

A Transient Inflammatory Response Induced by Lipopolysaccharide Infusion Lowers Markers of Endogenous Cholesterol and Bile Acid Synthesis in Healthy Normocholesterolemic Young Men
 ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects and Study Design
2.2. Blood Sampling
2.3. Biochemical Analysis
2.4. Non-Cholesterol Sterol and Oxysterol Concentrations
2.5. Statistical Analyses
3. Results
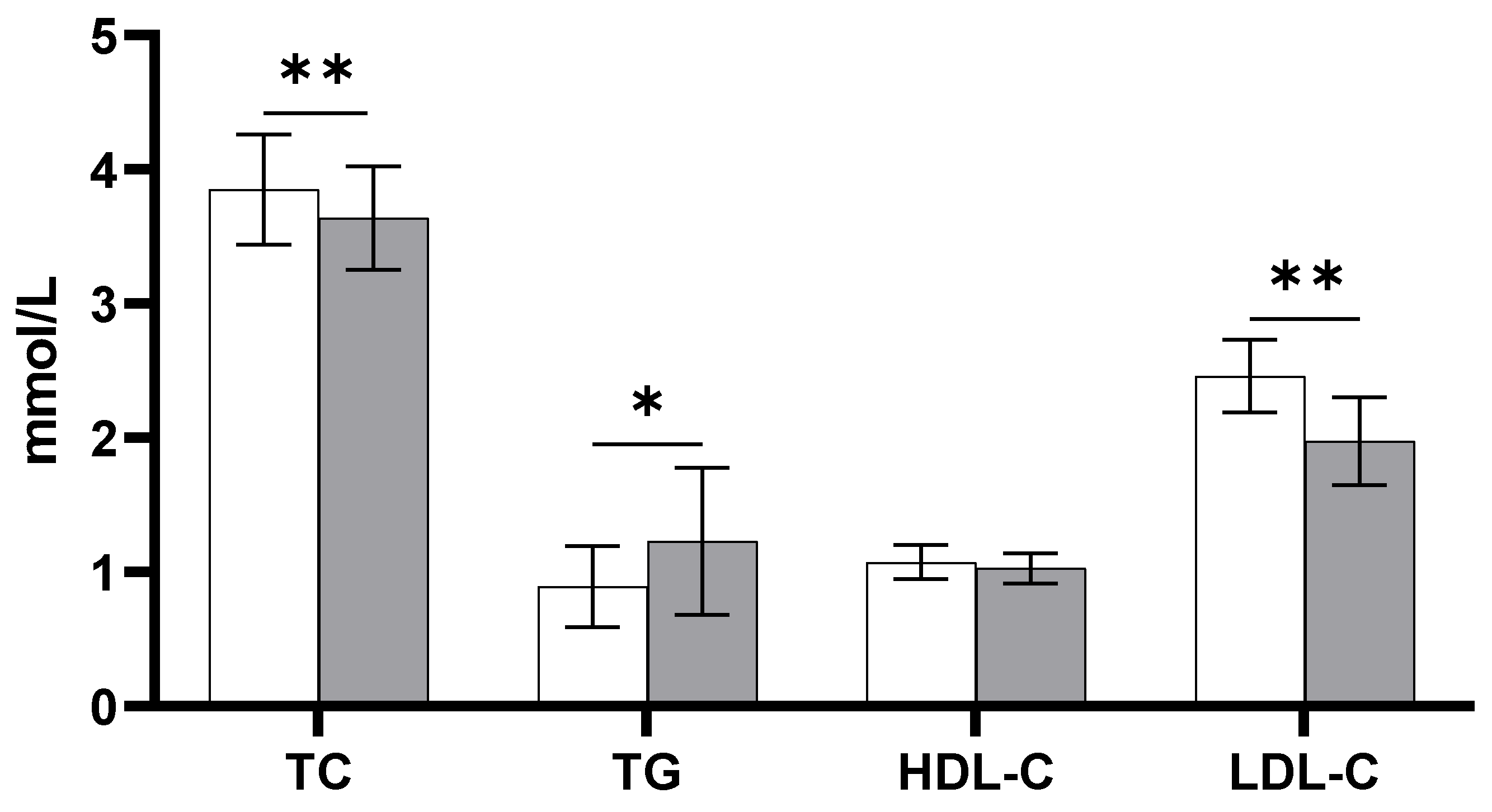
3.1. Lipids and Lipoproteins
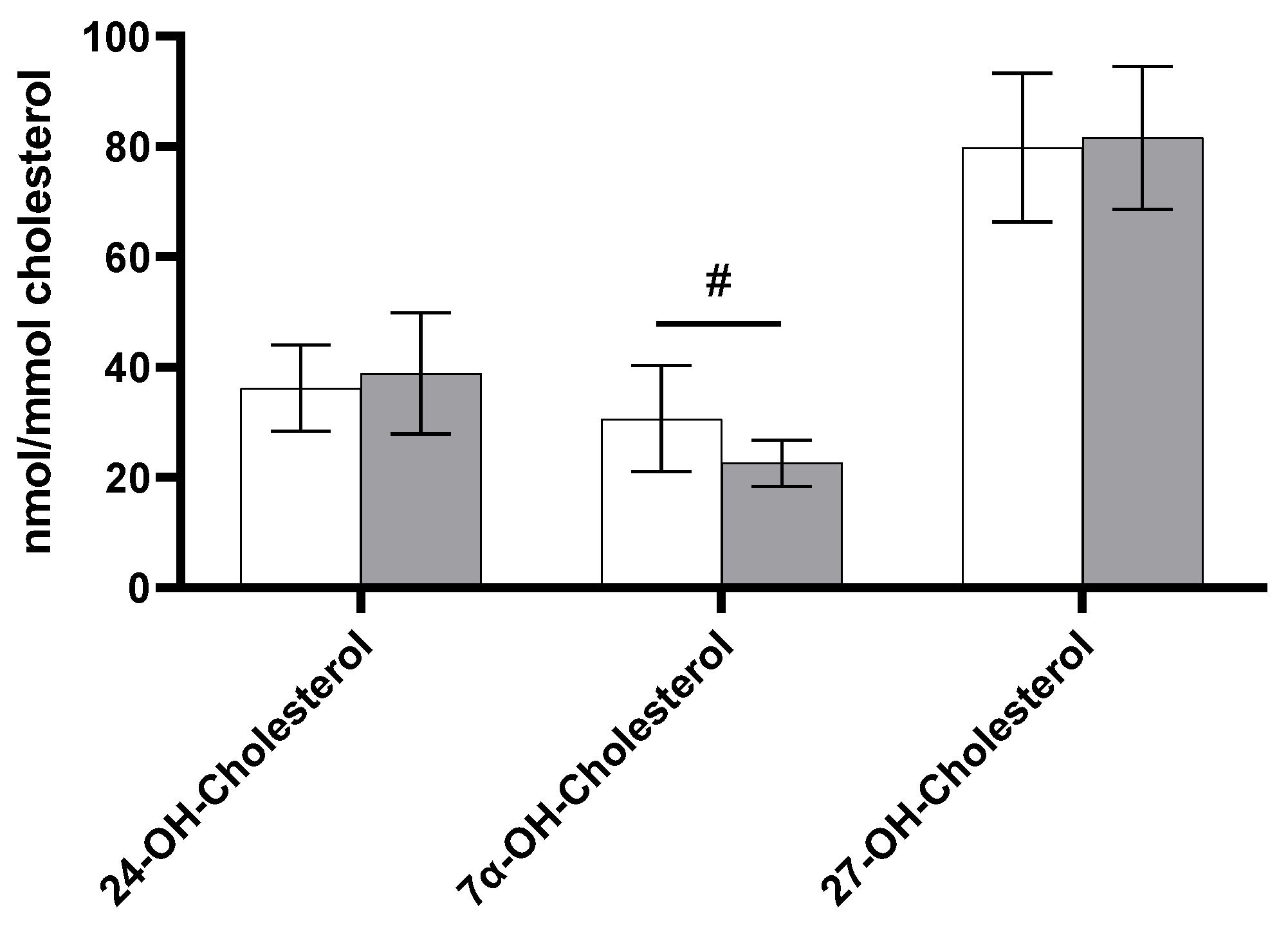
3.2. Non-Cholesterol Sterols and Oxysterols
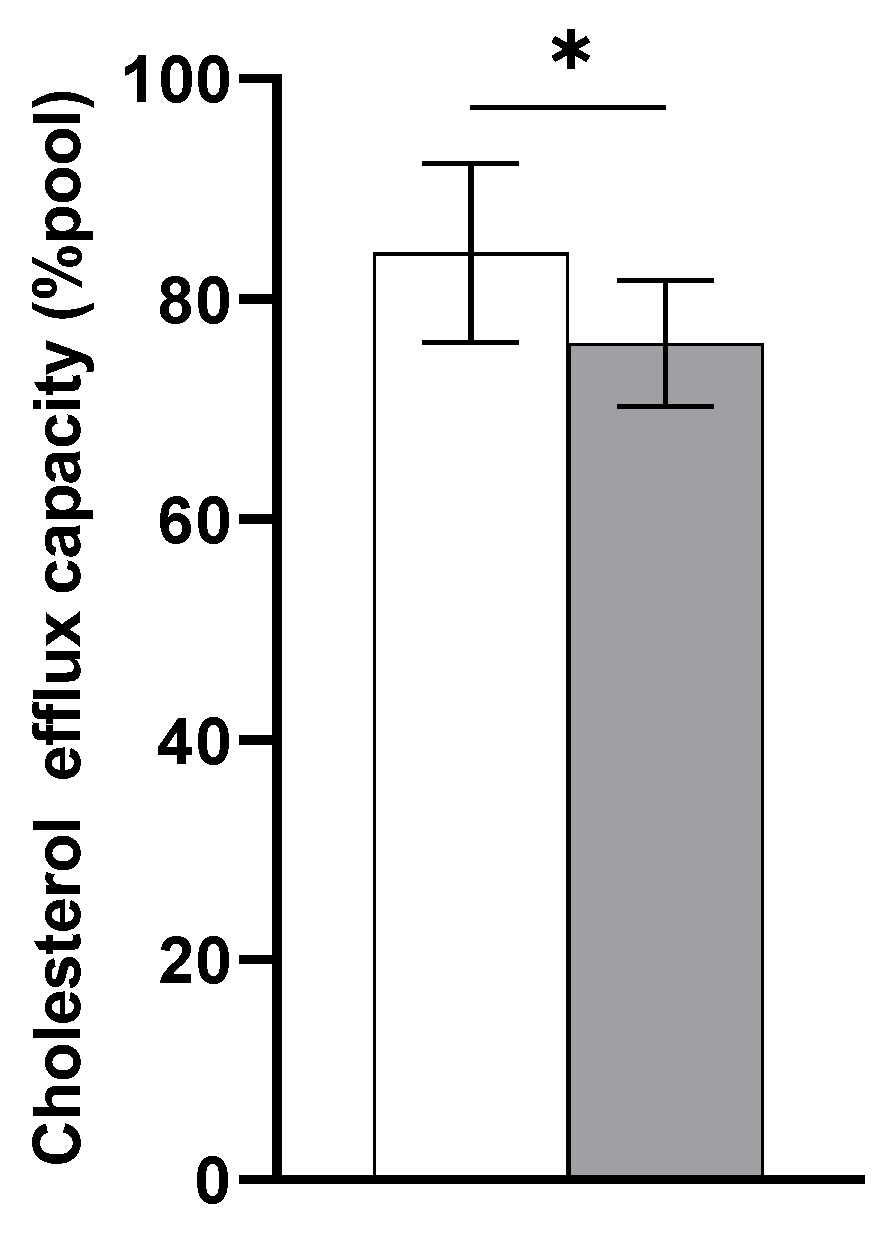
3.3. Inflammatory Responses
3.4. Correlations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Frostegard, J. Immunity, atherosclerosis and cardiovascular disease. BMC Med. 2013, 11, 117. [Google Scholar] [CrossRef] [PubMed]
- Weber, C.; Noels, H. Atherosclerosis: Current pathogenesis and therapeutic options. Nat. Med. 2011, 17, 1410–1422. [Google Scholar] [CrossRef] [PubMed]
- Ridker, P.M.; Luscher, T.F. Anti-inflammatory therapies for cardiovascular disease. Eur. Heart J. 2014, 35, 1782–1791. [Google Scholar] [CrossRef]
- Ridker, P.M.; Everett, B.M.; Thuren, T.; MacFadyen, J.G.; Chang, W.H.; Ballantyne, C.; Fonseca, F.; Nicolau, J.; Koenig, W.; Anker, S.D.; et al. Antiinflammatory Therapy with Canakinumab for Atherosclerotic Disease. N. Engl. J. Med. 2017, 377, 1119–1131. [Google Scholar] [CrossRef] [PubMed]
- Zanoli, L.; Briet, M.; Empana, J.P.; Cunha, P.G.; Maki-Petaja, K.M.; Protogerou, A.D.; Tedgui, A.; Touyz, R.M.; Schiffrin, E.L.; Spronck, B.; et al. Vascular consequences of inflammation: A position statement from the ESH Working Group on Vascular Structure and Function and the ARTERY Society. J. Hypertens. 2020, 38, 1682–1698. [Google Scholar] [CrossRef]
- Carpentier, Y.A.; Scruel, O. Changes in the concentration and composition of plasma lipoproteins during the acute phase response. Curr. Opin. Clin. Nutr. Metab. Care 2002, 5, 153–158. [Google Scholar] [CrossRef]
- Khovidhunkit, W.; Kim, M.S.; Memon, R.A.; Shigenaga, J.K.; Moser, A.H.; Feingold, K.R.; Grunfeld, C. Effects of infection and inflammation on lipid and lipoprotein metabolism: Mechanisms and consequences to the host. J. Lipid Res. 2004, 45, 1169–1196. [Google Scholar] [CrossRef]
