
Contribution of Adenosine in the Physiological Changes and Injuries Secondary to Exposure to Extreme Oxygen Pressure in Healthy Subjects


 ,
,  ,
, 
{kind=link}
{kind=link}
Abstract
:1. Introduction
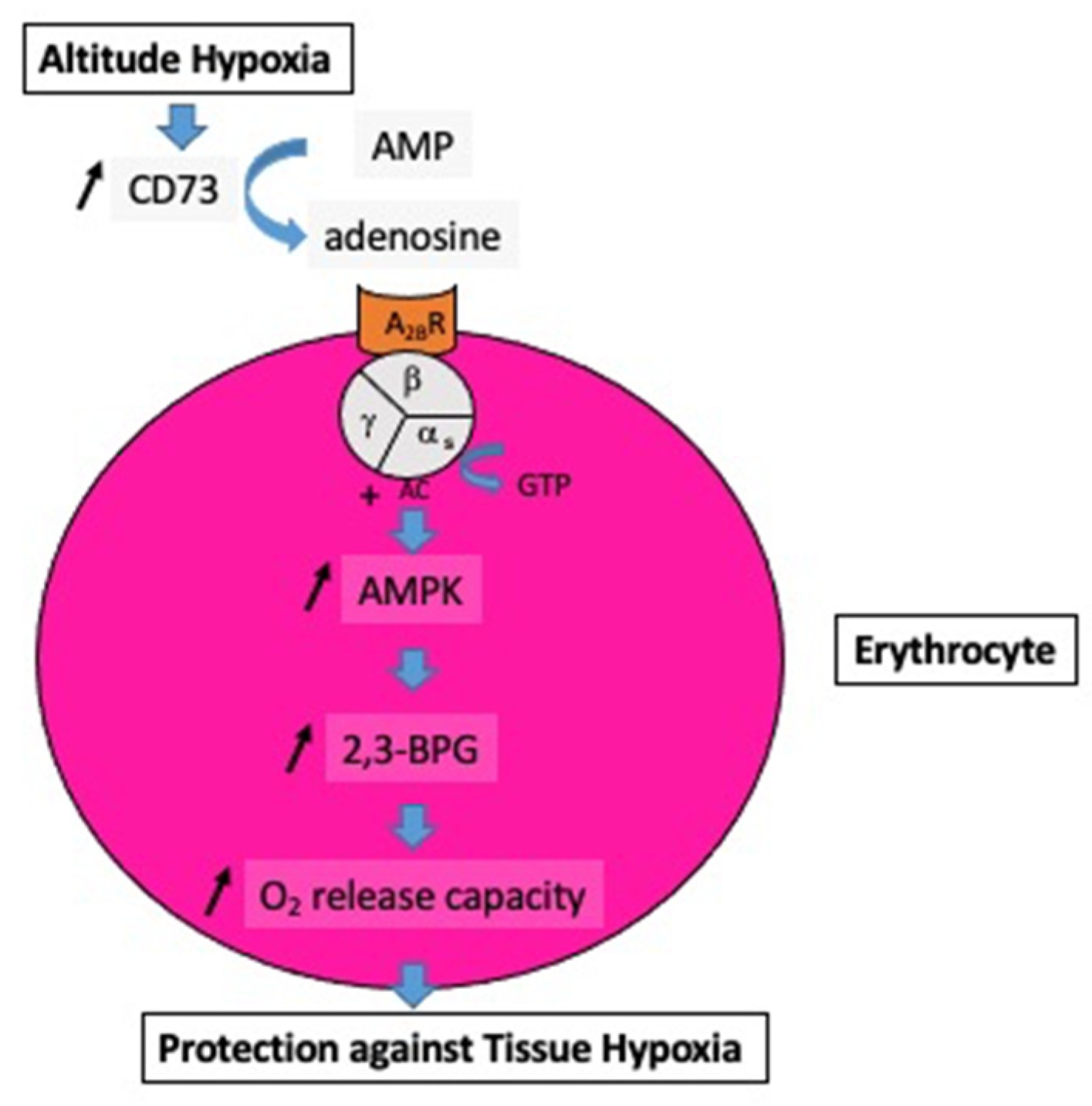
1.1. Altitude Hypoxia
1.2. Acute Altitude Illness
1.3. Chronic Hypoxia: Life at Altitude
2. Hyperoxia
2.1. Cardio-Vascular Changes
2.2. Oxygen Toxicity
2.2.1. Nervous System Toxicity
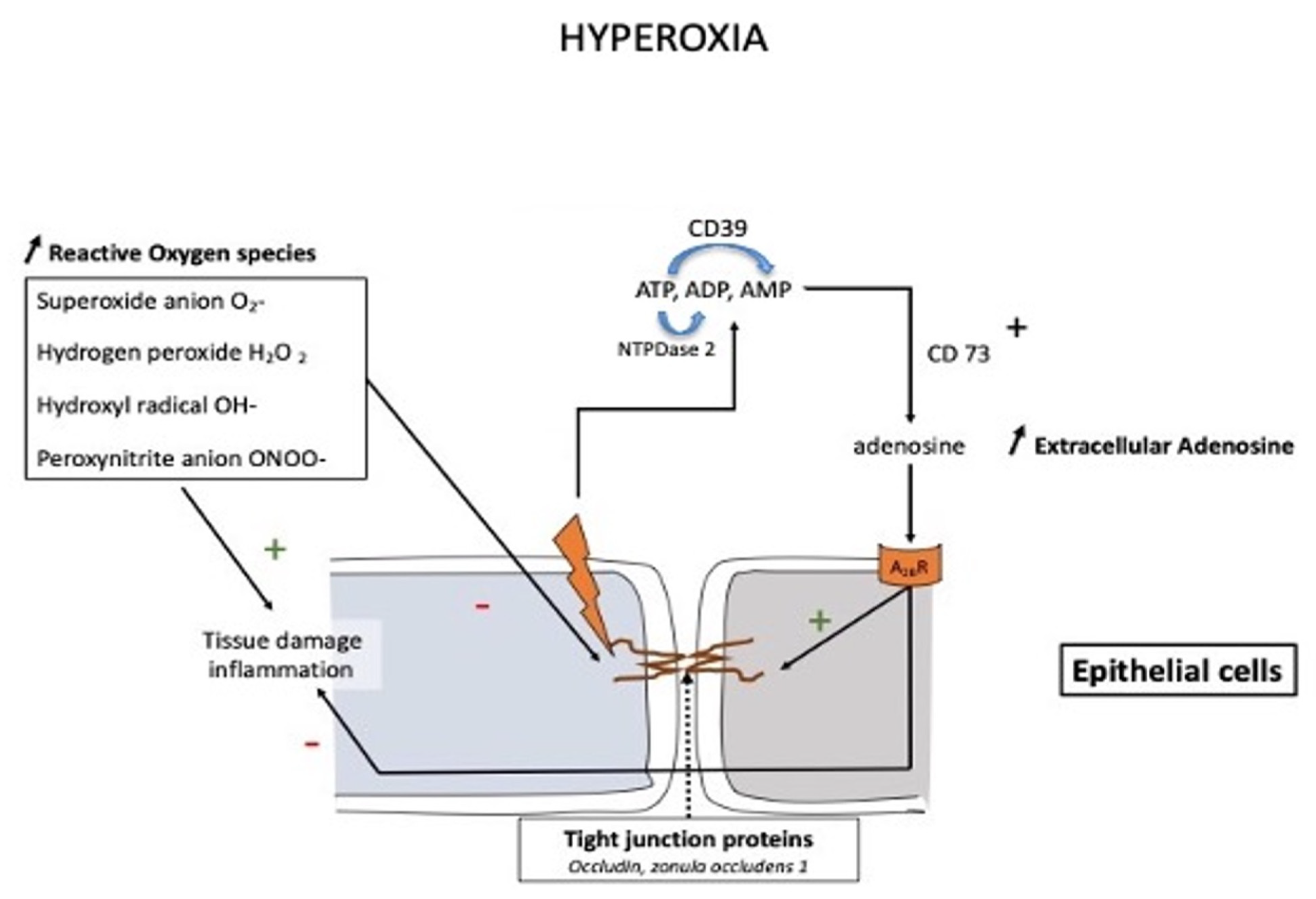
2.2.2. Lung Toxicity
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AMP | Adenosoine monophosphate |
| ADP | Adenosine diphosphate |
| AMPK | AMP kinase |
| ATP | Adenosine triphosphate |
| ENT1 | Equilibrative nucleoside transporter 1 |
| CD 73 | Ecto 5′ nucleotidase |
| CD39 (or NTPDase 1) | Ectonucleoside triphosphates diphosphohydrolase 1 |
| HIF | Hypoxia–inducible factor |
| KATP | ATP sensitive potassium channels |
| KV | Voltage gated potassium channels |
| NTPDase 2 | Ectonucleoside triphosphates diphosphohydrolase 2 |
| P1 receptors | Receptors for adenosine |
| P2 receptors | Receptors for nucleotides |
| PKC | C-protein kinase |
| 2,3-BPG | 2,3-Bisphosphoglycerate |
References
- Grenz, A.; Homann, D.; Eltzschig, H.K. Extracellular adenosine: A safety signal that dampens hypoxia-induced inflammation during ischemia. Antioxid. Redox. Signal. 2011, 15, 2221–2234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eltzschig, H.K.; Karhausen, J.; Kempf, V.A. Acute oxygen-sensing mechanisms. N. Engl. J. Med. 2006, 354, 975–977. [Google Scholar] [PubMed]
- Joulia, F.; Coulange, M.; Lemaitre, F.; Costalat, G.; Franceschi, F.; Gariboldi, V.; Nee, L.; Fromonot, J.; Bruzzese, L.; Gravier, G.; et al. Plasma adenosine release is associated with bradycardia and transient loss of consciousness during experimental breath-hold diving. Int. J. Cardiol. 2013, 168, e138–e141. [Google Scholar] [CrossRef] [PubMed]
- Bruzzese, L.; Rostain, J.C.; Née, L.; Condo, J.; Mottola, G.; Adjriou, N.; Mercier, L.; Berge-Lefranc, J.L.; Fromonot, J.; Kipson, N.; et al. Effect of hyperoxic and hyperbaric conditions on the adenosinergic pathway and CD26 expression in rat. J. Appl. Physiol. 2015, 119, 140–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fromonot, J.; Chaumet, G.; Gavarry, O.; Rostain, J.C.; Lucciano, M.; Joulia, F.; Brignole, M.; Deharo, J.C.; Guieu, R.; Boussuges, A. Hyperoxia Improves Hemodynamic Status During Head-up Tilt Testing in Healthy Volunteers: A Randomized Study. Medicine 2016, 95, e2876. [Google Scholar] [CrossRef] [PubMed]
