
Gamma-Synuclein Dysfunction Causes Autoantibody Formation in Glaucoma Patients and Dysregulation of Intraocular Pressure in Mice


Abstract
1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Detection of Autoantibodies to γ-Synuclein
2.3. Experimental Animals
3. Results
3.1. Autoantibodies to γ-Synuclein Were Found in the Blood Serum of Some Patients with Glaucoma
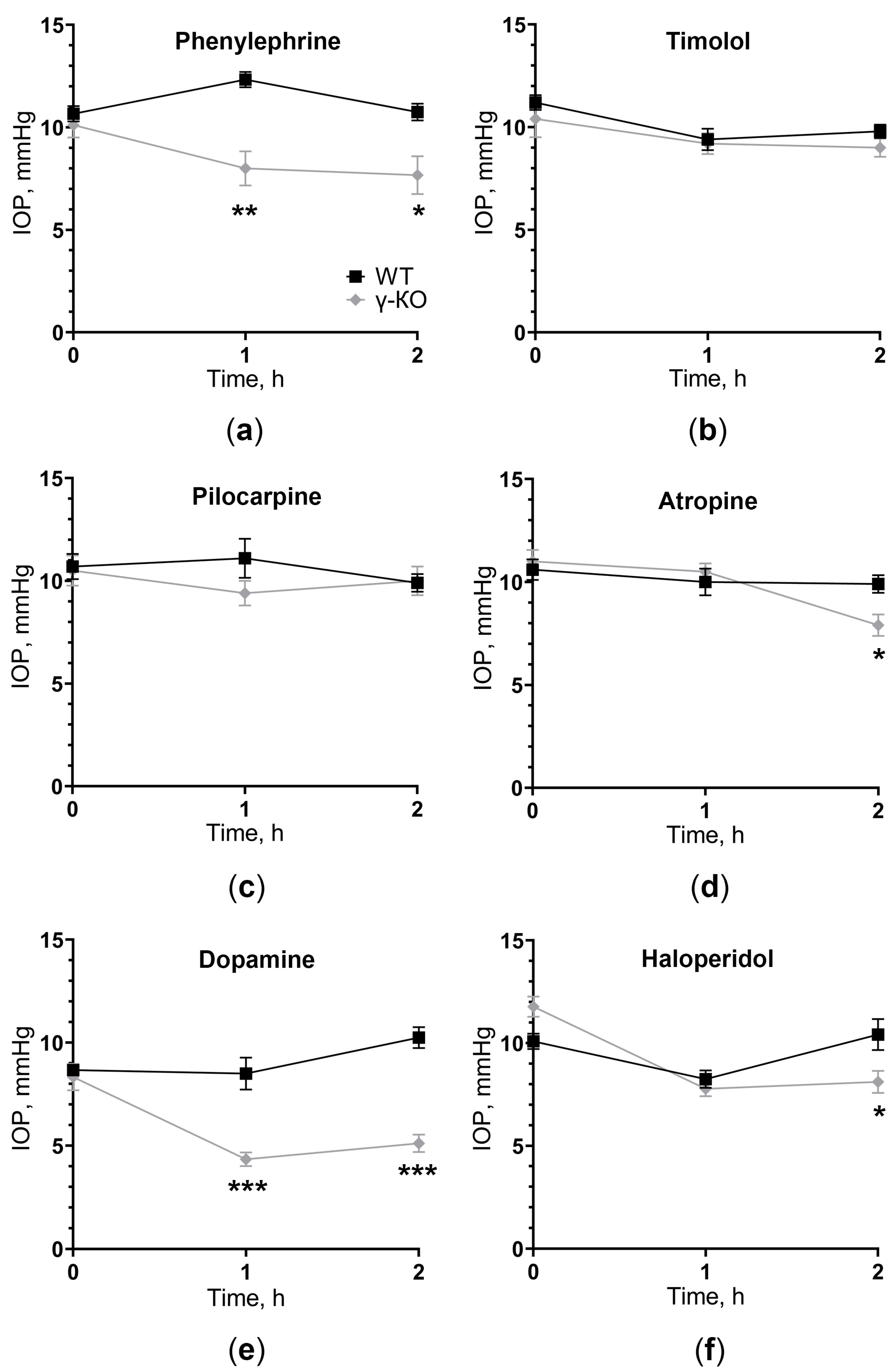
3.2. Loss of γ-Synuclein Function Led to Dysregulation of IOP in Mice
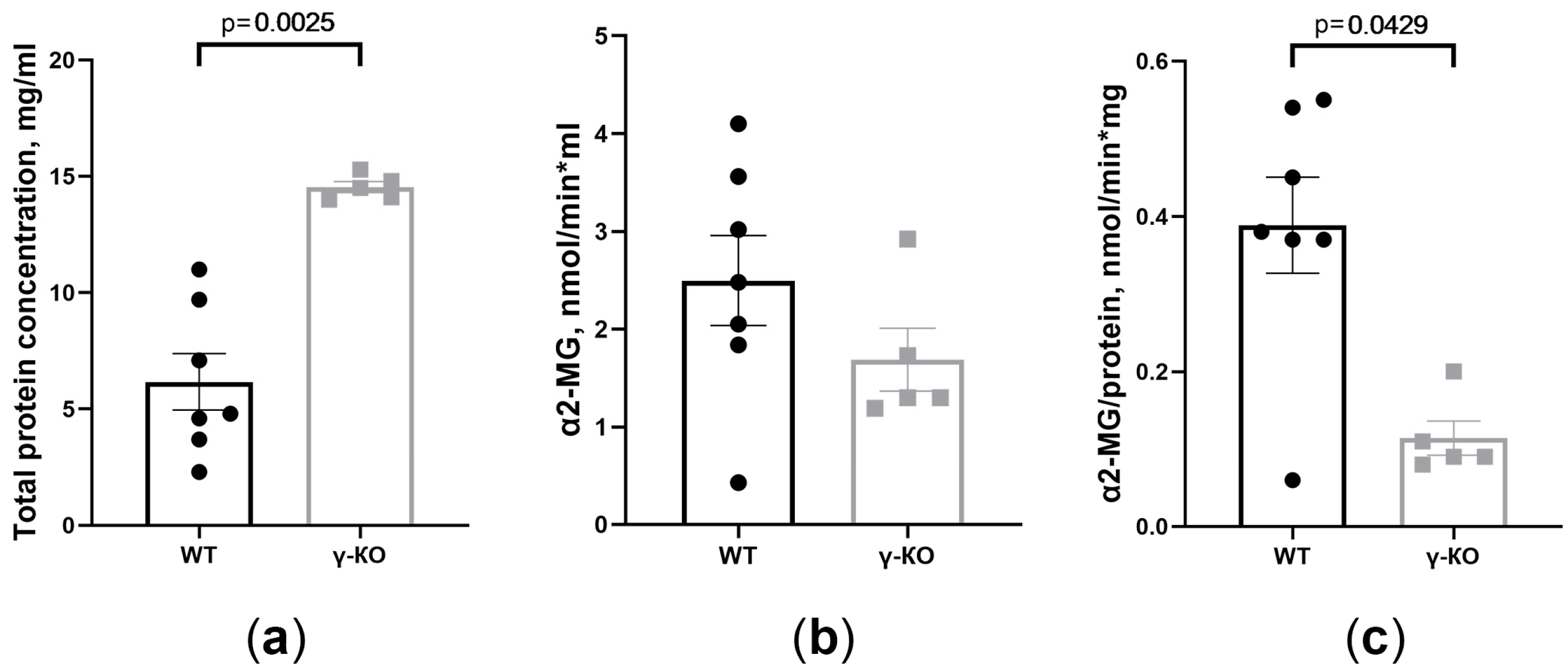
3.3. The Total Protein Concentration and the Activity of the α2-Macroglobulin Changed in the Tear Fluid of γ-KO Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tuulonen, A.; Airaksinen, P.J.; Erola, E.; Forsman, E.; Friberg, K.; Kaila, M.; Klemetti, A.; Makela, M.; Oskala, P.; Puska, P.; et al. The Finnish evidence-based guideline for open-angle glaucoma. Acta Ophthalmol. Scand. 2003, 81, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Ophthalmol, B.J. European Glaucoma Society Terminology and Guidelines for Glaucoma, 4th Edition—Chapter 2: Classification and terminologySupported by the EGS Foundation: Part 1: Foreword; Introduction; Glossary; Chapter 2 Classification and Terminology. Br. J. Ophthalmol. 2017, 101, 73–127. [Google Scholar] [CrossRef]
- Quigley, H.A.; Broman, A.T. The number of people with glaucoma worldwide in 2010 and 2020. Br. J. Ophthalmol. 2006, 90, 262–267. [Google Scholar] [CrossRef] [PubMed]
- Friedman, D.S.; Wilson, M.R.; Liebmann, J.M.; Fechtner, R.D.; Weinreb, R.N. An evidence-based assessment of risk factors for the progression of ocular hypertension and glaucoma. Am. J. Ophthalmol. 2004, 138, S19-312004. [Google Scholar] [CrossRef]
- Adornetto, A.; Russo, R.; Parisi, V. Neuroinflammation as a target for glaucoma therapy. Neural. Regen. Res. 2019, 14, 391–394. [Google Scholar] [CrossRef] [PubMed]
- Carelli, V.; La Morgia, C.; Ross-Cisneros, F.N.; Sadun, A.A. Optic neuropathies: The tip of the neurodegeneration iceberg. Hum. Mol. Genet. 2017, 26, R139-R1502017. [Google Scholar] [CrossRef]
- Jiang, S.; Kametani, M.; Chen, D.F. Adaptive Immunity: New Aspects of Pathogenesis Underlying Neurodegeneration in Glaucoma and Optic Neuropathy. Front. Immunol. 2020, 11, 65. [Google Scholar] [CrossRef] [PubMed]
