
Production and Functional Evaluation of Anti-Loxosceles Sera Raised by Immunizations of Rabbits with Mutated Recombinant Phospholipases-D

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Venom Solutions
2.2. Animals
2.3. Production and Characterization of Recombinant Phospholipases D (PLDs)
2.3.1. Recombinant Expression and Purification
2.3.2. Sphingomyelinase Activity
2.3.3. In Vivo Dermonecrosis
2.4. Production of Sera and Neutralization Assays
2.4.1. Immunization Protocols
2.4.2. Hematological and Biochemical Parameters
2.4.3. Clinical Follow-Up of Immunized Rabbits
2.4.4. Immunoassays
2.4.5. Neutralization Assays
2.4.6. Histological Analysis
3. Results
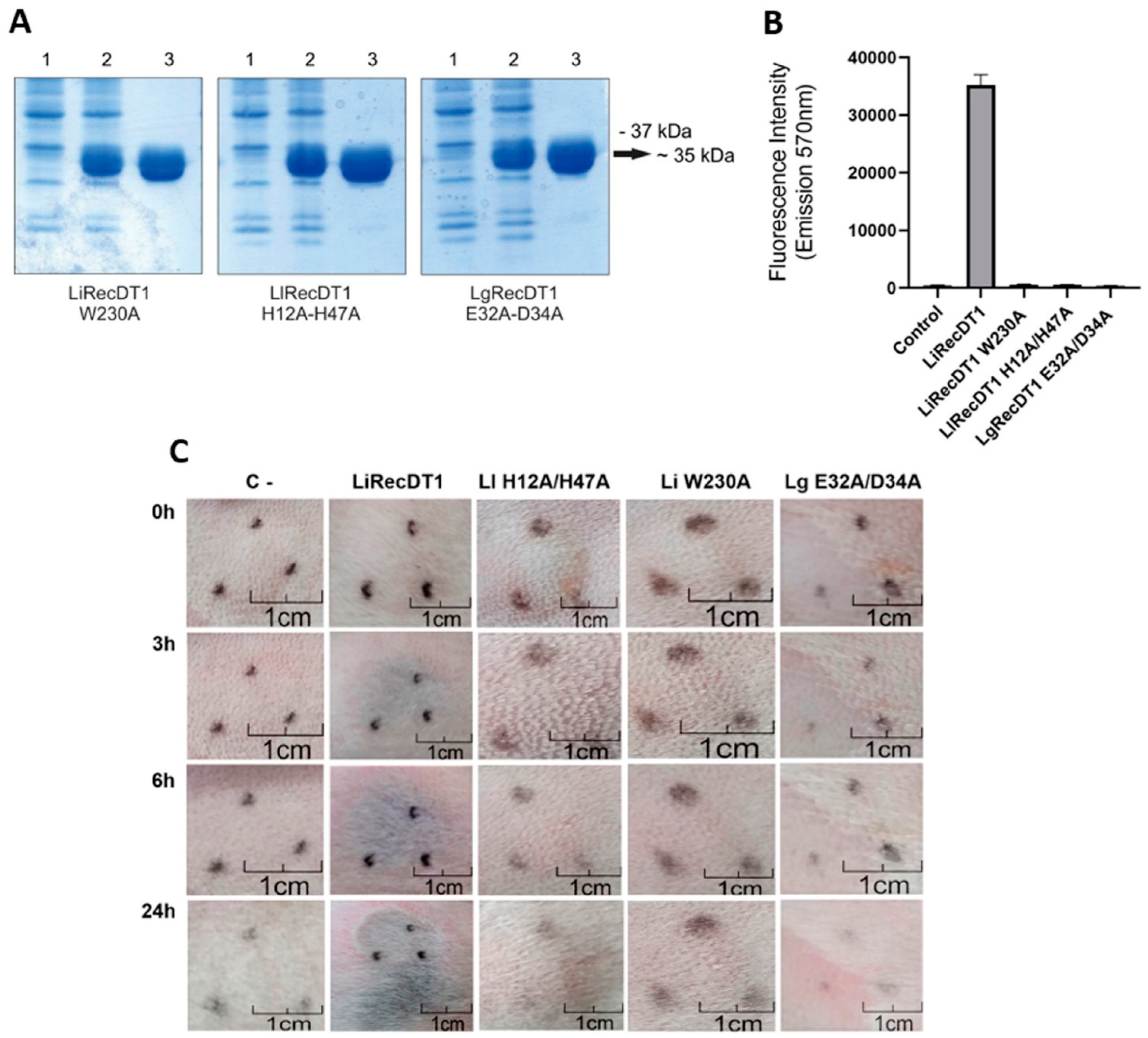
3.1. Mutated Recombinant PLDs Are Devoid of Enzymatic and Dermonecrotic Activities
3.2. Sera Production: Effects of Immunizations
3.3. Evaluation of the Produced Sera
3.4. Neutralization of In Vivo Toxicity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gremski, L.H.; Trevisan-Silva, D.; Ferrer, V.P.; Matsubara, F.H.; Meissner, G.O.; Wille, A.C.M.; Vuitika, L.; Dias-Lopes, C.; Ullah, A.; De Moraes, F.R.; et al. Recent advances in the understanding of brown spider venoms: From the biology of spiders to the molecular mechanisms of toxins. Toxicon 2014, 83, 91–120. [Google Scholar] [CrossRef] [PubMed]
- Lopes, P.H.; Murakami, M.T.; Portaro, F.C.V.; Mesquita Pasqualoto, K.F.; van den Berg, C.; Tambourgi, D.V. Targeting Loxosceles spider Sphingomyelinase D with small-molecule inhibitors as a potential therapeutic approach for loxoscelism. J. Enzyme Inhib. Med. Chem. 2019, 34, 310–321. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, C.F.B.; Vilela, A.; Coura, L.A.M.; Rodrigues, F.T.G.; Nagem, R.A.P.; Chávez-Olortegui, C.; Maioli, T.U.; Felicori, L.F. Protective antibodies against a sphingomyelinase D from Loxosceles intermedia spider venom elicited in mice with different genetic background. Vaccine 2016, 34, 3828–3834. [Google Scholar] [CrossRef] [PubMed]
- Grashof, D.; Zdenek, C.N.; Dobson, J.S.; Youngman, N.J.; Coimbra, F.; Benard-Valle, M.; Alagon, A.; Fry, B.G. A web of coagulotoxicity: Failure of antivenom to neutralize the destructive (non-clotting) fibrinogenolytic activity of loxosceles and sicarius spider venoms. Toxins 2020, 12, 91. [Google Scholar] [CrossRef] [PubMed]
- Isbister, G.K.; Fan, H.W. Spider bite. Lancet 2011, 378, 2039–2047. [Google Scholar] [CrossRef]
- Malaque, C.M.S.; Santoro, M.L.; Cardoso, J.L.C.; Conde, M.R.; Novaes, C.T.G.; Risk, J.Y.; França, F.O.S.; de Medeiros, C.R.; Fan, H.W. Clinical picture and laboratorial evaluation in human loxoscelism. Toxicon 2011, 58, 664–671. [Google Scholar] [CrossRef]
