

Targeting Features of Curaxin CBL0137 on Hematological Malignancies In Vitro and In Vivo
 ,
,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. Cytotoxicity Analysis
2.3. Cell Cycle Analysis Using Flow Cytometer
2.4. Annexin-FITC/Propidium Iodide Double Staining
2.5. RT2 Profiler PCR Arrays
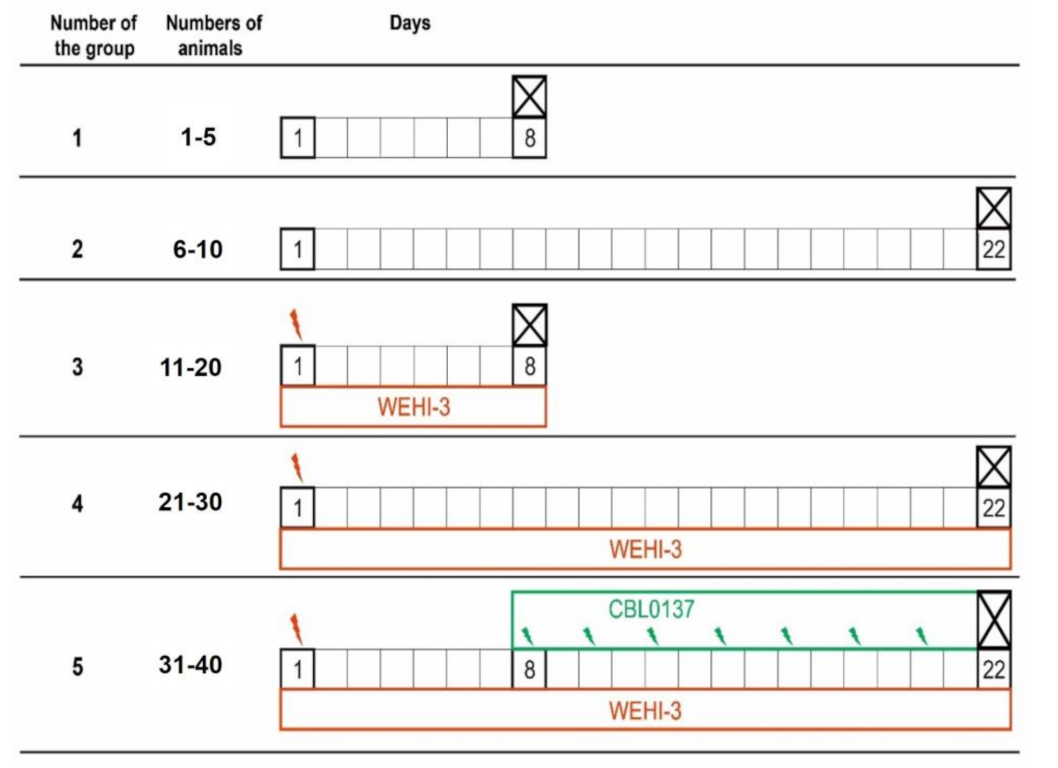
2.6. Experiments In Vivo
2.7. Data Processing
3. Results
3.1. Cytotoxic Effect of CBL0137
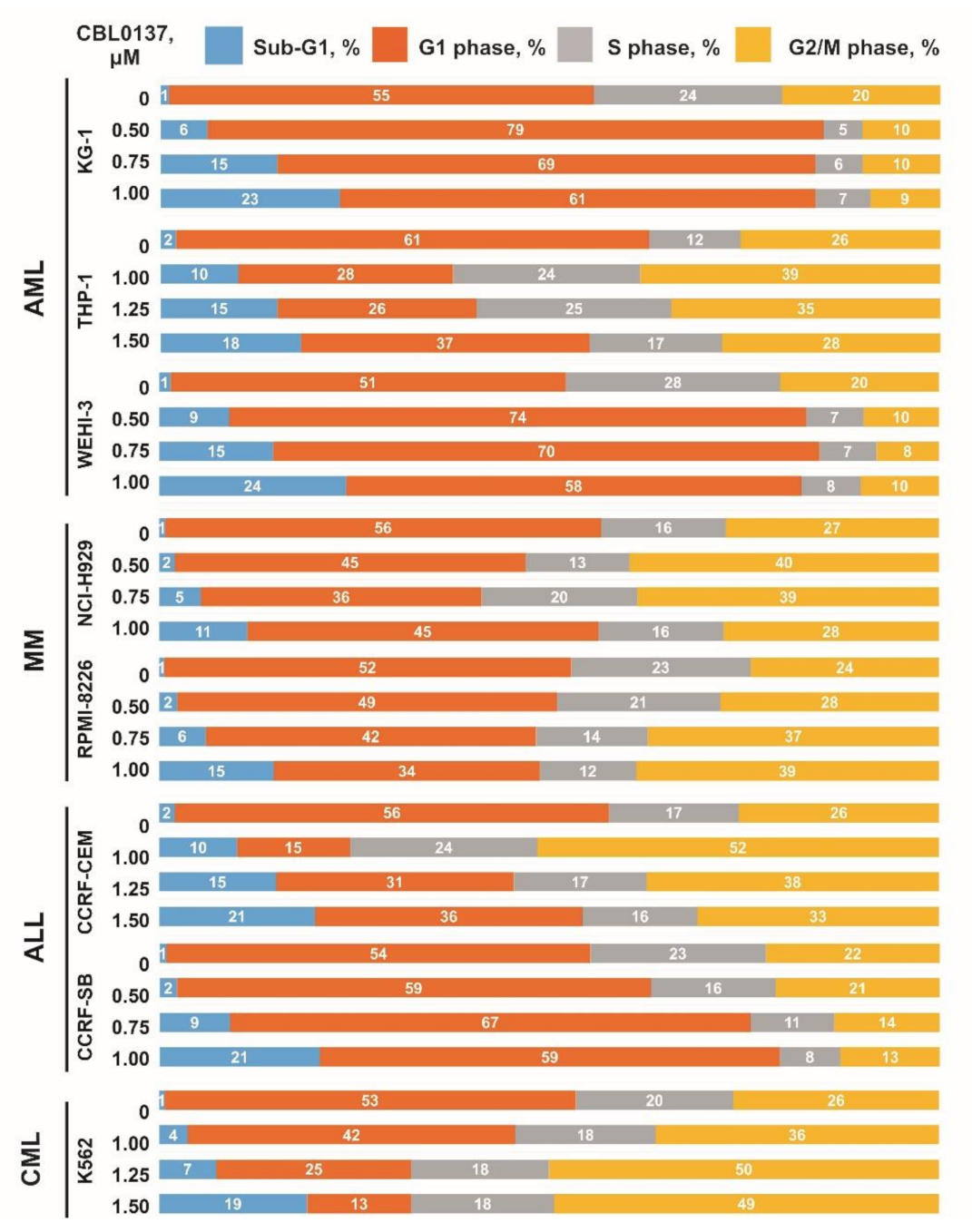
3.2. Influence CBL0137 on the Cell Cycle Distribution
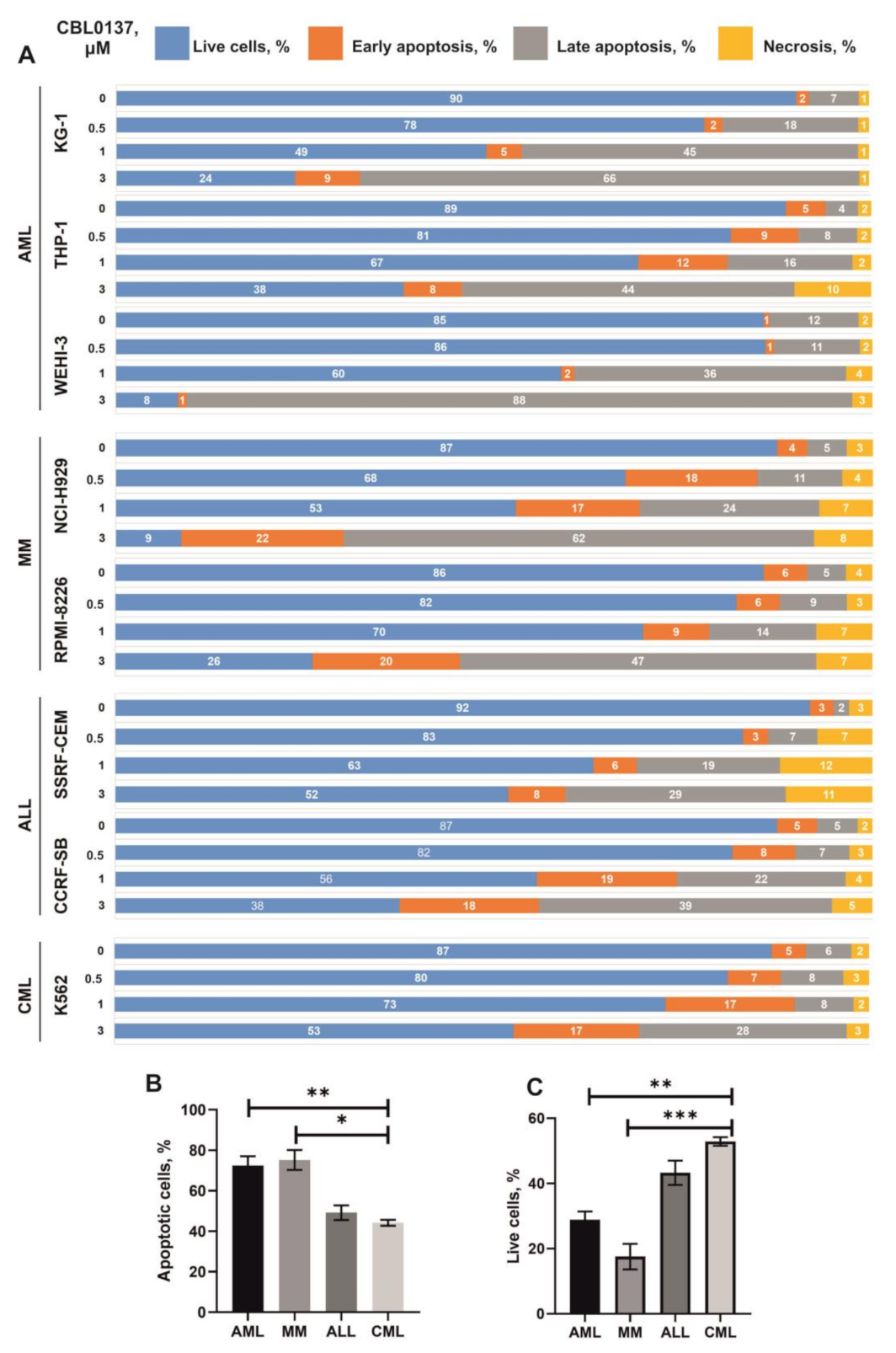
3.3. Influence CBL0137 on the Apoptosis Activation
3.4. Changes in the Activity of Signaling Pathways
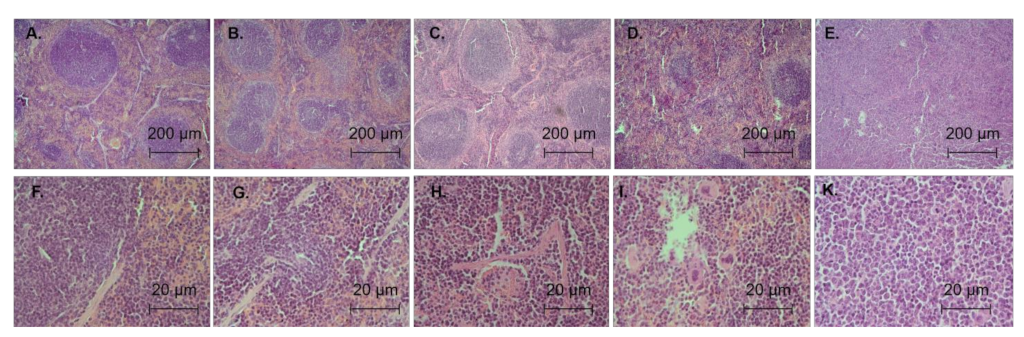
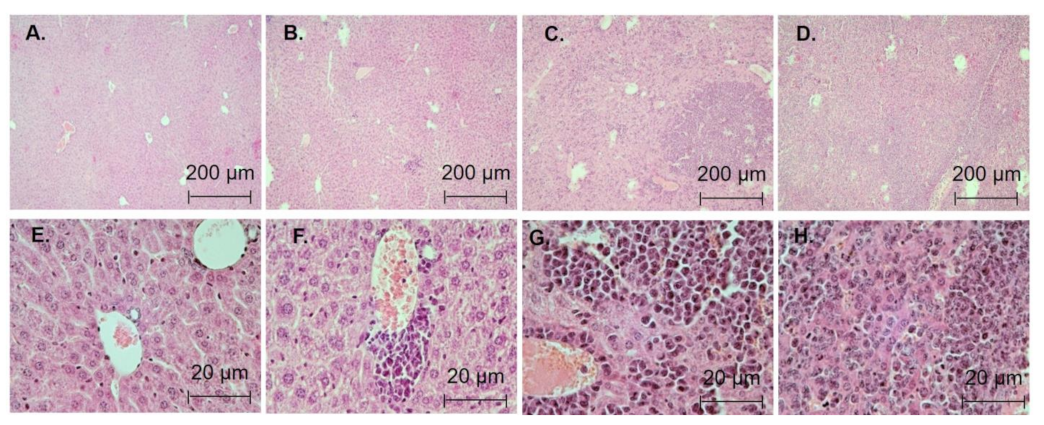
3.5. Antileukemic Activity In Vivo
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Burkhart, C.; Fleyshman, D.; Kohrn, R.; Commane, M.; Garrigan, J.; Kurbatov, V.; Toshkov, I.; Ramachandran, R.; Martello, L.; Gurova, K.V. Curaxin CBL0137 Eradicates Drug Resistant Cancer Stem Cells and Potentiates Efficacy of Gemcitabine in Preclinical Models of Pancreatic Cancer. Oncotarget 2014, 5, 11038–11053. [Google Scholar] [CrossRef] [PubMed]
- Gasparian, A.V.; Burkhart, C.A.; Purmal, A.A.; Brodsky, L.; Pal, M.; Saranadasa, M.; Bosykh, D.A.; Commane, M.; Guryanova, O.A.; Pal, S.; et al. Curaxins: Anticancer Compounds That Simultaneously Suppress NF-ΚB and Activate P53 by Targeting FACT. Sci. Transl. Med. 2011, 3, 95ra74. [Google Scholar] [CrossRef] [PubMed]
- Safina, A.; Cheney, P.; Pal, M.; Brodsky, L.; Ivanov, A.; Kirsanov, K.; Lesovaya, E.; Naberezhnov, D.; Nesher, E.; Koman, I.; et al. FACT Is a Sensor of DNA Torsional Stress in Eukaryotic Cells. Nucleic Acids Res. 2017, 45, 1925–1945. [Google Scholar] [CrossRef] [PubMed]
- Leonova, K.; Safina, A.; Nesher, E.; Sandlesh, P.; Pratt, R.; Burkhart, C.; Lipchick, B.; Gitlin, I.; Frangou, C.; Koman, I.; et al. TRAIN (Transcription of Repeats Activates INterferon) in Response to Chromatin Destabilization Induced by Small Molecules in Mammalian Cells. Elife 2018, 7, e30842. [Google Scholar] [CrossRef] [PubMed]
- Sergeev, A.; Vorobyov, A.; Yakubovskaya, M.; Kirsanova, O.; Gromova, E. Novel Anticancer Drug Curaxin CBL0137 Impairs DNA Methylation by Eukaryotic DNA Methyltransferase Dnmt3a. Bioorg. Med. Chem. Lett. 2020, 30, 127296. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.; Liu, C.; Liu, Y.; Luo, A.; Chen, J.; Lei, Z.; Kong, J.; Xiao, X.; Zhang, S.; Wang, Y.Z.; et al. Curaxin-Induced DNA Topology Alterations Trigger the Distinct Binding Response of CTCF and FACT at the Single-Molecule Level. Biochemistry 2021, 60, 494–499. [Google Scholar] [CrossRef]
- Kantidze, O.L.; Luzhin, A.V.; Nizovtseva, E.V.; Safina, A.; Valieva, M.E.; Golov, A.K.; Velichko, A.K.; Lyubitelev, A.V.; Feofanov, A.V.; Gurova, K.V.; et al. The Anti-Cancer Drugs Curaxins Target Spatial Genome Organization. Nat. Commun. 2019, 10, 1441. [Google Scholar] [CrossRef]
- De, S.; Lindner, D.J.; Coleman, C.J.; Wildey, G.; Dowlati, A.; Stark, G.R. The FACT Inhibitor CBL0137 Synergizes with Cisplatin in Small-Cell Lung Cancer by Increasing NOTCH1 Expression and Targeting Tumor-Initiating Cells. Cancer Res. 2018, 78, 2396–2406. [Google Scholar] [CrossRef] [PubMed]
- Fleyshman, D.; Prendergast, L.; Safina, A.; Paszkiewicz, G.; Commane, M.; Morgan, K.; Attwood, K.; Gurova, K. Level of FACT Defines the Transcriptional Landscape and Aggressive Phenotype of Breast Cancer Cells. Oncotarget 2017, 8, 20525–20542. [Google Scholar] [CrossRef]
- Kirsanov, K.; Fetisov, T.; Lesovaya, E.A.; Maksimova, V.; Trukhanova, L.; Antoshina, E.; Gor’kova, T.; Morozova, O.; Safina, A.; Fleyshman, D.; et al. Prevention of Colorectal Carcinogenesis by DNA-Binding Small-Molecule Curaxin CBL0137 Involves Suppression of Wnt Signaling. Cancer Prev. Res. 2020, 13, 53–64. [Google Scholar] [CrossRef]
- Lock, R.; Carol, H.; Maris, J.M.; Kolb, E.A.; Gorlick, R.; Reynolds, C.P.; Kang, M.H.; Keir, S.T.; Wu, J.; Purmal, A.; et al. Initial Testing (Stage 1) of the Curaxin CBL0137 by the Pediatric Preclinical Testing Program. Pediatr. Blood Cancer 2017, 64, e26263. [Google Scholar] [CrossRef] [PubMed]
- Somers, K.; Kosciolek, A.; Bongers, A.; El-Ayoubi, A.; Karsa, M.; Mayoh, C.; Wadham, C.; Middlemiss, S.; Neznanov, N.; Kees, U.R.; et al. Potent Antileukemic Activity of Curaxin CBL0137 against MLL-Rearranged Leukemia. Int. J. cancer 2020, 146, 1902–1916. [Google Scholar] [CrossRef]
- Verma, S.; Dhanda, H.; Singh, A.; Rishi, B.; Tanwar, P.; Chaudhry, S.; Siraj, F.; Misra, A. Systematic review of epigenetic targets in acute myeloid leukemia. Am. J. Blood Res. 2021, 11, 458–471. [Google Scholar] [PubMed]
- Salim, L.Z.; Othman, R.; Abdulla, M.A.; Al-Jashamy, K.; Ali, H.M.; Hassandarvish, P.; Dehghan, F.; Ibrahim, M.Y.; Omer, F.A.; Mohan, S. Thymoquinone Inhibits Murine Leukemia WEHI-3 Cells in Vivo and in Vitro. PLoS ONE 2014, 9, e115340. [Google Scholar] [CrossRef]
- Li, Y.; Jin, K.; Van Pelt, G.W.; Van Dam, H.; Yu, X.; Mesker, W.E.; Ten Dijke, P.; Zhou, F.; Zhang, L. C-Myb Enhances Breast Cancer Invasion and Metastasis through the Wnt/β-Catenin/Axin2 Pathway. Cancer Res. 2016, 76, 3364–3375. [Google Scholar] [CrossRef]
- Jiang, Y.; Prunier, C.; Howe, P.H. The Inhibitory Effects of Disabled-2 (Dab2) on Wnt Signaling Are Mediated through Axin. Oncogene 2008, 27, 1865–1875. [Google Scholar] [CrossRef]