- Bras, I.C.; Dominguez-Meijide, A.; Gerhardt, E.; Koss, D.; Lazaro, D.F.; Santos, P.I.; Vasili, E.; Xylaki, M.; Outeiro, T.F. Synucleinopathies: Where we are and where we need to go. J. Neurochem. 2020, 153, 433–454. [Google Scholar] [CrossRef]
- Surguchov, A. Intracellular Dynamics of Synucleins: “Here, There and Everywhere”. Int. Rev. Cell. Mol. Biol. 2015, 320, 103–169. [Google Scholar] [CrossRef]
- Carnazza, K.E.; Komer, L.E.; Xie, Y.X.; Pineda, A.; Briano, J.A.; Gao, V.; Na, Y.; Ramlall, T.; Buchman, V.L.; Eliezer, D.; et al. Synaptic vesicle binding of alpha-synuclein is modulated by beta- and gamma-synucleins. Cell. Rep. 2022, 39, 110675. [Google Scholar] [CrossRef]
- Burre, J.; Sharma, M.; Sudhof, T.C. Cell Biology and Pathophysiology of alpha-Synuclein. Cold Spring Harb. Perspect. Med. 2018, 8, a024091. [Google Scholar] [CrossRef] [PubMed]
- Spillantini, M.G.; Schmidt, M.L.; Lee, V.M.; Trojanowski, J.Q.; Jakes, R.; Goedert, M. Alpha-synuclein in Lewy bodies. Nature 1997, 388, 839–840. [Google Scholar] [CrossRef] [PubMed]
- Ueda, K.; Fukushima, H.; Masliah, E.; Xia, Y.; Iwai, A.; Yoshimoto, M.; Otero, D.A.; Kondo, J.; Ihara, Y.; Saitoh, T. Molecular cloning of cDNA encoding an unrecognized component of amyloid in Alzheimer disease. Proc. Natl. Acad. Sci. USA 1993, 90, 11282–11286. [Google Scholar] [CrossRef] [PubMed]
- Ninkina, N.; Peters, O.; Millership, S.; Salem, H.; van der Putten, H.; Buchman, V.L. Gamma-synucleinopathy: Neurodegeneration associated with overexpression of the mouse protein. Hum. Mol. Genet. 2009, 18, 1779–1794. [Google Scholar] [CrossRef] [PubMed]
- Peters, O.M.; Millership, S.; Shelkovnikova, T.A.; Soto, I.; Keeling, L.; Hann, A.; Marsh-Armstrong, N.; Buchman, V.L.; Ninkina, N. Selective pattern of motor system damage in gamma-synuclein transgenic mice mirrors the respective pathology in amyotrophic lateral sclerosis. Neurobiol. Dis. 2012, 48, 124–131. [Google Scholar] [CrossRef]
- Peters, O.M.; Shelkovnikova, T.; Highley, J.R.; Cooper-Knock, J.; Hortobagyi, T.; Troakes, C.; Ninkina, N.; Buchman, V.L. Gamma-synuclein pathology in amyotrophic lateral sclerosis. Ann. Clin. Transl. Neurol. 2015, 2, 29–37. [Google Scholar] [CrossRef]