- Dragulev, B.; Bao, Y.; Ramos-Cerrillo, B.; Vazquez, H.; Olvera, A.; Stock, R.; Algaron, A.; Fox, J.W. Upregulation of IL-6, IL-8, CXCL1, and CXCL2 dominates gene expression in human fibroblast cells exposed to Loxosceles reclusa sphingomyelinase D: Insights into spider venom dermonecrosis. J. Investig. Dermatol. 2007, 127, 1264–1266. [Google Scholar] [CrossRef]
- Vetter, R.S. The distribution of brown recluse spiders in the southeastern quadrant of the United States in relation to loxoscelism diagnoses. South. Med. J. 2009, 102, 518–522. [Google Scholar] [CrossRef]
- Nguyen, N.; Pandey, M. Loxoscelism: Cutaneous and Hematologic Manifestations. Adv. Hematol. 2019, 2019, 4091278. [Google Scholar] [CrossRef]
- Gremski, L.H.; Matsubara, F.H.; da Justa, H.C.; Schemczssen-Graeff, Z.; Baldissera, A.B.; de Caires Schluga, P.H.; de Oliveira Leite, I.; Boia-Ferreira, M.; Wille, A.C.M.; Senff-Ribeiro, A.; et al. Brown spider venom toxins: What are the functions of astacins, serine proteases, hyaluronidases, allergens, TCTP, serpins and knottins? J. Venom. Anim. Toxins Incl. Trop. Dis. 2021, 27, e20200188. [Google Scholar] [CrossRef]
- Dantas, A.E.; Carmo, A.O.; Horta, C.C.R.; Leal, H.G.; Oliveira-Mendes, B.B.R.; Martins, A.P.V.; Chávez-Olórtegui, C.; Kalapothakis, E. Description of Loxtox protein family and identification of a new group of Phospholipases D from Loxosceles similis venom gland. Toxicon 2016, 120, 97–106. [Google Scholar] [CrossRef]
- Medina-Santos, R.; Guerra-Duarte, C.; de Almeida Lima, S.; Costal-Oliveira, F.; Alves de Aquino, P.; Oliveira do Carmo, A.; Ferreyra, C.B.; Gonzalez-Kozlova, E.E.; Kalapothakis, E.; Chávez-Olórtegui, C. Diversity of astacin-like metalloproteases identified by transcriptomic analysis in Peruvian Loxosceles laeta spider venom and in vitro activity characterization. Biochimie 2019, 167, 81–92. [Google Scholar] [CrossRef]
- Fingermann, M.; de Roodt, A.R.; Cascone, O.; Miranda, M.V. Biotechnological potential of Phospholipase D for Loxosceles antivenom development. Toxicon X 2020, 6, 100036. [Google Scholar] [CrossRef]
- Gremski, L.H.; Da Justa, H.C.; Da Silva, T.P.; Polli, N.L.C.; Antunes, B.C.; Minozzo, J.C.; Wille, A.C.M.; Senff-Ribeiro, A.; Arni, R.K.; Veiga, S.S. Forty years of the description of brown spider venom phospholipases-D. Toxins 2020, 12, 164. [Google Scholar] [CrossRef]
- Andrade, S.A.D.; Murakami, M.T.; Cavalcante, D.P.; Arni, R.K.; Tambourgi, D.V. Kinetic and mechanistic characterization of the Sphingomyelinases D from Loxosceles intermedia spider venom. Toxicon 2006, 47, 380–386. [Google Scholar] [CrossRef]