- Thorfve, A.; Bergstrand, A.; Ekström, K.; Lindahl, A.; Thomsen, P.; Larsson, A.; Tengvall, P. Gene Expression Profiling of Peri-Implant Healing of PLGA-Li+ Implants Suggests an Activated Wnt Signaling Pathway in Vivo. PLoS ONE 2014, 9, e102597. [Google Scholar] [CrossRef]
- Zhang, M.; Du, H.; Wang, L.; Yue, Y.; Zhang, P.; Huang, Z.; Lv, W.; Ma, J.; Shao, Q.; Ma, M.; et al. Thymoquinone Suppresses Invasion and Metastasis in Bladder Cancer Cells by Reversing EMT through the Wnt/β-Catenin Signaling Pathway. Chem. Biol. Interact. 2020, 320, 109022. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.H.; Lafita-Navarro, M.C.; Zacharias, L.; Borenstein-Auerbach, N.; Kim, M.; Barnes, S.; Kim, J.; Shay, J.; Deberardinis, R.J.; Conacci-Sorrell, M. Induction of LEF1 by MYC Activates the WNT Pathway and Maintains Cell Proliferation. Cell Commun. Signal. 2019, 17, 129. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Song, Y.; Ye, M.; Hu, X.; Wang, Z.P.; Zhu, X. The Emerging Role of WISP Proteins in Tumorigenesis and Cancer Therapy. J. Transl. Med. 2019, 17, 28. [Google Scholar] [CrossRef]
- Scholtysek, C.; Katzenbeisser, J.; Fu, H.; Uderhardt, S.; Ipseiz, N.; Stoll, C.; Zaiss, M.M.; Stock, M.; Donhauser, L.; Böhm, C.; et al. PPARβ/δ Governs Wnt Signaling and Bone Turnover. Nat. Med. 2013, 19, 608–613. [Google Scholar] [CrossRef] [PubMed]
- Luiken, S.; Fraas, A.; Bieg, M.; Sugiyanto, R.; Goeppert, B.; Singer, S.; Ploeger, C.; Warsow, G.; Marquardt, J.U.; Sticht, C.; et al. NOTCH Target Gene HES5 Mediates Oncogenic and Tumor Suppressive Functions in Hepatocarcinogenesis. Oncogene 2020, 39, 3128–3144. [Google Scholar] [CrossRef] [PubMed]
- Braccioli, L.; Vervoort, S.J.; Puma, G.; Nijboer, C.H.; Coffer, P.J. SOX4 Inhibits Oligodendrocyte Differentiation of Embryonic Neural Stem Cells in Vitro by Inducing Hes5 Expression. Stem Cell Res. 2018, 33, 110–119. [Google Scholar] [CrossRef]
- Scroyen, I.; Bauters, D.; Vranckx, C.; Lijnen, H.R. The Anti-Adipogenic Potential of COUP-TFII Is Mediated by Downregulation of the Notch Target Gene Hey1. PLoS ONE 2015, 10, e0145608. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Zeng, Z.; Xing, J.; Zhang, A.; Jiang, W.; Wang, W.; Sun, H.; Ni, L. Hey1 Functions as a Positive Regulator of Odontogenic Differentiation in Odontoblast-lineage Cells. Int. J. Mol. Med. 2018, 41, 331–339. [Google Scholar] [CrossRef]
- Miao, L.; Li, J.; Li, J.; Tian, X.; Lu, Y.; Hu, S.; Shieh, D.; Kanai, R.; Zhou, B.Y.; Zhou, B.; et al. Notch Signaling Regulates Hey2 Expression in a Spatiotemporal Dependent Manner during Cardiac Morphogenesis and Trabecular Specification. Sci. Rep. 2018, 8, 2678. [Google Scholar] [CrossRef]
- Anderson, D.J.; Kaplan, D.I.; Bell, K.M.; Koutsis, K.; Haynes, J.M.; Mills, R.J.; Phelan, D.G.; Qian, E.L.; Leitoguinho, A.R.; Arasaratnam, D.; et al. NKX2-5 Regulates Human Cardiomyogenesis via a HEY2 Dependent Transcriptional Network. Nat. Commun. 2018, 9, 1373. [Google Scholar] [CrossRef]
- Weber, S.; Koschade, S.E.; Hoffmann, C.M.; Dubash, T.D.; Giessler, K.M.; Dieter, S.M.; Herbst, F.; Glimm, H.; Ball, C.R. The Notch Target Gene HEYL Modulates Metastasis Forming Capacity of Colorectal Cancer Patient-Derived Spheroid Cells in Vivo. BMC Cancer 2019, 19, 1181. [Google Scholar] [CrossRef]
- Jalali, A.; Bassuk, A.G.; Kan, L.; Israsena, N.; Mukhopadhyay, A.; McGuire, T.; Kessler, J.A. HeyL Promotes Neuronal Differentiation of Neural Progenitor Cells. J. Neurosci. Res. 2011, 89, 299–309. [Google Scholar] [CrossRef]
- Wang, H.-C.; Perry, S.S.; Sun, X.-H. Id1 Attenuates Notch Signaling and Impairs T-Cell Commitment by Elevating Deltex1 Expression. Mol. Cell. Biol. 2009, 29, 4640–4652. [Google Scholar] [CrossRef]
- Chadwick, N.; Zeef, L.; Portillo, V.; Fennessy, C.; Warrander, F.; Hoyle, S.; Buckle, A.M. Identification of Novel Notch Target Genes in T Cell Leukaemia. Mol. Cancer 2009, 8, 35. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhu, Y.; Li, F.; Xie, Y. GATA1-Regulated JAG1 Promotes Ovarian Cancer Progression by Activating Notch Signal Pathway. Protoplasma 2020, 257, 901–910. [Google Scholar] [CrossRef] [PubMed]
- Okubo, Y.; Sugawara, T.; Abe-Koduka, N.; Kanno, J.; Kimura, A.; Saga, Y. Lfng Regulates the Synchronized Oscillation of the Mouse Segmentation Clock via Trans-Repression of Notch Signalling. Nat. Commun. 2012, 3, 1141. [Google Scholar] [CrossRef] [PubMed]
- Venkatesh, V.; Nataraj, R.; Thangaraj, G.S.; Karthikeyan, M.; Gnanasekaran, A.; Kaginelli, S.B.; Kuppanna, G.; Kallappa, C.G.; Basalingappa, K.M. Targeting Notch Signalling Pathway of Cancer Stem Cells. Stem Cell Investig. 2018, 5, 5. [Google Scholar] [CrossRef] [PubMed]
- Bigelow, R.L.H.; Chari, N.S.; Undén, A.B.; Spurgers, K.B.; Lee, S.; Roop, D.R.; Toftgård, R.; McDonnell, T.J. Transcriptional Regulation of Bcl-2 Mediated by the Sonic Hedgehog Signaling Pathway through Gli-1. J. Biol. Chem. 2004, 279, 1197–1205. [Google Scholar] [CrossRef]
- Banerjee, S.; Corless, C.L.; Miettinen, M.M.; Noh, S.; Ustoy, R.; Davis, J.L.; Tang, C.M.; Yebra, M.; Burgoyne, A.M.; Sicklick, J.K. Loss of the PTCH1 Tumor Suppressor Defines a New Subset of Plexiform Fibromyxoma. J. Transl. Med. 2019, 17, 246–248. [Google Scholar] [CrossRef]
