9, 55. [Google Scholar] [CrossRef]
- Li, J.-Y.; Yu, M.; Pal, S.; Tyagi, A.M.; Dar, H.; Adams, J.; Neale Weitzmann, M.; Jones, R.M.; Pacifici, R. Parathyroid Hormone–Dependent Bone Formation Requires Butyrate Production by Intestinal Microbiota. J. Clin. Investig. 2020, 130, 1767–1781. [Google Scholar] [CrossRef]
- Jansson, P.-A.; Curiac, D.; Lazou Ahrén, I.; Hansson, F.; Martinsson Niskanen, T.; Sjögren, K.; Ohlsson, C. Probiotic Treatment Using a Mix of Three Lactobacillus Strains for Lumbar Spine Bone Loss in Postmenopausal Women: A Randomised, Double-Blind, Placebo-Controlled, Multicentre Trial. Lancet Rheumatol. 2019, 1, e154–e162. [Google Scholar] [CrossRef]
- Whisner, C.M.; Castillo, L.F. Prebiotics, Bone and Mineral Metabolism. Calcif. Tissue Int. 2018, 102, 443–479. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Xie, Z.; Sun, J.; Huang, S.; Chen, Y.; Li, C.; Sun, X.; Xia, B.; Tian, L.; Guo, C.; et al. Gut Microbiome Reveals Specific Dysbiosis in Primary Osteoporosis. Front. Cell. Infect. Microbiol. 2020, 10, 160. [Google Scholar] [CrossRef]
- Behera, J.; Ison, J.; Tyagi, S.C.; Tyagi, N. The Role of Gut Microbiota in Bone Homeostasis. Bone 2020, 135, 115317. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, A.G.; Sundh, D.; Bäckhed, F.; Lorentzon, M. Lactobacillus reuteri Reduces Bone Loss in Older Women with Low Bone Mineral Density: A Randomized, Placebo-Controlled, Double-Blind, Clinical Trial. J. Intern. Med. 2018, 284, 307–317. [Google Scholar] [CrossRef]
- Jafarnejad, S.; Djafarian, K.; Fazeli, M.R.; Yekaninejad, M.S.; Rostamian, A.; Keshavarz, S.A. Effects of a Multispecies Probiotic Supplement on Bone Health in Osteopenic Postmenopausal Women: A Randomized, Double-Blind, Controlled Trial. J. Am. Coll. Nutr. 2017, 36, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wu, Y.; Wang, Y.; Xu, H.; Mei, X.; Yu, D.; Wang, Y.; Li, W. Antioxidant Properties of Probiotic Bacteria. Nutrients 2017, 9, 521. [Google Scholar] [CrossRef] [PubMed]
- Feigenson, M.; Eliseev, R.A.; Jonason, J.H.; Mills, B.N.; O’Keefe, R.J. PGE2 Receptor Subtype 1 (EP1) Regulates Mesenchymal Stromal Cell Osteogenic Differentiation by Modulating Cellular Energy Metabolism. J. Cell. Biochem. 2017, 118, 4383–4393. [Google Scholar] [CrossRef]
- Pino, A.M.; Rodríguez, J.P. Is Fatty Acid Composition of Human Bone Marrow Significant to Bone Health? Bone 2019, 118, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Jing, Z.; Hu, W.; Yu, J.; Cui, X. α-Linolenic Acid Inhibits Receptor Activator of NF-κB Ligand Induced (RANKL-Induced) Osteoclastogenesis and Prevents Inflammatory Bone Loss via Downregulation of Nuclear Factor-KappaB-Inducible Nitric Oxide Synthases (NF-κB-iNOS) Signaling Pathways. Med. Sci. Monit. 2017, 23, 5056–5069. [Google Scholar] [CrossRef] [PubMed]
- Fintini, D.; Cianfarani, S.; Cofini, M.; Andreoletti, A.; Ubertini, G.M.; Cappa, M.; Manco, M. The Bones of Children with Obesity. Front. Endocrinol. 2020, 11, 200. [Google Scholar] [CrossRef] [PubMed]