- Chaim, O.M.; Sade, Y.B.; Da Silveira, R.B.; Toma, L.; Kalapothakis, E.; Chávez-Olórtegui, C.; Mangili, O.C.; Gremski, W.; Von Dietrich, C.P.; Nader, H.B.; et al. Brown spider dermonecrotic toxin directly induces nephrotoxicity. Toxicol. Appl. Pharmacol. 2006, 211, 64–77. [Google Scholar] [CrossRef]
- Da Silveira, R.B.; Pigozzo, R.B.; Chaim, O.M.; Appel, M.H.; Silva, D.T.; Dreyfuss, J.L.; Toma, L.; Dietrich, C.P.; Nader, H.B.; Veiga, S.S.; et al. Two novel dermonecrotic toxins LiRecDT4 and LiRecDT5 from Brown spider (Loxosceles intermedia) venom: From cloning to functional characterization. Biochimie 2007, 89, 289–300. [Google Scholar] [CrossRef]
- Appel, M.H.; da Silveira, R.B.; Chaim, O.M.; Paludo, K.S.; Silva, D.T.; Chaves, D.M.; da Silva, P.H.; Mangili, O.C.; Senff-Ribeiro, A.; Gremski, W.; et al. Identification, cloning and functional characterization of a novel dermonecrotic toxin (phospholipase D) from brown spider (Loxosceles intermedia) venom. Biochim. Biophys. Acta Gen. Subj. 2008, 1780, 167–178. [Google Scholar] [CrossRef]
- Chaves-Moreira, D.; Chaim, O.M.; Sade, Y.B.; Paludo, K.S.; Gremski, L.H.; Donatti, L.; De Moura, J.; Mangili, O.C.; Gremski, W.; Da Silveira, R.B.; et al. Identification of a direct hemolytic effect dependent on the catalytic activity induced by phospholipase-D (dermonecrotic toxin) from brown spider venom. J. Cell Biochem. 2009, 107, 655–666. [Google Scholar] [CrossRef]
- Murakami, M.T.; Fernandes-Pedrosa, M.F.; Tambourgi, D.V.; Arni, R.K. Structural basis for metal ion coordination and the catalytic mechanism of sphingomyelinases D. J. Biol. Chem. 2005, 280, 13658–13664. [Google Scholar] [CrossRef]
- De Giuseppe, P.O.; Ullah, A.; Silva, D.T.; Gremski, L.H.; Wille, A.C.M.; Chaves Moreira, D.; Ribeiro, A.S.; Chaim, O.M.; Murakami, M.T.; Veiga, S.S.; et al. Structure of a novel class II phospholipase D: Catalytic cleft is modified by a disulphide bridge. Biochem. Biophys. Res. Commun. 2011, 409, 622–627. [Google Scholar] [CrossRef] [PubMed]
- Vuitika, L.; Chaves-Moreira, D.; Caruso, I.; Lima, M.A.; Matsubara, F.H.; Murakami, M.T.; Takahashi, H.K.; Toledo, M.S.; Coronado, M.A.; Nader, H.B.; et al. Active site mapping of Loxosceles phospholipases D: Biochemical and biological features. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2016, 1861, 970–979. [Google Scholar] [CrossRef] [PubMed]
- Masood, R.; Ullah, K.; Ali, H.; Ali, I.; Betzel, C.; Ullah, A. Spider’s venom phospholipases D: A structural review. Int. J. Biol. Macromol. 2018, 107, 1054–1065. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, T.P.; de Castro, F.J.; Vuitika, L.; Polli, N.L.C.; Antunes, B.C.; Bóia-Ferreira, M.; Minozzo, J.C.; Mariutti, R.B.; Matsubara, F.H.; Arni, R.K.; et al. Brown Spiders’ Phospholipases-D with Potential Therapeutic Applications: Functional Assessment of Mutant Isoforms. Biomedicines 2021, 9, 320. [Google Scholar] [CrossRef] [PubMed]