- Surgucheva, I.; Newell, K.L.; Burns, J.; Surguchov, A. New alpha- and gamma-synuclein immunopathological lesions in human brain. Acta Neuropathol. Commun. 2014, 2, 132. [Google Scholar] [CrossRef]
- Liu, H.; Liu, W.; Wu, Y.; Zhou, Y.; Xue, R.; Luo, C.; Wang, L.; Zhao, W.; Jiang, J.D.; Liu, J. Loss of epigenetic control of synuclein-gamma gene as a molecular indicator of metastasis in a wide range of human cancers. Cancer Res. 2005, 65, 7635–7643. [Google Scholar] [CrossRef]
- Surguchov, A. γ-Synuclein as a Cancer Biomarker: Viewpoint and New Approaches. Oncomedicine 2016, 1, 1–3. [Google Scholar] [CrossRef][Green Version]
- Surgucheva, I.; McMahan, B.; Ahmed, F.; Tomarev, S.; Wax, M.B.; Surguchov, A. Synucleins in glaucoma: Implication of gamma-synuclein in glaucomatous alterations in the optic nerve. J. Neurosci. Res. 2002, 68, 97–106. [Google Scholar] [CrossRef]
- Surgucheva, I.; Weisman, A.D.; Goldberg, J.L.; Shnyra, A.; Surguchov, A. Gamma-synuclein as a marker of retinal ganglion cells. Mol. Vis. 2008, 14, 1540–1548. [Google Scholar] [PubMed]
- Gold, M.; Pul, R.; Bach, J.P.; Stangel, M.; Dodel, R. Pathogenic and physiological autoantibodies in the central nervous system. Immunol. Rev. 2012, 248, 68–86. [Google Scholar] [CrossRef] [PubMed]
- Papachroni, K.; Ninkina, N.; Papapanagiotou, A.; Hadjigeorgiou, G.; Xiromerisiou, G.; Papadimitriou, A.; Kalofoutis, A.; Buchman, V.L. Autoantibodies to alpha-synuclein in inherited Parkinson’s disease. Neurochem 2007, 101, 749–756. [Google Scholar] [CrossRef] [PubMed]
- Roman, A.Y.; Kovrazhkina, E.A.; Razinskaya, O.D.; Kukharsky, M.S.; Maltsev, A.V.; Ovchinnikov, R.K.; Lytkina, O.A.; Smirnov, A.P.; Moskovtsev, A.A.; Borodina, Y.V.; et al. Detection of autoantibodies to potentially amyloidogenic protein, gamma-synuclein, in the serum of patients with amyotrophic lateral sclerosis and cerebral circulatory disorders. Dokl. Biochem. Biophys. 2017, 472, 64–67. [Google Scholar] [CrossRef] [PubMed]
- Bell, K.; Und Hohenstein-Blaul, N.V.T.; Teister, J.; Grus, F. Modulation of the Immune System for the Treatment of Glaucoma. Curr. Neuropharmacol. 2018, 16, 942–958. [Google Scholar] [CrossRef] [PubMed]