- Shahi, M.H.; Afzal, M.; Sinha, S.; Eberhart, C.G.; Rey, J.A.; Fan, X.; Castresana, J.S. Regulation of Sonic Hedgehog-GLI1 Downstream Target Genes PTCH1, Cyclin D2, Plakoglobin, PAX6 and NKX2.2 and Their Epigenetic Status in Medulloblastoma and Astrocytoma. BMC Cancer 2010, 10, 614. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, Z.; Zhao, X.; Yu, X.; Hu, Y.; Geronimo, B.; Fromm, S.H.; Chen, Y.P. A New Function of BMP4: Dual Role for BMP4 in Regulation of Sonic Hedgehog Expression in the Mouse Tooth Germ. Development 2000, 127, 1431–1443. [Google Scholar] [CrossRef]
- Astorga, J.; Carlsson, P. Hedgehog Induction of Murine Vasculogenesis Is Mediated by Foxf1 and Bmp4. Development 2007, 134, 3753–3761. [Google Scholar] [CrossRef]
- Li, X.; Deng, W.; Lobo-Ruppert, S.M.; Ruppert, J.M. Gli1 Acts through Snail and E-Cadherin to Promote Nuclear Signaling by Beta-Catenin. Oncogene 2007, 26, 4489–4498. [Google Scholar] [CrossRef]
- Tomasic, N.L.; Piterkova, L.; Huff, C.; Bilic, E.; Yoon, D.; Miasnikova, G.Y.; Sergueeva, A.I.; Niu, X.; Nekhai, S.; Gordeuk, V.; et al. The Phenotype of Polycythemia Due to Croatian Homozygous VHL (571C>G:H191D) Mutation Is Different from That of Chuvash Polycythemia (VHL 598C>T:R200W). Haematologica 2013, 98, 560–567. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xu, Y.; Ma, J.; Pang, X.; Dong, M. Adrenomedullin Promotes Angiogenesis in Epithelial Ovarian Cancer through Upregulating Hypoxia-Inducible Factor-1α and Vascular Endothelial Growth Factor. Sci. Rep. 2017, 7, 40524. [Google Scholar] [CrossRef] [PubMed]
- van den Beucken, T.; Koritzinsky, M.; Niessen, H.; Dubois, L.; Savelkouls, K.; Mujcic, H.; Jutten, B.; Kopacek, J.; Pastorekova, S.; van der Kogel, A.J.; et al. Hypoxia-Induced Expression of Carbonic Anhydrase 9 Is Dependent on the Unfolded Protein Response. J. Biol. Chem. 2009, 284, 24204–24212. [Google Scholar] [CrossRef] [PubMed]
- Pastorekova, S.; Gillies, R.J. The Role of Carbonic Anhydrase IX in Cancer Development: Links to Hypoxia, Acidosis, and Beyond. Cancer Metastasis Rev. 2019, 38, 65–77. [Google Scholar] [CrossRef]
- Cheng, L.; Yu, H.; Yan, N.; Lai, K.; Xiang, M. Hypoxia-Inducible Factor-1α Target Genes Contribute to Retinal Neuroprotection. Front. Cell. Neurosci. 2017, 11, 20. [Google Scholar] [CrossRef] [PubMed]
- Kimáková, P.; Solár, P.; Solárová, Z.; Komel, R.; Debeljak, N. Erythropoietin and Its Angiogenic Activity. Int. J. Mol. Sci. 2017, 18, 1519. [Google Scholar] [CrossRef]
- Dunn, L.L.; Kong, S.M.Y.; Tumanov, S.; Chen, W.; Cantley, J.; Ayer, A.; Maghzal, G.J.; Midwinter, R.G.; Chan, K.H.; Ng, M.K.C.; et al. Hmox1 (Heme Oxygenase-1) Protects Against Ischemia-Mediated Injury via Stabilization of HIF-1α (Hypoxia-Inducible Factor-1α). Arterioscler. Thromb. Vasc. Biol. 2021, 41, 317–330. [Google Scholar] [CrossRef]
- Benita, Y.; Kikuchi, H.; Smith, A.D.; Zhang, M.Q.; Chung, D.C.; Xavier, R.J. An Integrative Genomics Approach Identifies Hypoxia Inducible Factor-1 (HIF-1)-Target Genes That Form the Core Response to Hypoxia. Nucleic Acids Res. 2009, 37, 4587–4602. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Zhang, F.; He, D.; Ji, X.; Gao, J.; Liu, W.; Wang, Y.; Liu, Q.; Xin, T. Immune-Related Gene SERPINE1 Is a Novel Biomarker for Diffuse Lower-Grade Gliomas via Large-Scale Analysis. Front. Oncol. 2021, 11, 1800. [Google Scholar] [CrossRef]
- Dengler, V.L.; Galbraith, M.D.; Espinosa, J.M. Transcriptional Regulation by Hypoxia Inducible Factors. Crit. Rev. Biochem. Mol. Biol. 2014, 49, 1–15. [Google Scholar] [CrossRef]
- Yan, S.; Wang, Y.; Chen, M.; Li, G.; Fan, J. Deregulated SLC2A1 Promotes Tumor Cell Proliferation and Metastasis in Gastric Cancer. Int. J. Mol. Sci. 2015, 16, 16144–16157. [Google Scholar] [CrossRef] [PubMed]
- Manalo, D.J.; Rowan, A.; Lavoie, T.; Natarajan, L.; Kelly, B.D.; Ye, S.Q.; Garcia, J.G.N.; Semenza, G.L. Transcriptional Regulation of Vascular Endothelial Cell Responses to Hypoxia by HIF-1. Blood 2005, 105, 659–669. [Google Scholar] [CrossRef]
- McFee, R.M.; Rozell, T.G.; Cupp, A.S. The Balance of Proangiogenic and Antiangiogenic VEGFA Isoforms Regulate Follicle Development. Cell Tissue Res. 2012, 349, 635–647. [Google Scholar] [CrossRef]
- Langhammer, S.; Najjar, M.; Hess-Stumpp, H.; Thierauch, K.H. LDH-A Influences Hypoxia-Inducible Factor 1α (HIF1 α) and Is Critical for Growth of HT29 Colon Carcinoma Cells in Vivo. Target. Oncol. 2011, 6, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Rubattu, S.; Mennuni, S.; Testa, M.; Mennuni, M.; Pierelli, G.; Pagliaro, B.; Gabriele, E.; Coluccia, R.; Autore, C.; Volpe, M. Pathogenesis of Chronic Cardiorenal Syndrome: Is There a Role for Oxidative Stress? Int. J. Mol. Sci. 2013, 14, 23011–23032. [Google Scholar] [CrossRef]
- Wu, Z.; Harne, R.; Chintoan-Uta, C.; Hu, T.J.; Wallace, R.; MacCallum, A.; Stevens, M.P.; Kaiser, P.; Balic, A.; Hume, D.A. Regulation and Function of Macrophage Colony-Stimulating Factor (CSF1) in the Chicken Immune System. Dev. Comp. Immunol. 2020, 105, 103586. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Shi, L.Z.; Zhao, H.; Chen, J.; Xiong, L.; He, Q.; Chen, T.; Roszik, J.; Bernatchez, C.; Woodman, S.E.; et al. Loss of IFN-γ Pathway Genes in Tumor Cells as a Mechanism of Resistance to Anti-CTLA-4 Therapy. Cell 2016, 167, 397–404.e9. [Google Scholar] [CrossRef]
- Khaidakov, M.; Mitra, S.; Kang, B.Y.; Wang, X.; Kadlubar, S.; Novelli, G.; Raj, V.; Winters, M.; Carter, W.C.; Mehta, J.L. Oxidized LDL Receptor 1 (OLR1) as a Possible Link between Obesity, Dyslipidemia and Cancer. PLoS ONE 2011, 6, e20277. [Google Scholar] [CrossRef]