- Loke, S.-S.; Chang, H.-W.; Li, W.-C. Association between Metabolic Syndrome and Bone Mineral Density in a Taiwanese Elderly Population. J. Bone Miner. Metab. 2018, 36, 200–208. [Google Scholar] [CrossRef]
- Palermo, A.; Tuccinardi, D.; De Feudis, G.; Watanabe, M.; D’Onofrio, L.; Lauria Pantano, A.; Napoli, N.; Pozzilli, P.; Manfrini, S. BMI and BMD: The Potential Interplay between Obesity and Bone Fragility. Int. J. Environ. Res. Public Health 2016, 13, 544. [Google Scholar] [CrossRef] [PubMed]
- Hetherington-Rauth, M.; Bea, J.W.; Blew, R.M.; Funk, J.L.; Hingle, M.D.; Lee, V.R.; Roe, D.J.; Wheeler, M.D.; Lohman, T.G.; Going, S.B. Relative Contributions of Lean and Fat Mass to Bone Strength in Young Hispanic and Non-Hispanic Girls. Bone 2018, 113, 144–150. [Google Scholar] [CrossRef]
- Xiao, Z.; Xu, H. Gender-Specific Body Composition Relationships between Adipose Tissue Distribution and Peak Bone Mineral Density in Young Chinese Adults. BioMed Res. Int. 2020, 2020, 6724749. [Google Scholar] [CrossRef]
- Nilsen, O.A.; Ahmed, L.A.; Winther, A.; Christoffersen, T.; Thrane, G.; Evensen, E.; Furberg, A.S.; Grimnes, G.; Dennison, E.; Emaus, N. Body Weight and Body Mass Index Influence Bone Mineral Density in Late Adolescence in a Two-Year Follow-Up Study. The Tromsø Study: Fit Futures. JBMR Plus 2019, 3, e10195. [Google Scholar] [CrossRef]
- Lagerquist, M.K.; Gustafsson, K.L.; Henning, P.; Farman, H.; Wu, J.; Sjögren, K.; Koskela, A.; Tuukkanen, J.; Ohlsson, C.; Wernstedt Asterholm, I.; et al. Acute Fat Loss Does not Affect Bone Mass. Sci. Rep. 2021, 11, 14177. [Google Scholar] [CrossRef]
- Laurent, M.R.; Dubois, V.; Claessens, F.; Verschueren, S.M.P.; Vanderschueren, D.; Gielen, E.; Jardí, F. Muscle-Bone Interactions: From Experimental Models to the Clinic? A Critical Update. Mol. Cell. Endocrinol. 2016, 432, 14–36. [Google Scholar] [CrossRef]
- Nguyen, H.G.; Pham, M.T.D.; Ho-Pham, L.T.; Nguyen, T.V. Lean Mass and Peak Bone Mineral Density. Osteoporos. Sarcopenia 2020, 6, 212–216. [Google Scholar] [CrossRef]
- Winther, A.; Jørgensen, L.; Ahmed, L.A.; Christoffersen, T.; Furberg, A.S.; Grimnes, G.; Jorde, R.; Nilsen, O.A.; Dennison, E.; Emaus, N. Bone Mineral Density at the Hip and its Relation to Fat Mass and Lean Mass in Adolescents: The Tromsø Study, Fit Futures. BMC Musculoskelet. Disord. 2018, 19, 21. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.; Ma, Y.; Wang, X.; Jin, F.; Ge, S. Short-Term Muscle Atrophy Caused by Botulinum Toxin-A Local Injection Impairs Fracture Healing in the Rat Femur. J. Orthop. Res. 2012, 30, 574–580. [Google Scholar] [CrossRef]
- Li, G.; Zhang, L.; Wang, D.; AI Qudsy, L.; Jiang, J.X.; Xu, H.; Shang, P. Muscle-Bone Crosstalk and Potential Therapies for Sarco-Osteoporosis. J. Cell. Biochem. 2019, 120, 14262–14273. [Google Scholar] [CrossRef]
- Bonewald, L. Use it or Lose it to Age: A Review of Bone and Muscle Communication. Bone 2019, 120, 212–218. [Google Scholar] [CrossRef]
- Puolakkainen, T.; Ma, H.; Kainulainen, H.; Pasternack, A.; Rantalainen, T.; Ritvos, O.; Heikinheimo, K.; Hulmi, J.J.; Kiviranta, R. Treatment with Soluble Activin Type IIB-Receptor Improves Bone Mass and Strength in a Mouse Model of Duchenne Muscular Dystrophy. BMC Musculoskelet. Disord. 2017, 18, 20. [Google Scholar] [CrossRef]