- Feingold, K.R.; Grunfeld, C. The acute phase response inhibits reverse cholesterol transport. J. Lipid Res. 2010, 51, 682–684. [Google Scholar] [CrossRef]
- Feingold, K.R.; Grunfeld, C. Effect of inflammation on HDL structure and function. Curr. Opin. Lipidol. 2016, 27, 521–530. [Google Scholar] [CrossRef]
- Jahangiri, A.; de Beer, M.C.; Noffsinger, V.; Tannock, L.R.; Ramaiah, C.; Webb, N.R.; van der Westhuyzen, D.R.; de Beer, F.C. HDL remodeling during the acute phase response. Arter. Thromb. Vasc. Biol. 2009, 29, 261–267. [Google Scholar] [CrossRef]
- Zimetti, F.; De Vuono, S.; Gomaraschi, M.; Adorni, M.P.; Favari, E.; Ronda, N.; Ricci, M.A.; Veglia, F.; Calabresi, L.; Lupattelli, G. Plasma cholesterol homeostasis, HDL remodeling and function during the acute phase reaction. J. Lipid Res. 2017, 58, 2051–2060. [Google Scholar] [CrossRef] [PubMed]
- van Lier, D.; Geven, C.; Leijte, G.P.; Pickkers, P. Experimental human endotoxemia as a model of systemic inflammation. Biochimie 2019, 159, 99–106. [Google Scholar] [CrossRef]
- Perlee, D.; van Vught, L.A.; Scicluna, B.P.; Maag, A.; Lutter, R.; Kemper, E.M.; Veer, C.V.; Punchard, M.A.; Gonzalez, J.; Richard, M.P.; et al. Intravenous Infusion of Human Adipose Mesenchymal Stem Cells Modifies the Host Response to Lipopolysaccharide in Humans: A Randomized, Single-Blind, Parallel Group, Placebo Controlled Trial. Stem Cells 2018, 36, 1778–1788. [Google Scholar] [CrossRef] [PubMed]
- Hudgins, L.C.; Parker, T.S.; Levine, D.M.; Gordon, B.R.; Saal, S.D.; Jiang, X.C.; Seidman, C.E.; Tremaroli, J.D.; Lai, J.; Rubin, A.L. A single intravenous dose of endotoxin rapidly alters serum lipoproteins and lipid transfer proteins in normal volunteers. J. Lipid Res. 2003, 44, 1489–1498. [Google Scholar] [CrossRef] [PubMed]
- de la Llera Moya, M.; McGillicuddy, F.C.; Hinkle, C.C.; Byrne, M.; Joshi, M.R.; Nguyen, V.; Tabita-Martinez, J.; Wolfe, M.L.; Badellino, K.; Pruscino, L.; et al. Inflammation modulates human HDL composition and function in vivo. Atherosclerosis 2012, 222, 390–394. [Google Scholar] [CrossRef]