- Von Thun Und Hohenstein-Blaul, N.; Kunst, S.; Pfeiffer, N.; Grus, F.H. Biomarkers for glaucoma: From the lab to the clinic. Eye 2017, 31, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Bell, K.; Wilding, C.; Funke, S.; Perumal, N.; Beck, S.; Wolters, D.; Holz-Muller, J.; Pfeiffer, N.; Grus, F.H. Neuroprotective effects of antibodies on retinal ganglion cells in an adolescent retina organ culture. J. Neurochem. 2016, 139, 256–269. [Google Scholar] [CrossRef] [PubMed]
- Wilding, C.; Bell, K.; Beck, S.; Funke, S.; Pfeiffer, N.; Grus, F.H. gamma-Synuclein antibodies have neuroprotective potential on neuroretinal cells via proteins of the mitochondrial apoptosis pathway. PLoS ONE 2014, 9, e90737. [Google Scholar] [CrossRef]
- Ninkina, N.; Papachroni, K.; Robertson, D.C.; Schmidt, O.; Delaney, L.; O’Neill, F.; Court, F.; Rosenthal, A.; Fleetwood-Walker, S.M.; Davies, A.M.; et al. Neurons expressing the highest levels of gamma-synuclein are unaffected by targeted inactivation of the gene. Mol. Cell. Biol. 2003, 23, 8233–8245. [Google Scholar] [CrossRef]
- Anwar, S.; Peters, O.; Millership, S.; Ninkina, N.; Doig, N.; Connor-Robson, N.; Threlfell, S.; Kooner, G.; Deacon, R.M.; Bannerman, D.M.; et al. Functional alterations to the nigrostriatal system in mice lacking all three members of the synuclein family. J. Neurosci. 2011, 31, 7264–7274. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Bogdanov, V.; Kim, A.; Nodel, M.; Pavlenko, T.; Pavlova, E.; Blokhin, V.; Chesnokova, N.; Ugrumov, M. A Pilot Study of Changes in the Level of Catecholamines and the Activity of alpha-2-Macroglobulin in the Tear Fluid of Patients with Parkinson’s Disease and Parkinsonian Mice. Int. J. Mol. Sci. 2021, 22, 4736. [Google Scholar] [CrossRef] [PubMed]
- Maurage, C.A.; Ruchoux, M.M.; de Vos, R.; Surguchov, A.; Destee, A. Retinal involvement in dementia with Lewy bodies: A clue to hallucinations? Ann. Neurol. 2003, 54, 542–547. [Google Scholar] [CrossRef]
- Surguchev, A.; Surguchov, A. Conformational diseases: Looking into the eyes. Brain Res. Bull. 2010, 81, 12–24. [Google Scholar] [CrossRef] [PubMed]
- Grimaldi, A.; Brighi, C.; Peruzzi, G.; Ragozzino, D.; Bonanni, V.; Limatola, C.; Ruocco, G.; Di Angelantonio, S. Inflammation, neurodegeneration and protein aggregation in the retina as ocular biomarkers for Alzheimer’s disease in the 3xTg-AD mouse model. Cell Death Dis. 2018, 9, 685. [Google Scholar] [CrossRef] [PubMed]