- Vogler, M. BCL2A1: The Underdog in the BCL2 Family. Cell Death Differ. 2012, 19, 67–74. [Google Scholar] [CrossRef]
- Mansouri, L.; Papakonstantinou, N.; Ntoufa, S.; Stamatopoulos, K.; Rosenquist, R. NF-ΚB Activation in Chronic Lymphocytic Leukemia: A Point of Convergence of External Triggers and Intrinsic Lesions. Semin. Cancer Biol. 2016, 39, 40–48. [Google Scholar] [CrossRef]
- Zhang, S.; Yang, Y.; Weng, W.; Guo, B.; Cai, G.; Ma, Y.; Cai, S. Fusobacterium Nucleatum Promotes Chemoresistance to 5-Fluorouracil by Upregulation of BIRC3 Expression in Colorectal Cancer. J. Exp. Clin. Cancer Res. 2019, 38, 14. [Google Scholar] [CrossRef] [PubMed]
- Tian, B.; Nowak, D.E.; Brasier, A.R. A TNF-Induced Gene Expression Program under Oscillatory NF-KappaB Control. BMC Genom. 2005, 6, 137. [Google Scholar] [CrossRef] [PubMed]
- Shakhov, A.N.; Kuprash, D.V.; Azizov, M.M.; Jongeneel, C.V.; Nedospasov, S.A. Structural Analysis of the Rabbit TNF Locus, Containing the Genes Encoding TNF-Beta (Lymphotoxin) and TNF-Alpha (Tumor Necrosis Factor). Gene 1990, 95, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Burke, S.J.; Lu, D.; Sparer, T.E.; Masi, T.; Goff, M.R.; Karlstad, M.D.; Collier, J.J. NF-ΚB and STAT1 Control CXCL1 and CXCL2 Gene Transcription. Am. J. Physiol. Endocrinol. Metab. 2014, 306, E131–E149. [Google Scholar] [CrossRef]
- Wickremasinghe, M.I.; Thomas, L.H.; O’Kane, C.M.; Uddin, J.; Friedland, J.S. Transcriptional Mechanisms Regulating Alveolar Epithelial Cell-Specific CCL5 Secretion in Pulmonary Tuberculosis. J. Biol. Chem. 2004, 279, 27199–27210. [Google Scholar] [CrossRef]
- Palchevskiy, V.; Hashemi, N.; Weigt, S.S.; Xue, Y.Y.; Derhovanessian, A.; Keane, M.P.; Strieter, R.M.; Fishbein, M.C.; Deng, J.C.; Lynch, J.P.; et al. Immune Response CC Chemokines CCL2 and CCL5 Are Associated with Pulmonary Sarcoidosis. Fibrogenesis Tissue Repair 2011, 4, 10. [Google Scholar] [CrossRef]
- Gonzalez-González, A.; Muñoz-Muela, E.; Marchal, J.A.; Cara, F.E.; Molina, M.P.; Cruz-Lozano, M.; Jiménez, G.; Verma, A.; Ramírez, A.; Qian, W.; et al. Activating Transcription Factor 4 Modulates TGFβ-Induced Aggressiveness in Triple-Negative Breast Cancer via SMAD2/3/4 and MTORC2 Signaling. Clin. Cancer Res. 2018, 24, 5697–5709. [Google Scholar] [CrossRef]
- Li, D.; Li, X.; Wang, A.; Meisgen, F.; Pivarcsi, A.; Sonkoly, E.; Ståhle, M.; Landén, N.X. MicroRNA-31 Promotes Skin Wound Healing by Enhancing Keratinocyte Proliferation and Migration. J. Investig. Dermatol. 2015, 135, 1676–1685. [Google Scholar] [CrossRef]
- Feng, X.H.; Liang, Y.Y.; Liang, M.; Zhai, W.; Lin, X. Direct Interaction of C-Myc with Smad2 and Smad3 to Inhibit TGF-Beta-Mediated Induction of the CDK Inhibitor P15(Ink4B). Mol. Cell 2002, 9, 133–143. [Google Scholar] [CrossRef]
- Lux, A.; Salway, F.; Dressman, H.K.; Kröner-Lux, G.; Hafner, M.; Day, P.J.R.; Marchuk, D.A.; Garland, J. ALK1 Signalling Analysis Identifies Angiogenesis Related Genes and Reveals Disparity between TGF-Beta and Constitutively Active Receptor Induced Gene Expression. BMC Cardiovasc. Disord. 2006, 6, 13. [Google Scholar] [CrossRef]
- Liu, B.; Sun, X.; Suyeoka, G.; Garcia, J.G.N.; Leiderman, Y.I. TGFβ Signaling Induces Expression of Gadd45b in Retinal Ganglion Cells. Investig. Ophthalmol. Vis. Sci. 2013, 54, 1061–1069. [Google Scholar] [CrossRef] [PubMed]
- Marinello, P.C.; Panis, C.; Silva, T.N.X.; Binato, R.; Abdelhay, E.; Rodrigues, J.A.; Mencalha, A.L.; Lopes, N.M.D.; Borges, F.H.; Luiz, R.C.; et al. Oxidative Stress and TGF-Β1 Induction by Metformin in MCF-7 and MDA-MB-231 Human Breast Cancer Cells Are Accompanied with the Downregulation of Genes Related to Cell Proliferation, Invasion and Metastasis. Pathol. Res. Pract. 2020, 216, 153135. [Google Scholar] [CrossRef] [PubMed]
- Lähde, M.; Heino, S.; Högström, J.; Kaijalainen, S.; Anisimov, A.; Flanagan, D.; Kallio, P.; Leppänen, V.M.; Ristimäki, A.; Ritvos, O.; et al. Expression of R-Spondin 1 in Apc Min/+ Mice Suppresses Growth of Intestinal Adenomas by Altering Wnt and Transforming Growth Factor Beta Signaling. Gastroenterology 2021, 160, 245–259. [Google Scholar] [CrossRef]
- Minarikova, P.; Benesova, L.; Halkova, T.; Belsanova, B.; Tuckova, I.; Belina, F.; Dusek, L.; Zavoral, M.; Minarik, M. Prognostic Importance of Cell Cycle Regulators Cyclin D1 (CCND1) and Cyclin-Dependent Kinase Inhibitor 1B (CDKN1B/P27) in Sporadic Gastric Cancers. Gastroenterol. Res. Pract. 2016, 2016, 9408190. [Google Scholar] [CrossRef]
- Budi, E.H.; Hoffman, S.; Gao, S.; Zhang, Y.E.; Derynck, R. Integration of TGF-β-Induced Smad Signaling in the Insulin-Induced Transcriptional Response in Endothelial Cells. Sci. Rep. 2019, 9, 16992. [Google Scholar] [CrossRef] [PubMed]
- Kuribayashi, K.; Krigsfeld, G.; Wang, W.; Xu, J.; Mayes, P.A.; Dicker, D.T.; Gen, S.W.; El-Deiry, W.S. TNFSF10 (TRAIL), a P53 Target Gene That Mediates P53-Dependent Cell Death. Cancer Biol. Ther. 2008, 7, 2034–2038. [Google Scholar] [CrossRef] [PubMed]
- Lim, D.; Chai, H.H.; Lee, S.H.; Cho, Y.M.; Choi, J.W.; Kim, N.K. Gene Expression Patterns Associated with Peroxisome Proliferator-Activated Receptor (PPAR) Signaling in the Longissimus Dorsi of Hanwoo (Korean Cattle). Asian-Australas. J. Anim. Sci. 2015, 28, 1075–1083. [Google Scholar] [CrossRef]
- Chui, P.C.; Guan, H.P.; Lehrke, M.; Lazar, M.A. PPARgamma Regulates Adipocyte Cholesterol Metabolism via Oxidized LDL Receptor 1. J. Clin. Investig. 2005, 115, 2244–2256. [Google Scholar] [CrossRef]