- Czubayko, F.; Beumers, B.; Lammsfuss, S.; Lutjohann, D.; von Bergmann, K. A simplified micro-method for quantification of fecal excretion of neutral and acidic sterols for outpatient studies in humans. J. Lipid Res. 1991, 32, 1861–1867. [Google Scholar] [CrossRef]
- Lutjohann, D.; Meese, C.O.; Crouse, J.R., 3rd; von Bergmann, K. Evaluation of deuterated cholesterol and deuterated sitostanol for measurement of cholesterol absorption in humans. J. Lipid Res. 1993, 34, 1039–1046. [Google Scholar] [CrossRef]
- Stellaard, F.; Lutjohann, D. The Interpretation of Cholesterol Balance Derived Synthesis Data and Surrogate Noncholesterol Plasma Markers for Cholesterol Synthesis under Lipid Lowering Therapies. Cholesterol 2017, 2017, 5046294. [Google Scholar] [CrossRef]
- Lutjohann, D.; Bjorkhem, I.; Friedrichs, S.; Kerksiek, A.; Lovgren-Sandblom, A.; Geilenkeuser, W.J.; Ahrends, R.; Andrade, I.; Ansorena, D.; Astiasaran, I.; et al. First international descriptive and interventional survey for cholesterol and non-cholesterol sterol determination by gas- and liquid-chromatography-Urgent need for harmonisation of analytical methods. J. Steroid Biochem. Mol. Biol. 2019, 190, 115–125. [Google Scholar] [CrossRef]
- Lutjohann, D.; Bjorkhem, I.; Friedrichs, S.; Kerksiek, A.; Geilenkeuser, W.J.; Lovgren-Sandblom, A.; Ansorena, D.; Astiasaran, I.; Baila-Rueda, L.; Barriuso, B.; et al. International descriptive and interventional survey for oxycholesterol determination by gas- and liquid-chromatographic methods. Biochimie 2018, 153, 26–32. [Google Scholar] [CrossRef]
- Spann, N.J.; Garmire, L.X.; McDonald, J.G.; Myers, D.S.; Milne, S.B.; Shibata, N.; Reichart, D.; Fox, J.N.; Shaked, I.; Heudobler, D.; et al. Regulated accumulation of desmosterol integrates macrophage lipid metabolism and inflammatory responses. Cell 2012, 151, 138–152. [Google Scholar] [CrossRef]
- Friedewald, W.T.; Levy, R.I.; Fredrickson, D.S. Estimation of the concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin. Chem. 1972, 18, 499–502. [Google Scholar] [CrossRef] [PubMed]
- Scharnagl, H.; Heuschneider, C.; Sailer, S.; Kleber, M.E.; Marz, W.; Ritsch, A. Decreased cholesterol efflux capacity in patients with low cholesteryl ester transfer protein plasma levels. Eur. J. Clin. Investig. 2014, 44, 395–401. [Google Scholar] [CrossRef]