- Morote-Garcia, J.C.; Rosenberger, P.; Kuhlicke, J.; Eltzschig, H.K. HIF-1-dependent repression of adenosine kinase attenuates hypoxia-induced vascular leak. Blood 2008, 111, 5571–5580. [Google Scholar] [CrossRef] [Green Version]
- Hellsten, Y.; Nyberg, M.; Mortensen, S.P. Contribution of intravascular versus interstitial purines and nitric oxide in the regulation of exercise hyperaemia in humans. J. Physiol. 2012, 590, 5015–5023. [Google Scholar] [CrossRef] [Green Version]
- Peacock, A.J. ABC of oxygen: Oxygen at high altitude. BMJ 1998, 317, 1063–1066. [Google Scholar] [CrossRef]
- Brugniaux, J.V.; Hodges, A.N.; Hanly, P.J.; Poulin, M.J. Cerebrovascular responses to altitude. Respir. Physiol. Neurobiol. 2007, 158, 212–223. [Google Scholar] [CrossRef]
- Winn, H.R.; Rubio, R.; Berne, R.M. Brain adenosine concentration during hypoxia in rats. Am. J. Physiol. 1981, 241, H235–H242. [Google Scholar] [CrossRef]
- Le, G.; Essackjee, H.; Ballard, H. Intracellular adenosine formation and release by freshly-isolated vascular endothelial cells from rat skeletal muscle: Effects of hypoxia and/or acidosis. Biochem. Biophys. Res. Commun. 2014, 450, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Idzko, M.; Ferrari, D.; Eltzschig, H.K. Nucleotide signalling during inflammation. Nature 2014, 509, 310–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sumi, Y.; Woehrle, T.; Chen, Y.; Yao, Y.; Li, A.; Junger, W.G. Adrenergic receptor activation involves ATP release and feedback through purinergic receptors. Am. J. Physiol. Physiol. 2010, 299, C1118–C1126. [Google Scholar] [CrossRef] [Green Version]
- Song, A.; Zhang, Y.; Han, L.; Yegutkin, G.G.; Liu, H.; Sun, K.; D’Alessandro, A.; Li, J.; Karmouty-Quintana, H.; Iriyama, T.; et al. Erythrocytes retain hypoxic adenosine response for faster acclimatization upon re-ascent. Nat. Commun. 2017, 8, 14108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Zhang, Y.; Wu, H.; D’Alessandro, A.; Yegutkin, G.G.; Song, A.; Sun, K.; Li, J.; Cheng, N.Y.; Huang, A.; et al. Beneficial Role of Erythrocyte Adenosine A2B Receptor-Mediated AMP-Activated Protein Kinase Activation in High-Altitude Hypoxia. Circulation 2016, 134, 405–421. [Google Scholar] [CrossRef] [PubMed]
- Erlinge, D.; Burnstock, G. P2 receptors in cardiovascular regulation and disease. Purinergic Signal. 2008, 4, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Marlinge, M.; Vairo, D.; Bertaud, A.; Vernet, C.; Chefrour, M.; Bruzzese, L.; Chaptal, M.C.; Mottola, G.; Boussuges, A.; Risso, J.J.; et al. Adaptative mechanism of the equilibrative nucleoside transporter 1 (ENT-1) and blood adenosine levels in elite freedivers. Eur. J. Appl. Physiol. 2021, 121, 279–285. [Google Scholar] [CrossRef]