- Leger, F.; Fernagut, P.O.; Canron, M.H.; Leoni, S.; Vital, C.; Tison, F.; Bezard, E.; Vital, A. Protein aggregation in the aging retina. J. Neuropathol. Exp. Neurol. 2011, 70, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Tapia, M.L.; Yeh, J.; He, R.C.; Pomerleu, D.; Lee, R.K. Differential Gamma-Synuclein Expression in Acute and Chronic Retinal Ganglion Cell Death in the Retina and Optic Nerve. Mol. Neurobiol. 2020, 57, 698–709. [Google Scholar] [CrossRef] [PubMed]
- Soto, I.; Oglesby, E.; Buckingham, B.P.; Son, J.L.; Roberson, E.D.; Steele, M.R.; Inman, D.M.; Vetter, M.L.; Horner, P.J.; Marsh-Armstrong, N. Retinal ganglion cells downregulate gene expression and lose their axons within the optic nerve head in a mouse glaucoma model. J. Neurosci. 2008, 28, 548–561. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Pescosolido, N.; Parisi, F.; Russo, P.; Buomprisco, G.; Nebbioso, M. Role of dopaminergic receptors in glaucomatous disease modulation. Biomed. Res. Int. 2013, 2013, 193048. [Google Scholar] [CrossRef]
- Oaks, A.W.; Sidhu, A. Synuclein modulation of monoamine transporters. FEBS Lett. 2011, 585, 1001–1006. [Google Scholar] [CrossRef]
- Kokhan, V.S.; Kokhan, T.Y.G.; Samsonova, A.N.; Fisenko, V.P.; Ustyugov, A.A.; Aliev, G. The Dopaminergic Dysfunction and Altered Working Memory Performance of Aging Mice Lacking Gamma-synuclein Gene. CNS Neurol. Disord. Drug Targets 2018, 17, 604–607. [Google Scholar] [CrossRef] [PubMed]
- Wersinger, C.; Sidhu, A. Partial regulation of serotonin transporter function by gamma-synuclein. Neurosci. Lett. 2009, 453, 157–161. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lei, S. Cross interaction of dopaminergic and adrenergic systems in neural modulation. Int. J. Physiol. Pathophysiol. Pharmacol. 2014, 6, 137–142. [Google Scholar] [PubMed]
- Schwahn, H.N.; Kaymak, H.; Schaeffel, F. Effects of atropine on refractive development, dopamine release, and slow retinal potentials in the chick. Vis. Neurosci. 2000, 17, 165–176. [Google Scholar] [CrossRef]
- Rehman, A.A.; Ahsan, H.; Khan, F.H. alpha-2-Macroglobulin: A physiological guardian. J. Cell. Physiol. 2013, 228, 1665–1675. [Google Scholar] [CrossRef]