- Chen, Y.S.; Guo, Q.; Guo, L.J.; Liu, T.; Wu, X.P.; Lin, Z.Y.; He, H.B.; Jiang, T.J. GDF8 Inhibits Bone Formation and Promotes Bone Resorption in Mice. Clin. Exp. Pharmacol. Physiol. 2017, 44, 500–508. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Peng, Y.; Zhao, W.; Pan, J.; Ksiezak-Reding, H.; Cardozo, C.; Wu, Y.; Pajevic, P.D.; Bonewald, L.F.; Bauman, W.A.; et al. Myostatin Inhibits Osteoblastic Differentiation by Suppressing Osteocyte-Derived Exosomal microRNA-218: A Novel Mechanism in Muscle-Bone Communication. J. Biol. Chem. 2017, 292, 11021–11033. [Google Scholar] [CrossRef]
- Callréus, M.; McGuigan, F.; Åkesson, K. Adverse Effects of Smoking on Peak Bone Mass May Be Attenuated by Higher Body Mass Index in Young Female Smokers. Calcif. Tissue Int. 2013, 93, 517–525. [Google Scholar] [CrossRef]
- Wu, Z.J.; Zhao, P.; Liu, B.; Yuan, Z.C. Effect of Cigarette Smoking on Risk of Hip Fracture in Men: A Meta-Analysis of 14 Prospective Cohort Studies. PLoS ONE 2016, 11, e0168990. [Google Scholar] [CrossRef]
- Yuan, S.; Michaëlsson, K.; Wan, Z.; Larsson, S.C. Associations of Smoking and Alcohol and Coffee Intake with Fracture and Bone Mineral Density: A Mendelian Randomization Study. Calcif. Tissue Int. 2019, 105, 582–588. [Google Scholar] [CrossRef] [PubMed]
- Al-Bashaireh, A.M.; Haddad, L.G.; Weaver, M.; Chengguo, X.; Kelly, D.L.; Yoon, S. The Effect of Tobacco Smoking on Bone Mass: An Overview of Pathophysiologic Mechanisms. J. Osteoporos. 2018, 2018, 1206235. [Google Scholar] [CrossRef] [PubMed]
- Pompe, E.; Bartstra, J.; Verhaar, H.J.; de Koning, H.J.; van der Aalst, C.M.; Oudkerk, M.; Vliegenthart, R.; Lammers, J.-W.; de Jong, P.; Hoesein, F.M. Bone Density Loss on Computed Tomography at 3-Year Follow-up in Current Compared to Former Male Smokers. Eur. J. Radiol. 2017, 89, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Al-Bashaireh, A.M.; Haddad, L.G.; Weaver, M.; Kelly, D.L.; Chengguo, X.; Yoon, S. The Effect of Tobacco Smoking on Mus-Culoskeletal Health: A Systematic Review. J. Environ. Public Health 2018, 2018, 4184190. [Google Scholar] [CrossRef] [PubMed]
- Strozyk, D.; Gress, T.M.; Breitling, L.P. Smoking and Bone Mineral Density: Comprehensive Analyses of the Third NATIONAL Health and Nutrition Examination Survey (NHANES III). Arch. Osteoporos. 2018, 13, 16. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.Y.; Lai, J.C.Y.; Huang, W.L.; Hsu, C.L.; Chen, S.J. Effects of Sex, Tobacco Smoking, and Alcohol Consumption Osteoporosis Development: Evidence from Taiwan Biobank Participants. Tob. Induc. Dis. 2021, 19, 8210532. [Google Scholar] [CrossRef]
- Sophocleous, A.; Robertson, R.; Ferreira, N.B.; McKenzie, J.; Fraser, W.D.; Ralston, S.H. Heavy Cannabis Use Is Associated with Low Bone Mineral Density and an Increased Risk of Fractures. Am. J. Med. 2017, 130, 214–221. [Google Scholar] [CrossRef]
- Raphael-Mizrahi, B.; Gabet, Y. The Cannabinoids Effect on Bone Formation and Bone Healing. Curr. Osteoporos. Rep. 2020, 18, 433–438. [Google Scholar] [CrossRef] [PubMed]