- Burnstock, G. Purinergic Signaling in the Cardiovascular System. Circ. Res. 2017, 120, 207–228. [Google Scholar] [CrossRef] [Green Version]
- Borea, P.A.; Gessi, S.; Merighi, S.; Vincenzi, F.; Varani, K. Pharmacology of Adenosine Receptors: The State of the Art. Physiol. Rev. 2018, 98, 1591–1625. [Google Scholar] [CrossRef]
- Beukers, M.W.; den Dulk, H.; van Tilburg, E.W.; Brouwer, J.; Ijzerman, A.P. Why are A(2B) receptors low-affinity adenosine receptors? Mutation of Asn273 to Tyr increases affinity of human A(2B) receptor for 2-(1-Hexynyl)adenosine. Mol. Pharmacol. 2000, 58, 1349–1356. [Google Scholar] [CrossRef] [Green Version]
- Görlach, A. Control of adenosine transport by hypoxia. Circ. Res. 2005, 97, 1–3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laborit, H.; Bonifacj, J.F. Action of adenosine on energy metabolism and on glucose-6-phosphate dehydrogenase in rat brains. Res. Commun. Chem. Pathol. Pharmacol. 1984, 44, 123–130. [Google Scholar] [PubMed]
- Coney, A.; Marshall, J.M. Role of adenosine and its receptors in the vasodilatation induced in the cerebral cortex of the rat by systemic hypoxia. J. Physiol. 1998, 509, 507–518. [Google Scholar] [CrossRef]
- Feoktistov, I.; Ryzhov, S.; Zhong, H.; Goldstein, A.E.; Matafonov, A.; Zeng, D.; Biaggioni, I. Hypoxia Modulates Adenosine Receptors in Human Endothelial and Smooth Muscle Cells Toward an A2B Angiogenic Phenotype. Hypertension 2004, 44, 649–654. [Google Scholar] [CrossRef] [Green Version]
- Hori, M.; Kitakaze, M. Adenosine, the heart, and coronary circulation. Hypertension. 1991, 18, 565–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacLean, D.A.; Sinoway, L.I.; Leuenberger, U. Systemic hypoxia elevates skeletal muscle interstitial adenosine levels in humans. Circulation 1998, 98, 1990–1992. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Cao, H.; Qiu, X.; Xu, D.; Chen, Y.; Barnes, G.N.; Tu, Y.; Gyabaah, A.T.; Gharbal, A.H.A.A.; Peng, C.; et al. Neuroprotective Effects of Adenosine A1 Receptor Signaling on Cognitive Impairment Induced by Chronic Intermittent Hypoxia in Mice. Front Cell Neurosci. 2020, 14, 202. [Google Scholar] [CrossRef] [PubMed]
- West, J.B. High-altitude medicine. Lancet Respir. Med. 2015, 3, 12–13. [Google Scholar] [CrossRef]
- Richalet, J.P.; Larmignat, P.; Poitrine, E.; Letournel, M.; Canouï-Poitrine, F. Physiological risk factors for severe high-altitude illness: A prospective cohort study. Am. J. Respir. Crit. Care. Med. 2012, 185, 192–198. [Google Scholar] [CrossRef]