- Cater, J.H.; Wilson, M.R.; Wyatt, A.R. Alpha-2-Macroglobulin, a Hypochlorite-Regulated Chaperone and Immune System Modulator. Oxid. Med. Cell. Longev. 2019, 2019, 5410657. [Google Scholar] [CrossRef]
- Wyatt, A.R.; Constantinescu, P.; Ecroyd, H.; Dobson, C.M.; Wilson, M.R.; Kumita, J.R.; Yerbury, J.J. Protease-activated alpha-2-macroglobulin can inhibit amyloid formation via two distinct mechanisms. FEBS Lett. 2013, 587, 398–403. [Google Scholar] [CrossRef]
- Inyushin, M.; Zayas-Santiago, A.; Rojas, L.; Kucheryavykh, Y.; Kucheryavykh, L. Platelet-generated amyloid beta peptides in Alzheimer’s disease and glaucoma. Histol. Histopathol. 2019, 34, 843–856. [Google Scholar] [CrossRef]
- Wang, L.; Mao, X. Role of Retinal Amyloid-beta in Neurodegenerative Diseases: Overlapping Mechanisms and Emerging Clinical Applications. Int. J. Mol. Sci. 2021, 22, 2360. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Patient Number | Gender | Age (Years) | Diagnosis | Autoantibodies to γ-Synuclein | Stage of the Disease | Surgical Treatment of Glaucoma |
|---|---|---|---|---|---|---|
| Control group | ||||||
| 1 | M | 44 | No glaucoma symptoms | - | NA | NA |
| 2 | F | 91 | No glaucoma symptoms | - | NA | NA |
| 3 | M | 70 | No glaucoma symptoms | - | NA | NA |
| 4 | M | 85 | No glaucoma symptoms | - | NA | NA |
| 5 | M | 64 | No glaucoma symptoms | - | NA | NA |
| 6 | M | 64 | No glaucoma symptoms | - | NA | NA |
| 7 | F | 80 | No glaucoma symptoms | - | NA | NA |
| 8 | F | 59 | No glaucoma symptoms | - | NA | NA |
| 9 | F | 48 | No glaucoma symptoms | - | NA | NA |
| 10 | F | 67 | No glaucoma symptoms | - | NA | NA |
| 11 | F | 63 | No glaucoma symptoms | - | NA | NA |
| 12 | M | 62 | No glaucoma symptoms | - | NA | NA |
| 13 | F | 67 | No glaucoma symptoms | - | NA | NA |
| Primary open-angle glaucoma | ||||||
| 14 | M | 80 | POAG | + | 4 OU | Yes |
| 15 | F | 78 | POAG | - | 3 OU | No |
| 16 | M | 76 | POAG | - | 5 OS; 2 OD | Yes |
| 17 | M | 78 | POAG | - | 2OU | Yes |
| 18 | F | 78 | POAG | - | 1 OU | No |
| 19 | M | 78 | POAG | - | 4 OS; 1 OD | No |
| 20 | F | 86 | POAG | - | 1 OU | No |
| 21 | F | 75 | POAG | - | 3 OU | Yes |
| 22 | M | 69 | POAG | - | 4OS; 3 OD | Yes |
| 23 | F | 59 | POAG | - | 1 OS glaucoma suspect; 1 OD | Yes |
| 24 | M | 76 | POAG | + | 3 OS; 1 OD | Yes |