- Mackay, D.S.; Jones, P.J.; Myrie, S.B.; Plat, J.; Lutjohann, D. Methodological considerations for the harmonization of non-cholesterol sterol bio-analysis. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2014, 957, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Sosic-Jurjevic, B.; Lutjohann, D.; Renko, K.; Filipovic, B.; Radulovic, N.; Ajdzanovic, V.; Trifunovic, S.; Nestorovic, N.; Zivanovic, J.; Manojlovic Stojanoski, M.; et al. The isoflavones genistein and daidzein increase hepatic concentration of thyroid hormones and affect cholesterol metabolism in middle-aged male rats. J. Steroid Biochem. Mol. Biol. 2019, 190, 1–10. [Google Scholar] [CrossRef]
- Feingold, K.R.; Staprans, I.; Memon, R.A.; Moser, A.H.; Shigenaga, J.K.; Doerrler, W.; Dinarello, C.A.; Grunfeld, C. Endotoxin rapidly induces changes in lipid metabolism that produce hypertriglyceridemia: Low doses stimulate hepatic triglyceride production while high doses inhibit clearance. J. Lipid Res. 1992, 33, 1765–1776. [Google Scholar] [CrossRef]
- Hardardottir, I.; Moser, A.H.; Memon, R.; Grunfeld, C.; Feingold, K.R. Effects of TNF, IL-1, and the combination of both cytokines on cholesterol metabolism in Syrian hamsters. Lymphokine Cytokine Res. 1994, 13, 161–166. [Google Scholar] [PubMed]
- McGillicuddy, F.C.; de la Llera Moya, M.; Hinkle, C.C.; Joshi, M.R.; Chiquoine, E.H.; Billheimer, J.T.; Rothblat, G.H.; Reilly, M.P. Inflammation impairs reverse cholesterol transport in vivo. Circulation 2009, 119, 1135–1145. [Google Scholar] [CrossRef]
- Khera, A.V.; Cuchel, M.; de la Llera-Moya, M.; Rodrigues, A.; Burke, M.F.; Jafri, K.; French, B.C.; Phillips, J.A.; Mucksavage, M.L.; Wilensky, R.L.; et al. Cholesterol efflux capacity, high-density lipoprotein function, and atherosclerosis. N. Engl. J. Med. 2011, 364, 127–135. [Google Scholar] [CrossRef]
- Annema, W.; Nijstad, N.; Tolle, M.; de Boer, J.F.; Buijs, R.V.; Heeringa, P.; van der Giet, M.; Tietge, U.J. Myeloperoxidase and serum amyloid A contribute to impaired in vivo reverse cholesterol transport during the acute phase response but not group IIA secretory phospholipase A(2). J. Lipid Res. 2010, 51, 743–754. [Google Scholar] [CrossRef]