- Polli, N.L.C.; da Justa, H.C.; Antunes, B.C.; da Silva, T.P.; Dittrich, R.L.; de Souza, G.S.; Wille, A.C.M.; Matsubara, F.H.; Minozzo, J.C.; Mariutti, R.B.; et al. A protective vaccine against the toxic activities following Brown spider accidents based on recombinant mutated phospholipases D as antigens. Int. J. Biol. Macromol. 2021, 192, 757–770. [Google Scholar] [CrossRef] [PubMed]
- Karim-Silva, S.; Becker-Finco, A.; Jiacomini, I.G.; Boursin, F.; Leroy, A.; Noiray, M.; de Moura, J.; Aubrey, N.; Billiald, P.; Alvarenga, L.M. Loxoscelism: Advances and challenges in the design of antibody fragments with therapeutic potential. Toxins 2020, 12, 256. [Google Scholar] [CrossRef]
- Pauli, I.; Minozzo, J.C.; Henrique da Silva, P.; Chaim, O.M.; Veiga, S.S. Analysis of therapeutic benefits of antivenin at different time intervals after experimental envenomation in rabbits by venom of the brown spider (Loxosceles intermedia). Toxicon 2009, 53, 660–671. [Google Scholar] [CrossRef]
- De Almeida Lima, S.; Guerra-Duarte, C.; Costal-Oliveira, F.; Mendes, T.M.; Luís, L.F.; Oliveira, D.; de Avila, R.A.M.; Ferrer, V.P.; Trevisan-Silva, D.; Veiga, S.S.; et al. Recombinant protein containing B-cell epitopes of different Loxosceles spider toxins generates neutralizing antibodies in immunized rabbits. Front. Immunol. 2018, 9, 653. [Google Scholar] [CrossRef]
- Bermúdez-Méndez, E.; Fuglsang-Madsen, A.; Føns, S.; Lomonte, B.; Gutiérrez, J.M.; Laustsen, A.H. Innovative immunization strategies for antivenom development. Toxins 2018, 10, 452. [Google Scholar] [CrossRef]
- Calabria, P.A.L.; Shimokava-Falcao, L.H.A.L.; Colombini, M.; Moura-da-Silva, A.M.; Barbaro, K.C.; Faquim-Mauro, E.L.; Magalhaes, G.S. Design and production of a recombinant hybrid toxin to raise protective antibodies against loxosceles spider venom. Toxins 2019, 11, 108. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Harlow, E.; Lane, D. Antibodies: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Long Island, NY, USA, 1988. [Google Scholar]
- Wintrobe, M.M.; Greer, J.P. Wintrobe’s Clinical Hematology, 13th ed.; Greer, J.P., Ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2014; ISBN 9781451172683. [Google Scholar]
- McPherson, R.A.; Pincus, M.R.; Henry, J.B. Henry’s Clinical Diagnosis and Management by Laboratory Methods, 23rd ed.; Sauders Elsevier: Philadelphia, PA, USA, 2017; ISBN 978-0-323-41315-2. [Google Scholar]