| 25 | F | 69 | POAG | - | OU glaucoma suspect | No |
| 26 | M | 80 | POAG | + | 2 OS; 1 OD | Yes |
| 27 | M | 69 | POAG | - | 1 OS; 3 OD | Yes |
| 28 | F | 80 | POAG | - | 2 OU | No |
| 29 | M | 53 | POAG | - | 3 OU | Yes |
| 30 | M | 76 | POAG | + | 2 OS; 3 OD | Yes |
| 31 | F | 85 | POAG | - | 2 OS, OD anophthalmia | No |
| 32 | M | 64 | POAG | - | 3 OU | No |
| 33 | F | 68 | POAG | - | OU glaucoma suspect | No |
| 34 | F | 71 | POAG | - | 3 OS; 1 OD | No |
| 35 | M | 79 | POAG | - | 4 OS; 3 OD | Yes |
| 36 | F | 68 | POAG | - | 1 OS; 2 OD | Yes |
| 37 | F | 71 | POAG | - | 2 OU | Yes |
| 38 | M | 65 | POAG | + | 1 OS; 2 OD | No |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pavlenko, T.A.; Roman, A.Y.; Lytkina, O.A.; Pukaeva, N.E.; Everett, M.W.; Sukhanova, I.S.; Soldatov, V.O.; Davidova, N.G.; Chesnokova, N.B.; Ovchinnikov, R.K.; et al. Gamma-Synuclein Dysfunction Causes Autoantibody Formation in Glaucoma Patients and Dysregulation of Intraocular Pressure in Mice. Biomedicines 2023, 11, 60. https://doi.org/10.3390/biomedicines11010060
Pavlenko TA, Roman AY, Lytkina OA, Pukaeva NE, Everett MW, Sukhanova IS, Soldatov VO, Davidova NG, Chesnokova NB, Ovchinnikov RK, et al. Gamma-Synuclein Dysfunction Causes Autoantibody Formation in Glaucoma Patients and Dysregulation of Intraocular Pressure in Mice. Biomedicines. 2023; 11(1):60. https://doi.org/10.3390/biomedicines11010060
Chicago/Turabian StylePavlenko, Tatiana A., Andrei Y. Roman, Olga A. Lytkina, Nadezhda E. Pukaeva, Martha W. Everett, Iuliia S. Sukhanova, Vladislav O. Soldatov, Nina G. Davidova, Natalia B. Chesnokova, Ruslan K. Ovchinnikov, and et al. 2023. "Gamma-Synuclein Dysfunction Causes Autoantibody Formation in Glaucoma Patients and Dysregulation of Intraocular Pressure in Mice" Biomedicines 11, no. 1: 60. https://doi.org/10.3390/biomedicines11010060
APA StylePavlenko, T. A., Roman, A. Y., Lytkina, O. A., Pukaeva, N. E., Everett, M. W., Sukhanova, I. S., Soldatov, V. O., Davidova, N. G., Chesnokova, N. B., Ovchinnikov, R. K., & Kukharsky, M. S. (2023). Gamma-Synuclein Dysfunction Causes Autoantibody Formation in Glaucoma Patients and Dysregulation of Intraocular Pressure in Mice. Biomedicines, 11(1), 60. https://doi.org/10.3390/biomedicines11010060