- Holzer, M.; Wolf, P.; Curcic, S.; Birner-Gruenberger, R.; Weger, W.; Inzinger, M.; El-Gamal, D.; Wadsack, C.; Heinemann, A.; Marsche, G. Psoriasis alters HDL composition and cholesterol efflux capacity. J. Lipid Res. 2012, 53, 1618–1624. [Google Scholar] [CrossRef] [PubMed]
- Pussinen, P.J.; Jauhiainen, M.; Vilkuna-Rautiainen, T.; Sundvall, J.; Vesanen, M.; Mattila, K.; Palosuo, T.; Alfthan, G.; Asikainen, S. Periodontitis decreases the antiatherogenic potency of high density lipoprotein. J. Lipid Res. 2004, 45, 139–147. [Google Scholar] [CrossRef]
- Ronda, N.; Favari, E.; Borghi, M.O.; Ingegnoli, F.; Gerosa, M.; Chighizola, C.; Zimetti, F.; Adorni, M.P.; Bernini, F.; Meroni, P.L. Impaired serum cholesterol efflux capacity in rheumatoid arthritis and systemic lupus erythematosus. Ann. Rheum. Dis. 2014, 73, 609–615. [Google Scholar] [CrossRef] [PubMed]
- Siegel, M.O.; Borkowska, A.G.; Dubrovsky, L.; Roth, M.; Welti, R.; Roberts, A.D.; Parenti, D.M.; Simon, G.L.; Sviridov, D.; Simmens, S.; et al. HIV infection induces structural and functional changes in high density lipoproteins. Atherosclerosis 2015, 243, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Ettinger, W.H.; Varma, V.K.; Sorci-Thomas, M.; Parks, J.S.; Sigmon, R.C.; Smith, T.K.; Verdery, R.B. Cytokines decrease apolipoprotein accumulation in medium from Hep G2 cells. Arterioscler. Thromb. 1994, 14, 8–13. [Google Scholar] [CrossRef]
- Feingold, K.R.; Serio, M.K.; Adi, S.; Moser, A.H.; Grunfeld, C. Tumor necrosis factor stimulates hepatic lipid synthesis and secretion. Endocrinology 1989, 124, 2336–2342. [Google Scholar] [CrossRef]
- Feingold, K.R.; Soued, M.; Serio, M.K.; Moser, A.H.; Dinarello, C.A.; Grunfeld, C. Multiple cytokines stimulate hepatic lipid synthesis in vivo. Endocrinology 1989, 125, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Feingold, K.R.; Soued, M.; Staprans, I.; Gavin, L.A.; Donahue, M.E.; Huang, B.J.; Moser, A.H.; Gulli, R.; Grunfeld, C. Effect of tumor necrosis factor (TNF) on lipid metabolism in the diabetic rat. Evidence that inhibition of adipose tissue lipoprotein lipase activity is not required for TNF-induced hyperlipidemia. J. Clin. Investig. 1989, 83, 1116–1121. [Google Scholar] [CrossRef]