- Anderson, C.M.; Stahl, A. SLC27 Fatty Acid Transport Proteins. Mol. Aspects Med. 2013, 34, 516–528. [Google Scholar] [CrossRef]
- Carow, B.; Rottenberg, M.E. SOCS3, a Major Regulator of Infection and Inflammation. Front. Immunol. 2014, 5, 58. [Google Scholar] [CrossRef]
- Cao, A.; Li, H.; Zhou, Y.; Wu, M.; Liu, J. Long Chain Acyl-CoA Synthetase-3 Is a Molecular Target for Peroxisome Proliferator-Activated Receptor Delta in HepG2 Hepatoma Cells. J. Biol. Chem. 2010, 285, 16664–16674. [Google Scholar] [CrossRef] [PubMed]
- Kan, C.F.K.; Singh, A.B.; Dong, B.; Shende, V.R.; Liu, J. PPARδ Activation Induces Hepatic Long-Chain Acyl-CoA Synthetase 4 Expression in Vivo and in Vitro. Biochim. Biophys. Acta 2015, 1851, 577–587. [Google Scholar] [CrossRef] [PubMed]
- Belle, J.I.; Petrov, J.C.; Langlais, D.; Robert, F.; Cencic, R.; Shen, S.; Pelletier, J.; Gros, P.; Nijnik, A. Repression of P53-Target Gene Bbc3/PUMA by MYSM1 Is Essential for the Survival of Hematopoietic Multipotent Progenitors and Contributes to Stem Cell Maintenance. Cell Death Differ. 2016, 23, 759–775. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Cheng, B.; Miao, L.; Me, Y.; Wu, M. Mutant P53-R273H Gains New Function in Sustained Activation of EGFR Signaling via Suppressing MiR-27a Expression. Cell Death Dis. 2013, 4, e574. [Google Scholar] [CrossRef]
- Liu, T.; Laurell, C.; Selivanova, G.; Lundeberg, J.; Nilsson, P.; Wiman, K.G. Hypoxia Induces P53-Dependent Transactivation and Fas/CD95-Dependent Apoptosis. Cell Death Differ. 2007, 14, 411–421. [Google Scholar] [CrossRef]
- Tsui, K.H.; Chiang, K.C.; Lin, Y.H.; Chang, K.S.; Feng, T.H.; Juang, H.H. BTG2 Is a Tumor Suppressor Gene Upregulated by P53 and PTEN in Human Bladder Carcinoma Cells. Cancer Med. 2018, 7, 184–195. [Google Scholar] [CrossRef]
- Xu, J.; Morris, G.F. P53-Mediated Regulation of Proliferating Cell Nuclear Antigen Expression in Cells Exposed to Ionizing Radiation. Mol. Cell. Biol. 1999, 19, 12–20. [Google Scholar] [CrossRef]
- De Feudis, P.; Vignati, S.; Rossi, C.; Mincioni, T.; Giavazzi, R.; D’Incalci, M.; Broggini, M. Driving P53 Response to Bax Activation Greatly Enhances Sensitivity to Taxol by Inducing Massive Apoptosis. Neoplasia 2000, 2, 202–207. [Google Scholar] [CrossRef]
- Kershaw, N.J.; Murphy, J.M.; Liau, N.P.D.; Varghese, L.N.; Laktyushin, A.; Whitlock, E.L.; Lucet, I.S.; Nicola, N.A.; Babon, J.J. SOCS3 Binds Specific Receptor-JAK Complexes to Control Cytokine Signaling by Direct Kinase Inhibition. Nat. Struct. Mol. Biol. 2013, 20, 469–476. [Google Scholar] [CrossRef]
- Ibrahim, H.; Askar, B.; Hulme, S.; Neilson, P.; Barrow, P.; Foster, N. Differential Immune Phenotypes in Human Monocytes Induced by Non-Host-Adapted Salmonella Enterica Serovar Choleraesuis and Host-Adapted S. Typhimurium. Infect. Immun. 2018, 86, e00509-18. [Google Scholar] [CrossRef]
- Liu, Y.; Liao, S.; Bennett, S.; Tang, H.; Song, D.; Wood, D.; Zhan, X.; Xu, J. STAT3 and Its Targeting Inhibitors in Osteosarcoma. Cell Prolif. 2021, 54, e12974. [Google Scholar] [CrossRef] [PubMed]
- Majoros, A.; Platanitis, E.; Kernbauer-Hölzl, E.; Rosebrock, F.; Müller, M.; Decker, T. Canonical and Non-Canonical Aspects of JAK-STAT Signaling: Lessons from Interferons for Cytokine Responses. Front. Immunol. 2017, 8, 29. [Google Scholar] [CrossRef] [PubMed]
- Sanford, D.C.; DeWille, J.W. C/EBPdelta Is a Downstream Mediator of IL-6 Induced Growth Inhibition of Prostate Cancer Cells. Prostate 2005, 63, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Xiao, S.; Zhu, H. Leucine-Rich Alpha-2-Glycoprotein1 Gene Interferes with Regulation of Apoptosis in Leukemia KASUMI-1 Cells. Med. Sci. Monit. 2018, 24, 8348–8356. [Google Scholar] [CrossRef] [PubMed]
- Lüersen, K.; Stegehake, D.; Daniel, J.; Drescher, M.; Ajonina, I.; Ajonina, C.; Hertel, P.; Woltersdorf, C.; Liebau, E. The Glutathione Reductase GSR-1 Determines Stress Tolerance and Longevity in Caenorhabditis Elegans. PLoS ONE 2013, 8, e60731. [Google Scholar] [CrossRef]
- Li, H.; Liu, B.; Lian, L.; Zhou, J.; Xiang, S.; Zhai, Y.; Chen, Y.; Ma, X.; Wu, W.; Hou, L. High Dose Expression of Heme Oxigenase-1 Induces Retinal Degeneration through ER Stress-Related DDIT3. Mol. Neurodegener. 2021, 16, 16. [Google Scholar] [CrossRef]
- Leone, A.; Roca, M.S.; Ciardiello, C.; Costantini, S.; Budillon, A. Oxidative Stress Gene Expression Profile Correlates with Cancer Patient Poor Prognosis: Identification of Crucial Pathways Might Select Novel Therapeutic Approaches. Oxidative Med. Cell. Longev. 2017, 2017, 2597581. [Google Scholar] [CrossRef]
- Dinkova-Kostova, A.T.; Talalay, P. NAD(P)H:Quinone Acceptor Oxidoreductase 1 (NQO1), a Multifunctional Antioxidant Enzyme and Exceptionally Versatile Cytoprotector. Arch. Biochem. Biophys. 2010, 501, 116–123. [Google Scholar] [CrossRef]
- Warner, N.L.; Moore, M.A.; Metcalf, D. A Transplantable Myelomonocytic Leukemia in BALB-c Mice: Cytology, Karyotype, and Muramidase Content. J. Natl. Cancer Inst. 1969, 43, 963–982. [Google Scholar]
- Wang, J.; Sui, Y.; Li, Q.; Zhao, Y.; Dong, X.; Yang, J.; Liang, Z.; Han, Y.; Tang, Y.; Ma, J. Effective Inhibition of MYC-Amplified Group 3 Medulloblastoma by FACT-Targeted Curaxin Drug CBL0137. Cell Death Dis. 2020, 11, 1029. [Google Scholar] [CrossRef]