- Goltzman, D. Functions of Vitamin D in Bone. Histochem. Cell Biol. 2018, 149, 305–312. [Google Scholar] [CrossRef]
- LeBoff, M.S.; Chou, S.H.; Ratliff, K.A.; Cook, N.R.; Khurana, B.; Kim, E.; Cawthon, P.M.; Bauer, D.C.; Black, D.; Gallagher, J.C.; et al. Supplemental Vitamin D and Incident Fractures in Midlife and Older Adults. N. Engl. J. Med. 2022, 387, 299–309. [Google Scholar] [CrossRef]
- Gou, G.H.; Tseng, F.J.; Wang, S.H.; Chen, P.J.; Shyu, J.F.; Pan, R.Y. Nutritional Factors Associated with Femoral Neck Bone Mineral Density in Children and Adolescents. BMC Musculoskelet. Disord. 2019, 20, 520. [Google Scholar] [CrossRef]
- Zhou, W.; Langsetmo, L.; Berger, C.; Poliquin, S.; Kreiger, N.; Barr, S.I.; Kaiser, S.M.; Josse, R.G.; Prior, J.C.; Towheed, T.E.; et al. Longitudinal Changes in Calcium and Vitamin D Intakes and Relationship to Bone Mineral Density in a Prospective Population-Based Study: The Canadian Multicentre Osteoporosis Study (CaMos). J. Musculoskelet. Neuronal Interact. 2013, 13, 470. [Google Scholar] [PubMed]
- Neville, C.E.; Robson, P.J.; Murray, L.J.; Strain, J.J.; Twisk, J.; Gallagher, A.M.; McGuinness, M.; Cran, G.W.; Ralston, S.H.; Boreham, C.A.G. The Effect of Nutrient Intake on Bone Mineral Status in Young Adults: The Northern Ireland Young Hearts Project. Calcif. Tissue Int. 2002, 70, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Krstic, N.; Bishop, N.; Curtis, B.; Cooper, C.; Harvey, N.; Lilycrop, K.; Murray, R.; Owen, R.; Reilly, G.; Skerry, T.; et al. Early Life Vitamin D Depletion and Mechanical Loading Determine Methylation Changes in the RUNX2, RXRA, and Osterix Promoters in Mice. Genes Nutr. 2022, 17, 7. [Google Scholar] [CrossRef]
- Li, H.; Hou, J.-L.; Yang, W.-Y.; Zhang, Q.; Feng, H.; Wang, X.-B.; Deng, K.-L.; Mao, X.-L.; Xiao, S.-M. Associations between Dietary Antioxidant Vitamin Intake and the Changes in Bone Mass in Chinese Adolescents: A 2.5-Year Longitudinal Study. Nutrients 2022, 14, 4187. [Google Scholar] [CrossRef]
- Movassagh, E.Z.; Baxter-Jones, A.D.G.; Kontulainen, S.; Whiting, S.; Szafron, M.; Vatanparast, H. Vegetarian-Style Dietary Pattern during Adolescence Has Long-Term Positive Impact on Bone from Adolescence to Young Adulthood: A Longitudinal Study. Nutr. J. 2018, 17, 36. [Google Scholar] [CrossRef] [PubMed]
- Scheller, E.L.; Khoury, B.; Moller, K.L.; Wee, N.K.Y.; Khandaker, S.; Kozloff, K.M.; Abrishami, S.H.; Zamarron, B.F.; Singer, K. Changes in Skeletal Integrity and Marrow Adiposity during High-Fat Diet and after Weight Loss. Front. Endocrinol. 2016, 7, 102. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Yu, X. Fat, Sugar, and Bone Health: A Complex Relationship. Nutrients 2017, 9, 506. [Google Scholar] [CrossRef] [PubMed]
- Weaver, C.M.; Gordon, C.M.; Janz, K.F.; Kalkwarf, H.J.; Lappe, J.M.; Lewis, R.; O’Karma, M.; Wallace, T.C.; Zemel, B.S. The National Osteoporosis Foundation’s Position Statement on Peak Bone Mass Development and Lifestyle Factors: A Systematic Review and Implementation Recommendations. Osteoporos. Int. 2016, 27, 1281–1386. [Google Scholar] [CrossRef]