- Liebergall, S.R.; Angdisen, J.; Chan, S.H.; Chang, Y.; Osborne, T.F.; Koeppel, A.F.; Turner, S.D.; Schulman, I.G. Inflammation Triggers Liver X Receptor-Dependent Lipogenesis. Mol. Cell. Biol. 2020, 40, e00364-19. [Google Scholar] [CrossRef]
- Jelinek, D.F.; Andersson, S.; Slaughter, C.A.; Russell, D.W. Cloning and regulation of cholesterol 7 alpha-hydroxylase, the rate-limiting enzyme in bile acid biosynthesis. J. Biol. Chem. 1990, 265, 8190–8197. [Google Scholar] [CrossRef]
- Feingold, K.R.; Spady, D.K.; Pollock, A.S.; Moser, A.H.; Grunfeld, C. Endotoxin, TNF, and IL-1 decrease cholesterol 7 alpha-hydroxylase mRNA levels and activity. J. Lipid Res. 1996, 37, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Dikopoulos, N.; Weidenbach, H.; Adler, G.; Schmid, R.M. Lipopolysaccharide represses cholesterol 7-alpha hydroxylase and induces binding activity to the bile acid response element II. Eur. J. Clin. Investig. 2003, 33, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Janowski, B.A.; Willy, P.J.; Devi, T.R.; Falck, J.R.; Mangelsdorf, D.J. An oxysterol signalling pathway mediated by the nuclear receptor LXR alpha. Nature 1996, 383, 728–731. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, J.M.; Kliewer, S.A.; Moore, L.B.; Smith-Oliver, T.A.; Oliver, B.B.; Su, J.L.; Sundseth, S.S.; Winegar, D.A.; Blanchard, D.E.; Spencer, T.A.; et al. Activation of the nuclear receptor LXR by oxysterols defines a new hormone response pathway. J. Biol. Chem. 1997, 272, 3137–3140. [Google Scholar] [CrossRef] [PubMed]
- Peet, D.J.; Turley, S.D.; Ma, W.; Janowski, B.A.; Lobaccaro, J.M.; Hammer, R.E.; Mangelsdorf, D.J. Cholesterol and bile acid metabolism are impaired in mice lacking the nuclear oxysterol receptor LXR alpha. Cell 1998, 93, 693–704. [Google Scholar] [CrossRef]
- Yang, C.; McDonald, J.G.; Patel, A.; Zhang, Y.; Umetani, M.; Xu, F.; Westover, E.J.; Covey, D.F.; Mangelsdorf, D.J.; Cohen, J.C.; et al. Sterol intermediates from cholesterol biosynthetic pathway as liver X receptor ligands. J. Biol. Chem. 2006, 281, 27816–27826. [Google Scholar] [CrossRef] [PubMed]