- Peng, Y.T.; Wu, W.R.; Chen, L.R.; Kuo, K.K.; Tsai, C.H.; Huang, Y.T.; Lan, Y.H.; Chang, F.R.; Wu, Y.C.; Shiue, Y.L. Upregulation of Cyclin-Dependent Kinase Inhibitors CDKN1B and CDKN1C in Hepatocellular Carcinoma-Derived Cells via Goniothalamin-Mediated Protein Stabilization and Epigenetic Modifications. Toxicol. Rep. 2015, 2, 322–332. [Google Scholar] [CrossRef] [PubMed]
- Montalto, F.I.; De Amicis, F. Cyclin D1 in Cancer: A Molecular Connection for Cell Cycle Control, Adhesion and Invasion in Tumor and Stroma. Cells 2020, 9, 2648. [Google Scholar] [CrossRef]
- Zhou, H.R.; Shen, J.Z.; Fu, H.Y.; Zhang, F. Genistein-Induced Anticancer Effects on Acute Leukemia Cells Involve the Regulation of Wnt Signaling Pathway Through H4K20me1 Rather Than DNA Demethylation. Curr. Med. Sci. 2021, 41, 869–879. [Google Scholar] [CrossRef] [PubMed]
- Dzobo, K.; Thomford, N.E.; Senthebane, D.A. Targeting the Versatile Wnt/β-Catenin Pathway in Cancer Biology and Therapeutics: From Concept to Actionable Strategy. OMICS 2019, 23, 517–538. [Google Scholar] [CrossRef] [PubMed]
- Fetisov, T.I.; Lesovaya, E.A.; Yakubovskaya, M.G.; Kirsanov, K.I.; Belitsky, G.A. Alterations in WNT Signaling in Leukemias. Biochemistry 2018, 83, 1448–1458. [Google Scholar] [CrossRef]
- Cortes, J.E.; Gutzmer, R.; Kieran, M.W.; Solomon, J.A. Hedgehog Signaling Inhibitors in Solid and Hematological Cancers. Cancer Treat. Rev. 2019, 76, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, A.; Banerjee, D.; Chandra, S.; Banerji, S.K.; Ghosh, R.; Roy, R.; Banerjee, S. Deregulation and Cross Talk among Sonic Hedgehog, Wnt, Hox and Notch Signaling in Chronic Myeloid Leukemia Progression. Leukemia 2007, 21, 949–955. [Google Scholar] [CrossRef]
- Varnat, F.; Siegl-Cachedenier, I.; Malerba, M.; Gervaz, P.; Ruiz, I.; Altaba, A. Loss of WNT-TCF Addiction and Enhancement of HH-GLI1 Signalling Define the Metastatic Transition of Human Colon Carcinomas. EMBO Mol. Med. 2010, 2, 440–457. [Google Scholar] [CrossRef]
- Aster, J.C.; Pear, W.S.; Blacklow, S.C. The Varied Roles of Notch in Cancer. Annu. Rev. Pathol. 2017, 12, 245–275. [Google Scholar] [CrossRef]
- Lim, J.S.; Ibaseta, A.; Fischer, M.M.; Cancilla, B.; O’Young, G.; Cristea, S.; Luca, V.C.; Yang, D.; Jahchan, N.S.; Hamard, C.; et al. Intratumoural Heterogeneity Generated by Notch Signalling Promotes Small-Cell Lung Cancer. Nature 2017, 545, 360–364. [Google Scholar] [CrossRef]
- Weng, A.P.; Ferrando, A.A.; Lee, W.; Morris IV, J.P.; Silverman, L.B.; Sanchez-Irizarry, C.; Blacklow, S.C.; Look, A.T.; Aster, J.C. Activating Mutations of NOTCH1 in Human T Cell Acute Lymphoblastic Leukemia. Science 2004, 306, 269–271. [Google Scholar] [CrossRef] [PubMed]
- Eischen, C.M.; Weber, J.D.; Roussel, M.F.; Sherr, C.J.; Cleveland, J.L. Disruption of the ARF-Mdm2-P53 Tumor Suppressor Pathway in Myc-Induced Lymphomagenesis. Genes Dev. 1999, 13, 2658–2669. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Tania, M.; Fu, J. Epigenetic role of thymoquinone: Impact on cellular mechanism and cancer therapeutics. Drug Discov. Today 2019, 24, 2315–2322. [Google Scholar] [CrossRef] [PubMed]
- Herold, T.; Rothenberg-Thurley, M.; Grunwald, V.V.; Janke, H.; Goerlich, D.; Sauerland, M.C.; Konstandin, N.P.; Dufour, A.; Schneider, S.; Neusser, M.; et al. Validation and refinement of the revised 2017 European LeukemiaNet genetic risk stratification of acute myeloid leukemia. Leukemia 2020, 34, 3161–3172. [Google Scholar] [CrossRef] [PubMed]
- Vrábel, D.; Pour, L.; Ševčíková, S. The Impact of NF-ΚB Signaling on Pathogenesis and Current Treatment Strategies in Multiple Myeloma. Blood Rev. 2019, 34, 56–66. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Function | Log2 (Expression Fold Change) | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| THP-1 | RPMI-8226 | CCRF-SB | CCRF-CEM | |||||||||||||||
| WNT signaling | ||||||||||||||||||
| AXIN2 | Negative regulator of WNT, expression activated by β-catenin [15] | −3.5 | −2.4 | −2.5 | −10.5 | |||||||||||||
| CCND1 | Activates the transition from G1 to S phase of the cell cycle. target gene of WNT pathway [15] | * | −2.1 | −2.9 | −3.4 | |||||||||||||
| CCND2 | Activates the transition from G1 to S phase of the cell cycle. target gene of WNT pathway [16] | −4.0 | −2.7 | −2.8 | −5.0 | |||||||||||||
| FOSL1 | Positive regulator of proliferation, target gene for WNT pathway [17] | * | * | 2.1 | 5.1 | |||||||||||||
| MMP7 | Regulates remodeling extracellular matrix, positive regulator of EMP, target gene of WNT pathway [18] | −5.9 | −2.5 | −2.7 | −2.3 | |||||||||||||
| MYC | Transcription factor, positive WNT pathway regulator [19] | −4.3 | −2.7 | −3.5 | −5.8 | |||||||||||||
| WISP | Positive WNT [20] pathway regulator, expression corresponds to pathway activity [20] | −9.8 | 3.1 | 3.6 | 2.9 | |||||||||||||
| PPARD | Positive WNT signaling regulator [21] | −2.9 | * | −2.5 | * | |||||||||||||
| DAB2 | Negative regulator of the WNT receptor [16] | −3.2 | −2.4 | * | * | |||||||||||||
| Notch signaling | ||||||||||||||||||
| HES5 | Notch pathway target gene [22], negative regulator of cell differentiation [23] | * | * | 2.6 | 4.0 | |||||||||||||
| HEY1 | Notch pathway target gene [24], positive regulator of cell differentiation [25] | −5.7 | * | 2.0 | 6.7 | |||||||||||||
| HEY2 | Notch pathway target gene [26], positive regulator of cell differentiation [27] | −8.1 | * | * | 3.9 | |||||||||||||
| HEYL | Notch pathway target gene [28], positive regulator of cell differentiation [29] | * | * | * | 4.9 | |||||||||||||
