
Genetic Susceptibility to Insulin Resistance and Its Association with Estimated Longevity in the Hungarian General and Roma Populations




Abstract
:1. Introduction
2. Materials and Methods
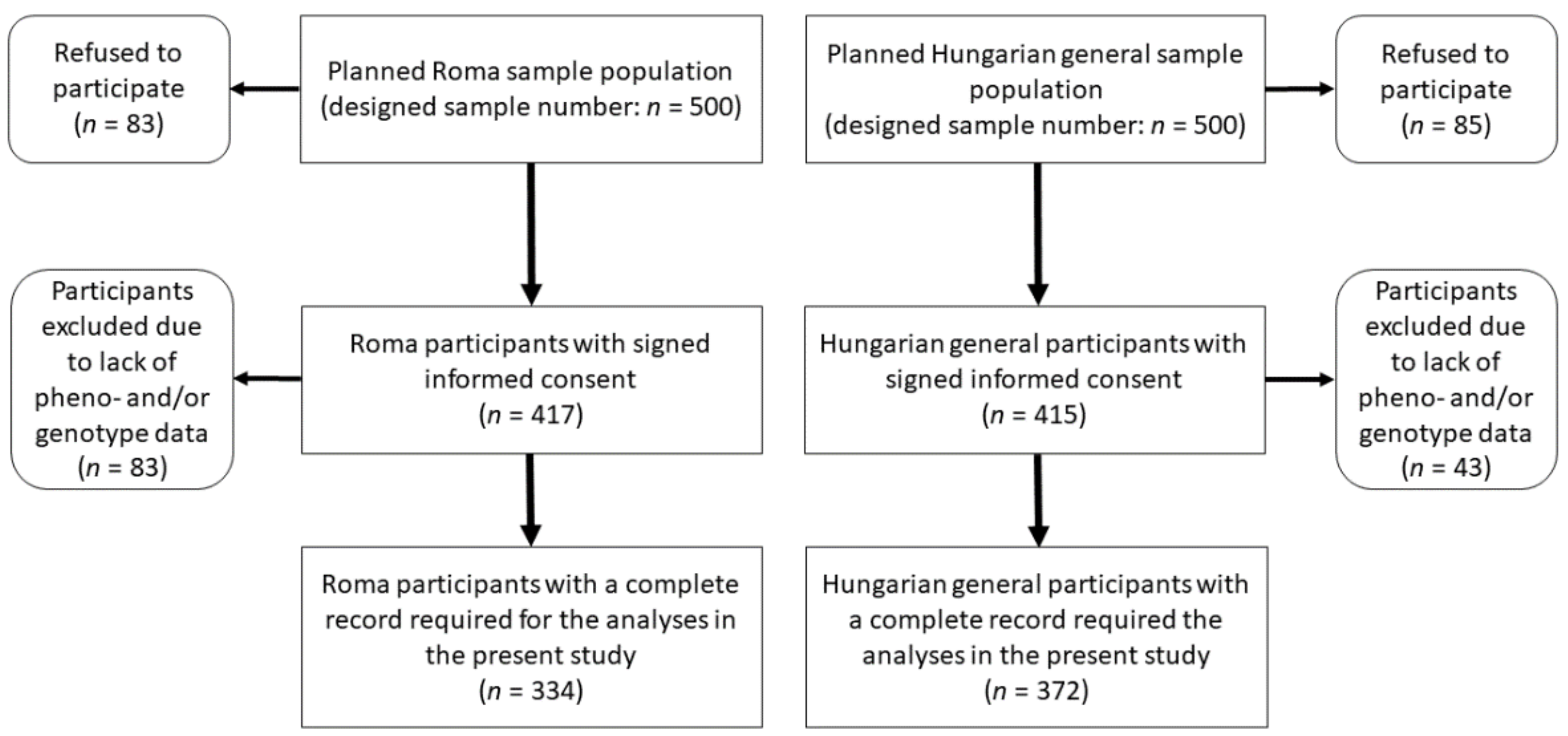
2.1. Sample Populations
2.2. Data Used to Identify Insulin Resistance in Sample Populations
2.3. DNA Extraction, SNP Selection, Genotyping, Testing Hardy-Weinberg Equilibrium, and Linkage Disequilibrium