- Escalas—Úlceras Fóra. Available online: https://ulcerasfora.sergas.gal/Informacion/Escala-visual-del-eritema-(EVE) (accessed on 11 October 2022).
- Figueiredo, L.F.M.; Dias-Lopes, C.; Alvarenga, L.M.; Mendes, T.M.; Machado-De-Ávila, R.A.; McCormack, J.; Minozzo, J.C.; Kalapothakis, E.; Chávez-Olórtegui, C. Innovative immunization protocols using chimeric recombinant protein for the production of polyspecific loxoscelic antivenom in horses. Toxicon 2014, 86, 59–67. [Google Scholar] [CrossRef]
- Angulo, Y.; Estrada, R.; Gutiérrez, J.M. Clinical and Laboratory Alterations in Horses during Immunization with Snake Venoms for the Production of Polyvalent (Crotalinae) Antivenom. Toxicon 1997, 35, 81–90. [Google Scholar] [CrossRef]
- Garcia-Navarro, C.E.K. Manual de Hematologia Veterinária, 2nd ed.; Varela: São Paulo, Brazil, 2005; ISBN 8585519088. [Google Scholar]
- Parra, A.C.; Távora, J.P.F.; Ferreira, R.A.; Betiol, P.S.; Birgel, E.H. Alterações hematológicas durante a imunização e após a sangria e plasmaferese em equinos de produção de soro hiperimune anticotálico. Ciência Anim. Bras. 2009, 10, 1225–1230. [Google Scholar]
- Deeb, B.J.; DiGiacomo, R.F.; Kunz, L.L.; Stewart, J.L. Comparison of Freund’s and Ribi adjuvants for inducing antibodies to the synthetic antigen (TG)-AL in rabbits. J. Immunol. Methods 1992, 152, 105–113. [Google Scholar] [CrossRef]
- Laustsen, A.H.; Solà, M.; Jappe, E.C.; Oscoz, S.; Lauridsen, L.P.; Engmark, M. Biotechnological trends in spider and scorpion antivenom development. Toxins 2016, 8, 226. [Google Scholar] [CrossRef]
- de Souza, A.L.; Malaque, C.M.; Sztajnbok, J.; Romano, C.C.; Duarte, A.J.; Seguro, A.C. Loxosceles venom-induced cytokine activation, hemolysis, and acute kidney injury. Toxicon 2008, 51, 151–156. [Google Scholar] [CrossRef]
- Cox, J.C.; Coulter, A.R. Adjuvants—A classification and review of their modes of action. Vaccine 1997, 15, 248–256. [Google Scholar] [CrossRef]
- Abbas, A.; Lichtman, A.; Pillai, S. Cellular and Molecular Immunology, 9th ed.; Elsevier: Amsterdam, The Netherlands, 2016; ISBN 9780323479783. [Google Scholar]
- Meredith, A.; Rayment, L. Liver disease in rabbits. In Pract. 2000, 9, 146–152. [Google Scholar] [CrossRef]
- De Oliveira Christoff, A.; de Oliveira, A.; Chaim, O.M.; Lugarini, D.; Bastos Pereira, A.L.; Paludo, K.S.; Queiroz Telles, J.E.; Bracht, A.; Veiga, S.S.; Acco, A. Effects of the venom and the dermonecrotic toxin LiRecDT1 of Loxosceles intermedia in the rat liver. Toxicon 2008, 52, 695–704. [Google Scholar] [CrossRef]
- Carpenter, J.W.; Marion, C.J. Exotic Animal Formulary, 5th ed.; Elsevier: Amsterdam, The Netherlands, 2018; ISBN 978-0-323-44450-7. [Google Scholar]