- Karlsson, M.K.; Rosengren, B.E. Exercise and Peak Bone Mass. Curr. Osteoporos. Rep. 2020, 18, 285–290. [Google Scholar] [CrossRef]
- Nguyen, V.H. School-Based Exercise Interventions Effectively Increase Bone Mineralization in Children and Adolescents. Osteoporos. Sarcopenia 2018, 4, 39–46. [Google Scholar] [CrossRef]
- Larsen, M.N.; Nielsen, C.M.; Helge, E.W.; Madsen, M.; Manniche, V.; Hansen, L.; Hansen, P.R.; Bangsbo, J.; Krustrup, P. Positive Effects on Bone Mineralisation and Muscular Fitness after 10 Months of Intense School-Based Physical Training for Children Aged 8–10 Years: The Fit First Randomised Controlled Trial. Br. J. Sport. Med. 2018, 52, 254–260. [Google Scholar] [CrossRef]
- Tang, Y.; Wang, S.; Yi, Q.; Xia, Y.; Geng, B. Sleep Pattern and Bone Mineral Density: A Cross-Sectional Study of National Health and Nutrition Examination Survey (NHANES) 2017–2018. Arch. Osteoporos. 2021, 16, 157. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.; Liu, X.; Jia, G.; Liu, Z.; Zhang, K.; He, L.; Geng, B.; Xia, Y. The Association between Depression and Bone Metabolism: A US Nationally Representative Cross-Sectional Study. Arch. Osteoporos. 2022, 17, 113. [Google Scholar] [CrossRef] [PubMed]
- Mikkola, T.M.; von Bonsdorff, M.B.; Osmond, C.; Salonen, M.K.; Kajantie, E.; Cooper, C.; Välimäki, M.J.; Eriksson, J.G. Childhood Growth Predicts Higher Bone Mass and Greater Bone Area in Early Old Age: Findings among a Subgroup of Women from the Helsinki Birth Cohort Study. Osteoporos. Int. 2017, 28, 2717–2722. [Google Scholar] [CrossRef]
- Baird, J.; Kurshid, M.A.; Kim, M.; Harvey, N.; Dennison, E.; Cooper, C. Does Birthweight Predict Bone Mass in Adulthood? A Systematic Review and Meta-Analysis. Osteoporos. Int. 2011, 22, 1323–1334. [Google Scholar] [CrossRef] [PubMed]
- Callréus, M.; McGuigan, F.; Åkesson, K. Birth Weight is More Important for Peak Bone Mineral Content than for Bone Density: The PEAK-25 Study of 1,061 Young Adult Women. Osteoporos. Int. 2013, 24, 1347–1355. [Google Scholar] [CrossRef]
- Akhiiarova, K.E.; Gantseva, K.K.; Khusainova, R.I.; Tyurin, A.V. Phenotypic Manifestations of Connective Tissue Dysplasia in Individuals with Joint Hypermobility. Med. Counc. 2022, 156–161. [Google Scholar] [CrossRef]
- Yalaev, B.; Tyurin, A.; Prokopenko, I.; Karunas, A.; Khusnutdinova, E.; Khusainova, R. Using a Polygenic Score to Predict the Risk of Developing Primary Osteoporosis. Int. J. Mol. Sci. 2022, 23, 10021. [Google Scholar] [CrossRef]
- Yu, W.J.; Zhang, Z.; Fu, W.Z.; He, J.W.; Wang, C.; Zhang, Z.L. Association between LGR4 Polymorphisms and Peak Bone Mineral Density and Body Composition. J. Bone Miner. Metab. 2020, 38, 658–669. [Google Scholar] [CrossRef]
- Zheng, Y.; Wang, C.; Zhang, H.; Shao, C.; Gao, L.H.; Li, S.S.; Yu, W.J.; He, J.W.; Fu, W.Z.; Hu, Y.Q.; et al. Polymorphisms in Wnt Signaling Pathway Genes Are Associated with Peak Bone Mineral Density, Lean Mass, and Fat Mass in Chinese Male Nuclear Families. Osteoporos. Int. 2016, 27, 1805–1815. [Google Scholar] [CrossRef]