- Lund, E.G.; Peterson, L.B.; Adams, A.D.; Lam, M.H.; Burton, C.A.; Chin, J.; Guo, Q.; Huang, S.; Latham, M.; Lopez, J.C.; et al. Different roles of liver X receptor alpha and beta in lipid metabolism: Effects of an alpha-selective and a dual agonist in mice deficient in each subtype. Biochem. Pharmacol. 2006, 71, 453–463. [Google Scholar] [CrossRef] [PubMed]
- Tall, A.R. Protease variants, LDL, and coronary heart disease. N. Engl. J. Med. 2006, 354, 1310–1312. [Google Scholar] [CrossRef]
- Tall, A.R.; Yvan-Charvet, L. Cholesterol, inflammation and innate immunity. Nat. Rev. Immunol. 2015, 15, 104–116. [Google Scholar] [CrossRef]
- Zhang, X.; McDonald, J.G.; Aryal, B.; Canfran-Duque, A.; Goldberg, E.L.; Araldi, E.; Ding, W.; Fan, Y.; Thompson, B.M.; Singh, A.K.; et al. Desmosterol suppresses macrophage inflammasome activation and protects against vascular inflammation and atherosclerosis. Proc. Natl. Acad. Sci. USA 2021, 118, e2107682118. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mean ± SD | p-Value | ||
|---|---|---|---|
| Albumin (g/L) | Baseline | 38.43 ± 1.60 | |
| 24 h | 40.59 ± 2.32 | 0.033 | |
| Change | 2.16 ± 2.31 | ||
| CRP (mg/L) | Baseline | 3.01 ± 4.42 | |
| 24 h | 24.50 ± 6.83 | <0.001 | |
| Change | 21.49 ± 2.21 | ||
| TNFα (pg/mL) | Baseline | 3.02 ± 1.30 | |
| 24 h | 3.76 ± 1.37 | 0.002 | |
| Change | 0.74 ± 0.45 | ||
| IL-8 (pg/mL) | Baseline | 0.32 ± 0.22 | |
| 24 h | 0.60 ± 0.46 | 0.110 | |
| Change | 0.29 ± 0.45 | ||
| IL-10 (pg/mL) | Baseline | 0.19 ± 0.09 | |
| 24 h | 0.23 ± 0.13 | 0.457 | |
| Change | 0.04 ± 0.15 | ||
| IL12p40 (pg/mL) | Baseline | 1.86 ± 1.13 | |
| 24 h | 2.25 ± 2.12 | 0.355 | |
| Change | 0.39 ± 1.12 | ||
| SAA (mg/L) | Baseline | 5.40 ± 1.89 | |
| 24 h | 6.64 ± 1.95 | 0.017 | |
| Change | 1.24 ± 1.13 | ||
| MPO (ng/mL) | Baseline | 2.96± 1.05 | |
| 24 h | 5.70 ± 1.56 | <0.001 | |
| Change | 2.73 ± 1.33 |
| Variable | Variable | Correlation | p-Value |
|---|---|---|---|
| Baseline desmosterol | 24 h CRP | 0.849 | 0.008 |
| Baseline desmosterol | ∆CRP | 0.829 | 0.011 |
| Baseline 27-OH-cholesterol | 24 h IL8 | 0.771 | 0.025 |