- Padula, A.M.; Winkel, K.D. Antivenom production in the alpaca (Vicugna pacos): Physiological and antibody responses to monovalent and polyvalent immunisation with Australian elapid venoms. Small Rumin. Res. 2016, 141, 63–69. [Google Scholar] [CrossRef]
- Bennett, B.; Check, I.J.; Olsen, M.R.; Hunter, R.L. A comparison of commercially available adjuvants for use in research. J. Immunol. Methods 1992, 153, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Pay, T.W.F.; Hingley, P.J. Correlation of 140S antigen dose with the serum neutralizing antibody response and the level of protection induced in cattle by foot-and-mouth disease vaccines. Vaccine 1987, 5, 60–64. [Google Scholar] [CrossRef] [PubMed]
- Katare, Y.K.; Muthukumaran, T.; Panda, A.K. Influence of particle size, antigen load, dose and additional adjuvant on the immune response from antigen loaded PLA microparticles. Int. J. Pharm. 2005, 301, 149–160. [Google Scholar] [CrossRef] [PubMed]
- Romstad, A.B.; Reitan, L.J.; Midtlyng, P.; Gravningen, K.; Evensen, Ø. Antibody responses correlate with antigen dose and in vivo protection for oil-adjuvanted, experimental furunculosis (Aeromonas salmonicida subsp. salmonicida) vaccines in Atlantic salmon (Salmo salar L.) and can be used for batch potency testing of vaccin. Vaccine 2013, 31, 791–796. [Google Scholar] [CrossRef]
- Barbaro, K.C.; Knysak, I.; Martins, R.; Hogan, C.; Winkel, K. Enzymatic characterization, antigenic cross-reactivity and neutralization of dermonecrotic activity of five Loxosceles spider venoms of medical importance in the Americas. Toxicon 2005, 45, 489–499. [Google Scholar] [CrossRef]
- Leal, H.G.; de Oliveira Mendes, B.B.R.; Horta, C.C.R.; Pereira, N.B.; Ferreira, D.S.M.; da Silva, T.S.; Biscoto, G.L.; Kalapothakis, Y.; Machado de Avila, R.A.; Chávez-Olórtegui, C.; et al. Molecular cloning and functional characterization of recombinant Loxtox from Loxosceles similis venom. Int. J. Biol. Macromol. 2020, 164, 1112–1123. [Google Scholar] [CrossRef]
- Magalhães, G.S.; Caporrino, M.C.; Della-Casa, M.S.; Kimura, L.F.; Prezotto-Neto, J.P.; Fukuda, D.A.; Portes-Junior, J.A.; Neves-Ferreira, A.G.C.; Santoro, M.L.; Barbaro, K.C. Cloning, expression and characterization of a phospholipase D from Loxosceles gaucho venom gland. Biochimie 2013, 95, 1773–1783. [Google Scholar] [CrossRef]
- Tavares, F.L.; Peichoto, M.E.; Marcelino, J.R.; Barbaro, K.C.; Cirillo, M.C.; Santoro, M.L.; Sano-Martins, I.S. Platelet participation in the pathogenesis of dermonecrosis induced by Loxosceles gaucho venom. Hum. Exp. Toxicol. 2016, 35, 666–676. [Google Scholar] [CrossRef]