| ID1 | Negative regulator of Notch pathway [30], expression dependent on pathway activity [31] | −6.6 | 3.6 | 2.8 | 6 | |||||||||||||
| JAG1 | Positive regulator of Notch pathway [32] | * | * | * | 7.8 | |||||||||||||
| LFNG | Positive regulator of Notch pathway [33] | * | * | * | 3.3 | |||||||||||||
| NOTCH1 | Positive regulator of Notch pathway [34] | * | * | * | 4.4 | |||||||||||||
| Hedgehog-signaling | ||||||||||||||||||
| BCL2 | Target gene of Hedgehog pathway, negative regulator of apoptosis [35] | * | −2.7 | −2.0 | −2.9 | |||||||||||||
| PTCH1 | Negative regulator of Hedgehog pathway [36], expression dependent on pathway activity [37] | * | 2.7 | −3.0 | 4.3 | |||||||||||||
| BMP4 | Positive regulator of the Hedgehog pathway [38], expression depends on the activity of the pathway [39] | −4.5 | * | −3.5 | * | |||||||||||||
| WNT1 | Hedgehog pathway target gene [40], positive regulator of WNT pathway | −5 | −3.1 | * | −5.8 | |||||||||||||
| WNT2B | Hedgehog pathway target gene [40], positive regulator of WNT pathway | −3.1 | −5.1 | −2.9 | −4.5 | |||||||||||||
| WNT3A | Hedgehog pathway target gene [40], positive regulator of WNT pathway | * | * | * | −4.8 | |||||||||||||
| WNT5A | Hedgehog pathway target gene [40], positive regulator of WNT pathway | −13.9 | −2.5 | −2.4 | −5.7 | |||||||||||||
| WNT6 | Hedgehog pathway target gene [40], positive regulator of WNT pathway | −4.8 | −2.4 | * | * | |||||||||||||
| Hypoxia-induced signaling | ||||||||||||||||||
| ADM | HiF-1/HiF-2-dependent gene [41], vasodilator hormone. angiogenesis stimulant [42] | −2.9 | * | 2.5 | * | |||||||||||||
| CA9 | HiF-1-dependent gene [43], catalyzes CO2 hydration, trunk maintenance [44] | −13.3 | −2.5 | * | −2.3 | |||||||||||||
| EPO | HiF-1-dependent gene [45], regulates erythropoiesis and angiogenesis [46] | −15.5 | 2.3 | * | * | |||||||||||||
| HMOX1 | HiF-1-dependent gene, involved in heme metabolism and response to oxidative stress [47] | −2.5 | * | * | * | |||||||||||||
| SERPINE1 | HiF-1-dependent gene [48]. activator of hemostasis, cellular migration, angiogenesis [49] | −6.5 | −3.3 | * | * | |||||||||||||
| SLC2A1 | HiF-1-dependent gene [50], transports glucose to the cell [51] | −3.2 | 2.3 | −2.6 | * | |||||||||||||
| VEGFA | HiF-1-dependent gene [52], positive regulator of angiogenesis [53] | −2.9 | −2.7 | −3.2 | −5.3 | |||||||||||||
| LDH1 | Positive regulator of HiF-1 expression [54] | * | −2.3 | −2.5 | * | |||||||||||||
| NF-κΒ signaling | ||||||||||||||||||
| CSF1 | Genomic target of NF-κΒ pathway [55], acts as a stimulant of proliferation [56] | −2.9 | * | * | * | |||||||||||||
| IFNG | The target gene that positively regulates the immune response [57] | −4.4 | −2.6 | 3.2 | 5.9 | |||||||||||||
| BCL2A1 | A target gene of the signaling pathway [58], negatively regulates apoptosis [59] | * | −3.0 | * | * | |||||||||||||
| BIRC3 | A target gene of the signaling pathway [60], negatively regulates apoptosis [61] | * | * | −5.3 | −3.3 | |||||||||||||
| TNF | A positive regulator of the NF-kB pathway [62], expression is dependent on the activity of the pathway [63] | * | −3.0 | −2.4 | 3.2 | |||||||||||||
| STAT1 | Is a positive regulator of the NF-kB signaling pathway [64] | * | 6.0 | * | * | |||||||||||||
| CCL5 | A target gene of pathway [65], positively regulates the immune response [66] | * | * | * | 2.9 | |||||||||||||
| TGFβ signaling pathway | ||||||||||||||||||
| ATF4 | Target gene of TGFβ pathway, promotes stem-like phenotype in tumor cells [67] | −3.5 | −2.8 | −2.1 | * | |||||||||||||
| EMP1 | Target gene of TGFβ pathway, activates proliferation and cellular migration [68] | −4.1 | * | −3.9 | * | |||||||||||||
| MYC | Transcription factor, negative regulator of TGFβ [69] | −4.3 | −2.7 | −3.5 | −5.8 | |||||||||||||
| HERPUD1 | TGFβ target gene, component of ER-dependent degradation system [70] | * | −5 | −2.2 | * | |||||||||||||
| GADD45B | Target gene of TGFβ pathway, negatively regulates apoptosis, activates repair [71] | * | 3.2 | −7.9 | * | |||||||||||||
| IFRD1 | Target gene of TGFβ pathway, regulates immune response [72] | * | 2.3 | −2.4 | −2.0 | |||||||||||||
| CDKN1B | Target gene of TGFβ pathway [73], negatively regulates cell cycle progression [74] | * | * | 2.1 | * | |||||||||||||
| TNFSF10 | Target gene in TGFβ pathway [75], activates apoptosis [76] | * | * | 6.4 | ||||||||||||||
| PPARγ signaling pathway | ||||||||||||||||||
| CPT2 | PPARγ target gene, fatty acid oxidation enzyme [77] | −3.2 | 2.7 | −2.4 | * | |||||||||||||
| FABP1 | PPARγ target gene, transport of fatty acids [77] | −6.6 | * | * | * | |||||||||||||
| OLR1 | PPARγ target gene, mediates endocytosis of low-density lipoproteins [78] | −7.1 | −3.5 | −2.6 | 3.8 | |||||||||||||
| SLC27A4 | PPARγ target gene, mediates the transport of fatty acids inside the cell [79] | −4.6 | 2.4 | * | * | |||||||||||||
| SOCS3 | PPARγ target gene, negative regulator via JAK/STAT [80] | −2.1 | * | 2.0 | 3.2 | |||||||||||||
| ACSL3 | Target gene of PPARγ-pathway, mediates acyl-CoA joining to fatty acids [81] | * | 2.3 | −2.5 | * | |||||||||||||
| ACSL4 | Target gene of PPARγ-pathway, mediates acyl-CoA joining to fatty acids [82] | * | * | −2.4 | * | |||||||||||||
| ACSL5 | Target gene PPARγ pathways, mediates acyl-CoA joining to fatty acids. [77] | * | * | −2.1 | * | |||||||||||||
| P53 signaling pathway | ||||||||||||||||||