- Roach, R.C.; Maes, D.; Sandoval, D.; Robergs, R.A.; Icenogle, M.; Hinghofer-Szalkay, H.; Lium, D.; Loeppky, J.A. Exercise exacerbates acute mountain sickness at simulated high altitude. J. Appl. Physiol. 2000, 88, 581–585. [Google Scholar] [CrossRef]
- Hu, S.L.; Xiong, W.; Dai, Z.Q.; Zhao, H.L.; Feng, H. Cognitive Changes during Prolonged Stay at High Altitude and Its Correlation with C-Reactive Protein. PLoS ONE 2016, 11, e0146290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becher, B.; Spath, S.; Goverman, J. Cytokine networks in neuroinflammation. Nat. Rev. Immunol. 2017, 17, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.Z.; He, W.J.; Zhu, Z.R.; Xu, G.; Chen, D.W.; Gao, Y.Q. Adenosine A2A receptor involves in neuroinflammation-mediated cognitive decline through activating microglia under acute hypobaric hypoxia. Behav. Brain Res. 2018, 347, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Pedata, F.; Pugliese, A.M.; Coppi, E.; Dettori, I.; Maraula, G.; Cellai, L.; Melani, A. Adenosine A2A receptors modulate acute injury and neuroinflammation in brain ischemia. Mediat. Inflamm. 2014, 2014, 805198. [Google Scholar] [CrossRef] [Green Version]
- Dai, S.S.; Zhou, Y.G.; Li, W.; An, J.H.; Li, P.; Yang, N.; Chen, X.Y.; Xiong, R.P.; Liu, P.; Zhao, Y.; et al. Local glutamate level dictates adenosine A2A receptor regulation of neuroinflammation and traumatic brain injury. J. Neurosci. 2010, 30, 5802–5810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salys, J.; Kratzer, A.; Zamora, M.; Taraseviciene-Stewart, L. Hypoxia-mediated alterations in adenosine receptor expression in rat lung. Eur. Respir. J. 2011, 38, 333. [Google Scholar]
- Sharma Kandel, R.; Mishra, R.; Gautam, J.; Alaref, A.; Hassan, A.; Jahan, N. Patchy vasoconstriction versus inflammation: A debate in the pathogenesis of high altitude pulmonary edema. Cureus 2020, 12, e10371. [Google Scholar] [CrossRef]
- Mentzer, R.M., Jr.; Rubio, R.; Berne, R.M. Release of adenosine by hypoxic canine lung tissue and its possible role in pulmonary circulation. Am. J. Physiol. 1975, 229, 1625–1631. [Google Scholar] [CrossRef] [Green Version]
- Heinonen, I.; Kemppainen, J.; Kaskinoro, K.; Peltonen, J.E.; Sipilä, H.T.; Nuutila, P.; Knuuti, J.; Boushel, R.; Kalliokoski, K.K. Effects of adenosine, exercise, and moderate acute hypoxia on energy substrate utilization of human skeletal muscle. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 302, R385–R390. [Google Scholar] [CrossRef]
- Calbet, J.A.; Boushel, R.; Robach, P.; Hellsten, Y.; Saltin, B.; Lundby, C. Chronic hypoxia increases arterial blood pressure and reduces adenosine and ATP induced vasodilatation in skeletal muscle in healthy humans. Acta Physiol. 2014, 211, 574–584. [Google Scholar] [CrossRef]