- Zhao, F.; Gao, L.-H.; Li, S.-S.; Wei, Z.-Y.; Fu, W.-Z.; He, J.-W.; Liu, Y.-J.; Hu, Y.-Q.; Dong, J.; Zhang, Z.-L. Association between SNPs and Haplotypes in the METTL21C Gene and Peak Bone Mineral Density and Body Composition in Chinese Male Nuclear Families. J. Bone Miner. Metab. 2017, 35, 437–447. [Google Scholar] [CrossRef] [PubMed]
- He, J.W.; Yue, H.; Hu, W.W.; Hu, Y.Q.; Zhang, Z.L. Contribution of the Sclerostin Domain-Containing Protein 1 (SOSTDC1) Gene to Normal Variation of Peak Bone Mineral Density in Chinese Women and Men. J. Bone Miner. Metab. 2011, 29, 571–581. [Google Scholar] [CrossRef] [PubMed]
- Chesi, A.; Wagley, Y.; Johnson, M.E.; Manduchi, E.; Su, C.; Lu, S.; Leonard, M.E.; Hodge, K.M.; Pippin, J.A.; Hankenson, K.D.; et al. Genome-Scale Capture C Promoter Interactions Implicate Effector Genes at GWAS Loci for Bone Mineral Density. Nat. Commun. 2019, 10, 1260. [Google Scholar] [CrossRef] [PubMed]
- Pippin, J.A.; Chesi, A.; Wagley, Y.; Su, C.; Pahl, M.C.; Hodge, K.M.; Johnson, M.E.; Wells, A.D.; Hankenson, K.D.; Grant, S.F.A. CRISPR-Cas9–Mediated Genome Editing Confirms EPDR1 as an Effector Gene at the BMD GWAS-Implicated ‘STARD3NL’ Locus. JBMR Plus 2021, 5, e10531. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.F.; Tobias, J.H.; Duncan, E.; Evans, D.M.; Eriksson, J.; Paternoster, L.; Yerges-Armstrong, L.M.; Lehtimäki, T.; Bergström, U.; Kähönen, M.; et al. WNT16 Influences Bone Mineral Density, Cortical Bone Thickness, Bone Strength, and Osteoporotic Fracture Risk. PLoS Genet. 2012, 8, e1002745. [Google Scholar] [CrossRef] [PubMed]
- Mullin, B.H.; Walsh, J.P.; Zheng, H.F.; Brown, S.J.; Surdulescu, G.L.; Curtis, C.; Breen, G.; Dudbridge, F.; Richards, J.B.; Spector, T.D.; et al. Genome-Wide Association Study Using Family-Based Cohorts Identifies the WLS and CCDC170/ESR1 Loci as Associated with Bone Mineral Density. BMC Genom. 2016, 17, 136. [Google Scholar] [CrossRef] [PubMed]
- Luo, F.; Xie, Y.; Chen, H.; Huang, J.; Li, C.; Chen, L.; Yang, J.; Su, N. Fgfr1 Deficiency in Osteocytes Leads to Increased Bone Mass by Enhancing Wnt/β-Catenin Signaling. Bone 2023, 174, 116817. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akhiiarova, K.; Khusainova, R.; Minniakhmetov, I.; Mokrysheva, N.; Tyurin, A. Peak Bone Mass Formation: Modern View of the Problem. Biomedicines 2023, 11, 2982. https://doi.org/10.3390/biomedicines11112982
Akhiiarova K, Khusainova R, Minniakhmetov I, Mokrysheva N, Tyurin A. Peak Bone Mass Formation: Modern View of the Problem. Biomedicines. 2023; 11(11):2982. https://doi.org/10.3390/biomedicines11112982
Chicago/Turabian StyleAkhiiarova, Karina, Rita Khusainova, Ildar Minniakhmetov, Natalia Mokrysheva, and Anton Tyurin. 2023. "Peak Bone Mass Formation: Modern View of the Problem" Biomedicines 11, no. 11: 2982. https://doi.org/10.3390/biomedicines11112982
APA StyleAkhiiarova, K., Khusainova, R., Minniakhmetov, I., Mokrysheva, N., & Tyurin, A. (2023). Peak Bone Mass Formation: Modern View of the Problem. Biomedicines, 11(11), 2982. https://doi.org/10.3390/biomedicines11112982