- Futrell, J.M. Loxoscelism. Am. J. Med. Sci. 1992, 304, 261–267. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group Name | Antigen | 1st Injection | 2nd Injection | 3rd Injection | 4th Injection |
|---|---|---|---|---|---|
| AV | Venom pool * | 7.5 µg of pool + CFA | 10 µg of pool + IFA | 10 µg of pool + IFA | 15 µg of pool + IFA |
| REC | Recombinant proteins’ pool ** | 100 µg of pool + CFA | 200 µg of pool + IFA | 400 µg of pool + IFA | 800 µg of pool + IFA |
| MIX | Venom pool * + Recombinant proteins’ pool ** | 2.5 µg of venom pool + 97.5 µg of recombinant protein’s pool + CFA | 2.5 µg of venom pool + 197.5 µg of recombinant protein’s pool + IFA | 2.5 µg of venom pool + 397.5 µg of recombinant protein’s pool + IFA | 2.5 µg of venom pool + 797.5 µg of recombinant protein’s pool + IFA |
| C | Negative control | 0.85% saline + CFA | 0.85% saline + IFA | 0.85% saline + IFA | 0.85% saline + IFA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antunes, B.C.; Polli, N.L.C.; Schluga, P.H.d.C.; Silva, T.P.d.; Wille, A.C.M.; Locatelli-Dittrich, R.; Souza, G.S.d.; Matsubara, F.H.; Minozzo, J.C.; Senff-Ribeiro, A.; et al. Production and Functional Evaluation of Anti-Loxosceles Sera Raised by Immunizations of Rabbits with Mutated Recombinant Phospholipases-D. Biomedicines 2023, 11, 79. https://doi.org/10.3390/biomedicines11010079
Antunes BC, Polli NLC, Schluga PHdC, Silva TPd, Wille ACM, Locatelli-Dittrich R, Souza GSd, Matsubara FH, Minozzo JC, Senff-Ribeiro A, et al. Production and Functional Evaluation of Anti-Loxosceles Sera Raised by Immunizations of Rabbits with Mutated Recombinant Phospholipases-D. Biomedicines. 2023; 11(1):79. https://doi.org/10.3390/biomedicines11010079
Chicago/Turabian StyleAntunes, Bruno Cesar, Nayanne Louise Costacurta Polli, Pedro Henrique de Caires Schluga, Thais Pereira da Silva, Ana Carolina Martins Wille, Rosangela Locatelli-Dittrich, Giovana Scuissiatto de Souza, Fernando Hitomi Matsubara, João Carlos Minozzo, Andrea Senff-Ribeiro, and et al. 2023. "Production and Functional Evaluation of Anti-Loxosceles Sera Raised by Immunizations of Rabbits with Mutated Recombinant Phospholipases-D" Biomedicines 11, no. 1: 79. https://doi.org/10.3390/biomedicines11010079
APA StyleAntunes, B. C., Polli, N. L. C., Schluga, P. H. d. C., Silva, T. P. d., Wille, A. C. M., Locatelli-Dittrich, R., Souza, G. S. d., Matsubara, F. H., Minozzo, J. C., Senff-Ribeiro, A., Gremski, L. H., & Veiga, S. S. (2023). Production and Functional Evaluation of Anti-Loxosceles Sera Raised by Immunizations of Rabbits with Mutated Recombinant Phospholipases-D. Biomedicines, 11(1), 79. https://doi.org/10.3390/biomedicines11010079