- Waring, W.; Thomson, A.; Adwani, S.; Rosseel, A.; Potter, J.; Webb, D.; Maxwell, S. Cardiovascular effects of acute oxygen administration in healthy adults. J. Cardiovasc. Pharmacol. 2003, 42, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Molenat, F.; Boussuges, A.; Grandfond, A.; Rostain, J.C.; Sainty, J.M.; Robinet, C.; Galland, F.; Meliet, J.L. Modifications of cardiovascular function secondary to hyperbaric hyperoxia in healthy volunteers: An Echocardiographic and Doppler study. Clin. Sci. 2004, 106, 389–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gole, Y.; Gargne, O.; Coulange, M.; Steinberg, J.G.; Bouhaddi, M.; Jammes, Y.; Regnard, J.; Boussuges, A. Hyperoxia-induced alterations in cardiovascular function and autonomic control during return to normoxic breathing. Eur. J. Appl. Physiol. 2011, 111, 937–946. [Google Scholar] [CrossRef] [PubMed]
- Harten, J.M.; Anderson, K.J.; Angerson, W.J.; Booth, M.G.; Kinsella, J. The effect of normobaric hyperoxia on cardiac index in healthy awake volunteers. Anaesthesia 2003, 58, 885–888. [Google Scholar] [CrossRef]
- Rossi, P.; Boussuges, A. Hyperoxia-induced arterial compliance decrease in healthy man. Clin. Physiol. Funct. Imag. 2005, 25, 10–15. [Google Scholar] [CrossRef]
- Rubanyi, G.M.; Vanhoutte, P.M. Superoxide anions and hyperoxia inactivate endothelium derived relaxing factor. Am. J. Physiol. 1986, 250, H822–H827. [Google Scholar] [CrossRef]
- Welsh, D.G.; Jackson, W.F.; Segal, S.S. Oxygen induces electromechanical coupling in arteriolar smooth muscle cells: A role for L-type Ca2+ channels. Am. J. Physiol. 1998, 274, H2018–H2024. [Google Scholar] [CrossRef]
- Boussuges, A.; Rives, S.; Marlinge, M.; Chaumet, G.; Vallée, N.; Guieu, R.; Gavarry, O. Hyperoxia during exercise: Impact on adenosine plasma levels and hemodynamic data. Front. Physiol. 2020, 11, 97. [Google Scholar] [CrossRef] [Green Version]
- Sarelius, I.; Pohl, U. Control of muscle blood flow during exercise: Local factors and integrative mechanisms. Acta Physiol. 2010, 199, 349–365. [Google Scholar] [CrossRef]
- Marshall, J.M. The roles of adenosine and related substances in exercise hyperaemia. J. Physiol. 2007, 583, 835–845. [Google Scholar] [CrossRef]