| BBC3 | Target gene of P53, activates apoptosis [83] | −3.2 | −2.8 | 2.7 | * | |||||||||||||
| EGFR | Expression mediated by p53 interaction with miR27a, tyrosine kinase receptor [84] | −23.0 | * | −3.9 | −3.9 | |||||||||||||
| FAS | Target gene of P53, activates external apoptosis pathway [85] | 2.4 | * | −2.4 | * | |||||||||||||
| BTG2 | Target gene of P53, inhibits cell cycle progression [86] | * | −3.4 | * | * | |||||||||||||
| PCNA | Target gene of P53, increases DNA polymerase efficiency [87] | * | 2.0 | −2.4 | * | |||||||||||||
| BAX | Target gene of P53, activates apoptosis [88] | * | * | * | −5.0 | |||||||||||||
| JAK/STAT signaling pathway | ||||||||||||||||||
| SOCS3 | Negative regulator of JAK/STAT pathway [89] | −2.1 | * | 2.0 | 3.0 | |||||||||||||
| FCER2 | Target gene of JAK/STAT pathway, stimulates growth and differentiation of B cells [90] | −16.6 | * | −4.4 | * | |||||||||||||
| CCND1 | Activates the transition from G1 to S phase of the cell cycle, target gene of the JAK/STAT pathway [91] | * | −2.2 | −2.0 | −3.4 | |||||||||||||
| IRF1 | Receptor that activates the JAK/STAT signaling pathway [92] | * | * | * | −7.6 | |||||||||||||
| CEBPD | Negative regulator of the JAK/STAT signaling pathway, induces cell arrest and apoptosis initiation [93] | * | * | 2.5 | * | |||||||||||||
| LRG1 | Positive JAK/STAT pathway regulator, suppresses apoptosis and induces G1 phase passage [94] | * | * | 2.6 | * | |||||||||||||
| Oxidative stress signaling pathway | ||||||||||||||||||
| GSR | Target gene of the pathway, reduces oxidized glutathione disulfide [95] | −3.3 | * | * | * | |||||||||||||
| HMOX1 | Target gene of the pathway, activates heme metabolism [96] | −2.5 | * | * | * | |||||||||||||
| TXNRD1 | Target gene of the signaling pathway, protection against oxidative stress [97] | * | 2.4 | 2.4 | * | |||||||||||||
| NQO1 | Target gene of the signaling pathway, protection against oxidative stress (quinone reduction) [98] | * | * | * | −2.5 | |||||||||||||
| Log2(fold change) | ||||||||||||||||||
| −10.0 | −9.0 | −8.0 | −7.0 | −6.0 | −5.0 | −4.0 | −3.0 | −2.0 | 2.0 | 3.0 | 4.0 | 5.0 | 6.0 | 7.0 | 8.0 | 9.0 | 10.0 | |
| Degree of Damage | Changes in Spleen Structure |
|---|---|
| Stage 0 | Intact organ |
| Stage 1 | The nodules of the white pulp are preserved, distinct contours are observed, and the structure with hermial centers is intact. The red pulp contains megacaryocytes (as a marker of hematopoiesis) and single tumor cells |
| Stage 2 | There is a change in the structure of the white pulp, but with distinct contours. Germinal centers are observed only in half of nodules. The red pulp contains groups of tumor cells and megacaryocytes |
| Stage 3 | The number of nodules of the white pulp is reduced to 2 or less in the field of view; the structure of nodules is changed. The red pulp is infiltrated by tumor cells, and no hemopoiesis has been observed |
| Stage 4 | The cytoarchitectonics of the spleen is broken, the boundary between the red and white pulp is missing, and there is an infiltration of tumor cells throughout |
| Degree of Damage | Changes in Organ Structure |
|---|---|
| Stage 0 | Intact organ |
| Stage 1 | Single tumor cells or small infiltrates |
| Stage 2 | Medium-sized tumor infiltrators, large-scale tumor infiltrates |
| Stage 3 | Large tumor infiltrates, liver lesions destroying cytoarchitectonics |
| Degree of Damage | Control Mice (7 Days) | WEHI-3 (7 Days) | Control Mice (21 Days) | WEHI-3 (21 Days) | WEHI-3 (21 Days) + CBL0137 (7–21 Days) |
|---|---|---|---|---|---|
| Spleens | |||||
| 0 | 5/5 | 0/10 | 5/5 | 0/10 | 5/10 |
| 1 | 0/5 | 8/10 | 0/5 | 2/10 | 1/10 |
| 2 | 0/5 | 2/10 | 0/5 | 2/10 | 2/10 |
| 3 | 0/5 | 0/10 | 0/5 | 4/10 | 2/10 |
| 4 | 0/5 | 0/10 | 0/5 | 2/10 | 0/10 |
| Livers | |||||
| 0 | 5/5 | 8/10 | 5/5 | 2/10 | 6/10 |
| 1 | 0/5 | 2/10 | 0/5 | 1/10 | 2/10 |
| 3 | 0/5 | 0/10 | 0/5 | 3/10 | 1/10 |
| 4 | 0/5 | 0/10 | 0/5 | 4/10 | 1/10 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fetisov, T.I.; Borunova, A.A.; Antipova, A.S.; Antoshina, E.E.; Trukhanova, L.S.; Gorkova, T.G.; Zuevskaya, S.N.; Maslov, A.; Gurova, K.; Gudkov, A.; et al. Targeting Features of Curaxin CBL0137 on Hematological Malignancies In Vitro and In Vivo. Biomedicines 2023, 11, 230. https://doi.org/10.3390/biomedicines11010230
Fetisov TI, Borunova AA, Antipova AS, Antoshina EE, Trukhanova LS, Gorkova TG, Zuevskaya SN, Maslov A, Gurova K, Gudkov A, et al. Targeting Features of Curaxin CBL0137 on Hematological Malignancies In Vitro and In Vivo. Biomedicines. 2023; 11(1):230. https://doi.org/10.3390/biomedicines11010230
Chicago/Turabian StyleFetisov, Timur I., Anna A. Borunova, Alina S. Antipova, Elena E. Antoshina, Lubov S. Trukhanova, Tatyana G. Gorkova, Svetlana N. Zuevskaya, Alexei Maslov, Katerina Gurova, Andrei Gudkov, and et al. 2023. "Targeting Features of Curaxin CBL0137 on Hematological Malignancies In Vitro and In Vivo" Biomedicines 11, no. 1: 230. https://doi.org/10.3390/biomedicines11010230
APA StyleFetisov, T. I., Borunova, A. A., Antipova, A. S., Antoshina, E. E., Trukhanova, L. S., Gorkova, T. G., Zuevskaya, S. N., Maslov, A., Gurova, K., Gudkov, A., Lesovaya, E. A., Belitsky, G. A., Yakubovskaya, M. G., & Kirsanov, K. I. (2023). Targeting Features of Curaxin CBL0137 on Hematological Malignancies In Vitro and In Vivo. Biomedicines, 11(1), 230. https://doi.org/10.3390/biomedicines11010230