| Baseline cholestanol | iMAX MPO | 0.758 | 0.029 |
| Baseline desmosterol | iMAX TNFα | 0.917 | 0.010 |
| Baseline 7α-OH-cholesterol | iMAX IL-8 | 0.766 | 0.027 |
| Baseline 7α-OH-cholesterol | iMAX IL-6 | 0.763 | 0.028 |
| Baseline 7α-OH-cholesterol | iMAX TNFα | 0.814 | 0.049 |
| ∆desmosterol | iMAX IL-8 | −0.761 | 0.028 |
| ∆7α-OH-cholesterol | iMAX IL-8 | −0.856 | 0.007 |
| ∆7α-OH-cholesterol | iMAX IL-6 | −0.751 | 0.032 |
| ∆7α-OH-cholesterol | iMAX TNFα | −0.821 | 0.045 |
| Baseline 27-OH-cholesterol | iAUC-TNFα | 0.765 | 0.027 |
| Baseline 7α-OH-cholesterol | iAUC-IL6 | 0.869 | 0.005 |
| ∆7α-OH-cholesterol | iAUC-IL6 | −0.904 | 0.002 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mashnafi, S.; Baumgartner, S.; Mensink, R.P.; Perlee, D.; van Vught, L.A.; Lütjohann, D.; Plat, J. A Transient Inflammatory Response Induced by Lipopolysaccharide Infusion Lowers Markers of Endogenous Cholesterol and Bile Acid Synthesis in Healthy Normocholesterolemic Young Men. Biomedicines 2023, 11, 126. https://doi.org/10.3390/biomedicines11010126
Mashnafi S, Baumgartner S, Mensink RP, Perlee D, van Vught LA, Lütjohann D, Plat J. A Transient Inflammatory Response Induced by Lipopolysaccharide Infusion Lowers Markers of Endogenous Cholesterol and Bile Acid Synthesis in Healthy Normocholesterolemic Young Men. Biomedicines. 2023; 11(1):126. https://doi.org/10.3390/biomedicines11010126
Chicago/Turabian StyleMashnafi, Sultan, Sabine Baumgartner, Ronald P. Mensink, Desiree Perlee, Lonneke A. van Vught, Dieter Lütjohann, and Jogchum Plat. 2023. "A Transient Inflammatory Response Induced by Lipopolysaccharide Infusion Lowers Markers of Endogenous Cholesterol and Bile Acid Synthesis in Healthy Normocholesterolemic Young Men" Biomedicines 11, no. 1: 126. https://doi.org/10.3390/biomedicines11010126
APA StyleMashnafi, S., Baumgartner, S., Mensink, R. P., Perlee, D., van Vught, L. A., Lütjohann, D., & Plat, J. (2023). A Transient Inflammatory Response Induced by Lipopolysaccharide Infusion Lowers Markers of Endogenous Cholesterol and Bile Acid Synthesis in Healthy Normocholesterolemic Young Men. Biomedicines, 11(1), 126. https://doi.org/10.3390/biomedicines11010126