- Scott, A.; Fruttiger, M. Oxygen-induced retinopathy: A model for vascular pathology in the retina. Eye 2010, 24, 416–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Li, H.; Li, B.; Zhong, D.; Gu, X.; Tang, L.; Wang, Y.; Wang, C.; Zhou, R.; Li, Y.; et al. Adenosine A1 Receptors Selectively Modulate Oxygen-Induced Retinopathy at the Hyperoxic and Hypoxic Phases by Distinct Cellular Mechanisms. Investig. Ophthalmol Vis. Sci. 2015, 56, 8108–8119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ottolenghi, S.; Sabbatini, G.; Brizzolari, A.; Samaja, M.; Chiumello, D. Hyperoxia and oxidative stress in anesthesia and critical care medicine. Minerva Anestesiol. 2020, 86, 64–75. [Google Scholar] [CrossRef] [PubMed]
- Arieli, R.; Arieli, Y.; Daskalovic, Y.; Eynan, M.; Abramovich, A. CNS oxygen toxicity in closed-circuit diving: Signs and symptoms before loss of consciousness. Aviat. Space Environ. Med. 2006, 77, 1153–1157. [Google Scholar] [PubMed]
- Nayeem, M.A.; Poloyac, S.M.; Falck, J.R.; Zeldin, D.C.; Ledent, C.; Ponnoth, D.S.; Ansari, H.R.; Mustafa, S.J. Role of CYP epoxygenases in A2A AR-mediated relaxation using A2A AR-null and wild-type mice. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H2068–H2078. [Google Scholar] [CrossRef] [Green Version]
- Barry, W.A.; Demchenko, I.T.; Piantadosi, C.A. Two faces of nitric oxide, implications for cellular mechanisms of oxygen toxicity. J. Appl. Physiol. 2009, 106, 662–667. [Google Scholar]
- Demchenko, I.T.; Boso, A.E.; Whorton, A.R.; Piantadosi, C.A. Nitric oxide production is enhanced in rat brain before oxygen-induced convulsions. Brain Res. 2001, 917, 253–261. [Google Scholar] [CrossRef]
- Sato, T.; Takeda, Y.; Hagioka, S.; Zhang, S.; Hirakawa, M. Changes in nitric oxide production and cerebral blood flow before development of hyperbaric oxygen-induced seizures in rats. Brain Res. 2001, 918, 131–140. [Google Scholar] [CrossRef] [Green Version]
- Jones, M.W.; Brett, K.; Han, N.; Wyatt, H.A. Hyperbaric Physics; StatPearls Publishing: Treasure Island, FL, USA, 2017. [Google Scholar]
- You, K.; Xu, X.; Fu, J.; Xu, S.; Yue, X.; Yu, Z.; Xue, X. Hyperoxia disrupts pulmonary epithelial barrier in newborn rats via the deterioration of occludin and ZO-1. Respir. Res. 2012, 13, 36. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Xue, X.; You, K.; Fu, J. Caveolin-1 regulates the expression of tight junction proteins during hyperoxia-induced pulmonary epithelial barrier breakdown. Respir. Res. 2016, 17, 50. [Google Scholar] [CrossRef] [Green Version]
- Maniatis, N.A.; Chernaya, O.; Shinin, V.; Minshall, R.D. Caveolins and lung function. Adv. Exp. Med. Biol. 2012, 729, 157–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volmer, J.B.; Thompson, L.F.; Blackburn, M.R. Ecto-50 -nucleotidase (CD73)-mediated adenosine production is tissue protective in a model of bleomycin-induced lung injury. J. Immunol. 2006, 176, 4449–4458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eckle, T.; Fullbier, L.; Wehrmann, M.; Khoury, J.; Mittelbronn, M.; Ibla, J.; Rosenberger, P.; Eltzschig, H.K. Identification of ectonucleotidases CD39 and CD73 in innate protection during acute lung injury. J. Immunol. 2007, 178, 8127–8137. [Google Scholar] [CrossRef] [PubMed]
- Davies, J.; Karmouty-Quintana, H.; Le, T.T.; Chen, N.Y.; Weng, T.; Luo, F.; Molina, J.; Moorthy, B.; Blackburn, M.R. Adenosine promotes vascular barrier function in hyperoxic lung injury. Physiol. Rep. 2014, 2, e12155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schingnitz, U.; Hartmann, K.; Macmanus, C.F.; Eckle, T.; Zug, S.; Colgan, S.P.; Eltzschig, H.K. Signaling through the A2B adenosine receptor dampens endotoxin-induced acute lung injury. J. Immunol. 2010, 184, 5271–5279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thiel, M.; Chouker, A.; Ohta, A.; Jackson, E.; Caldwell, C.; Smith, P.; Lukashev, D.; Bittmann, I.; Sitkovsky, M.V. Oxygenation inhibits the physiological tissue-protecting mechanism and thereby exacerbates acute inflammatory lung injury. PLoS Biol. 2005, 3, e174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boussuges, A.; Bourenne, J.; Eloufir, F.; Fromonot, J.; Mottola, G.; Risso, J.J.; Vallee, N.; Bregeon, F.; Guieu, R. Contribution of Adenosine in the Physiological Changes and Injuries Secondary to Exposure to Extreme Oxygen Pressure in Healthy Subjects. Biomedicines 2022, 10, 2059. https://doi.org/10.3390/biomedicines10092059
Boussuges A, Bourenne J, Eloufir F, Fromonot J, Mottola G, Risso JJ, Vallee N, Bregeon F, Guieu R. Contribution of Adenosine in the Physiological Changes and Injuries Secondary to Exposure to Extreme Oxygen Pressure in Healthy Subjects. Biomedicines. 2022; 10(9):2059. https://doi.org/10.3390/biomedicines10092059
Chicago/Turabian StyleBoussuges, Alain, Jeremy Bourenne, Farid Eloufir, Julien Fromonot, Giovanna Mottola, Jean Jacques Risso, Nicolas Vallee, Fabienne Bregeon, and Régis Guieu. 2022. "Contribution of Adenosine in the Physiological Changes and Injuries Secondary to Exposure to Extreme Oxygen Pressure in Healthy Subjects" Biomedicines 10, no. 9: 2059. https://doi.org/10.3390/biomedicines10092059
APA StyleBoussuges, A., Bourenne, J., Eloufir, F., Fromonot, J., Mottola, G., Risso, J. J., Vallee, N., Bregeon, F., & Guieu, R. (2022). Contribution of Adenosine in the Physiological Changes and Injuries Secondary to Exposure to Extreme Oxygen Pressure in Healthy Subjects. Biomedicines, 10(9), 2059. https://doi.org/10.3390/biomedicines10092059