2.4. Identification and Coding of the Genetic Model Best Associated with HOMA—IR for SNPs
2.5. Calculation and Optimization of the Genetic Risk Score
2.6. Statistical Analysis
2.7. Ethical Statement
3. Results
3.1. Characteristics of the Study Populations and Results of the HWE and LD Analyses
3.2. Results Obtained from the Analysis of the Determination of the Best Fitting Genetic Model for SNPs
3.3. Results of the Calculation and Optimization of the GRS Model and Determination of the Bonferroni Corrected p Value
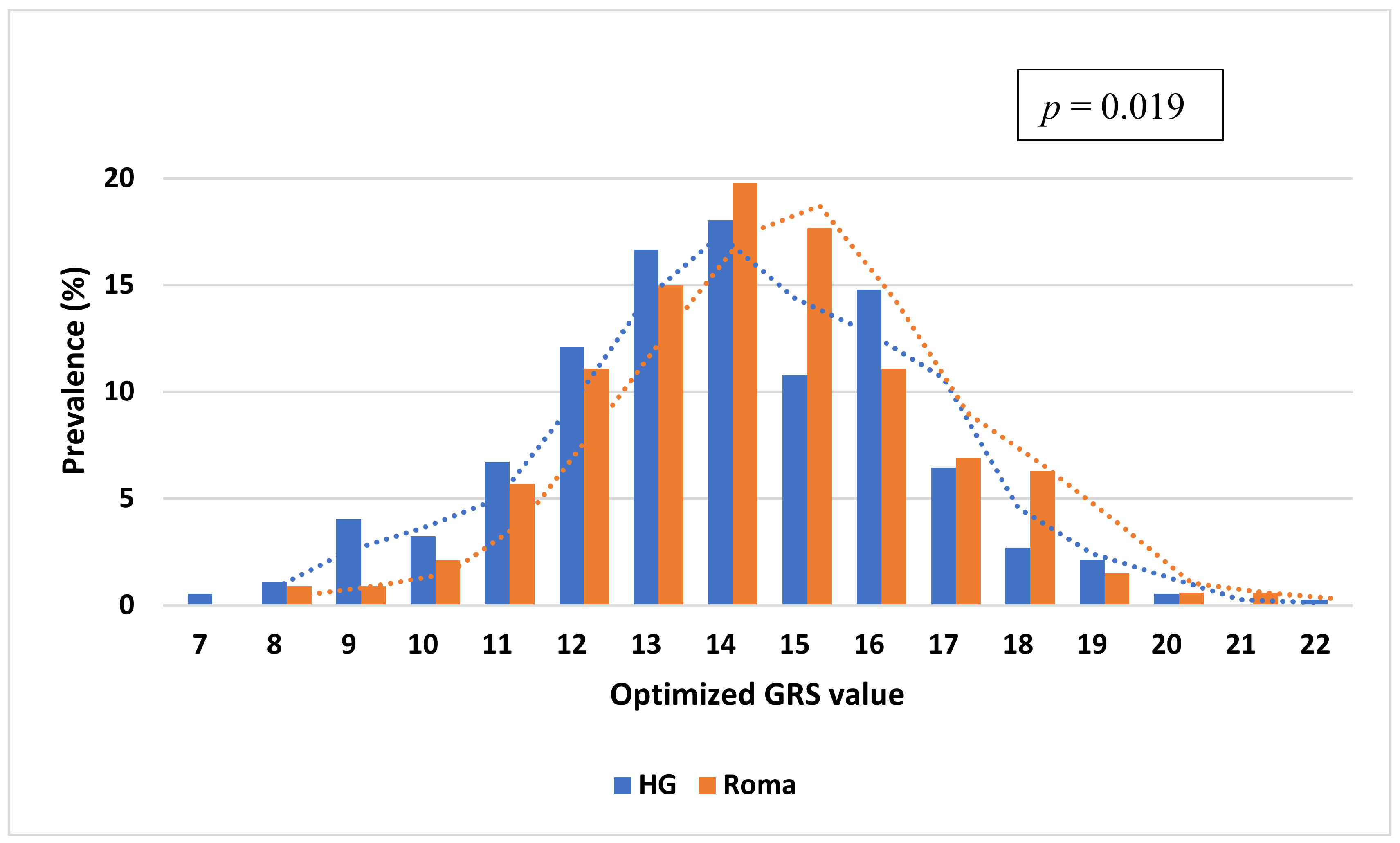
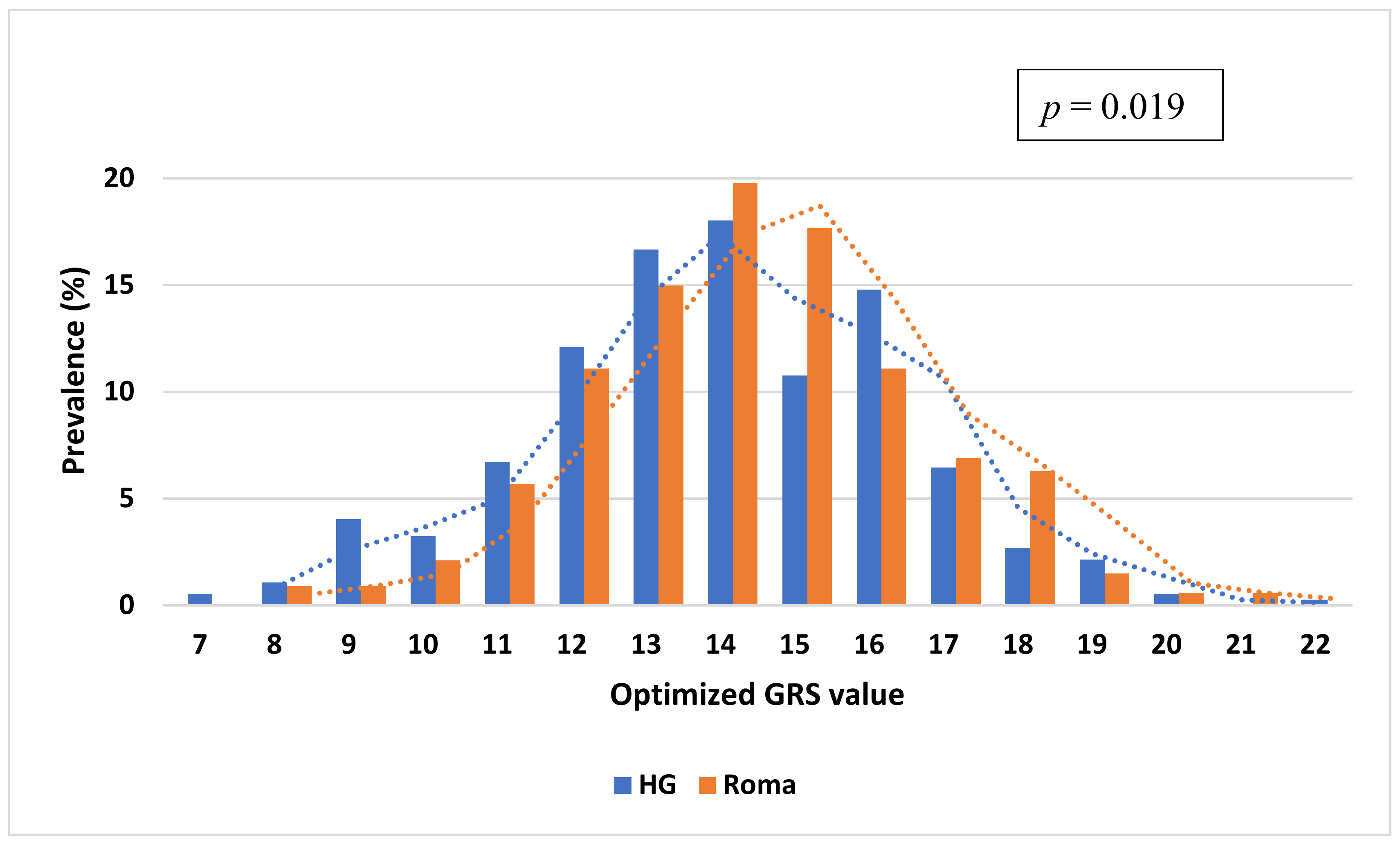
3.4. Comparison of the Distribution of oGRS Value and Its Association with Fasting Insulin, Fasting Glucose Levels, and HOMA—IR in the Study Populations
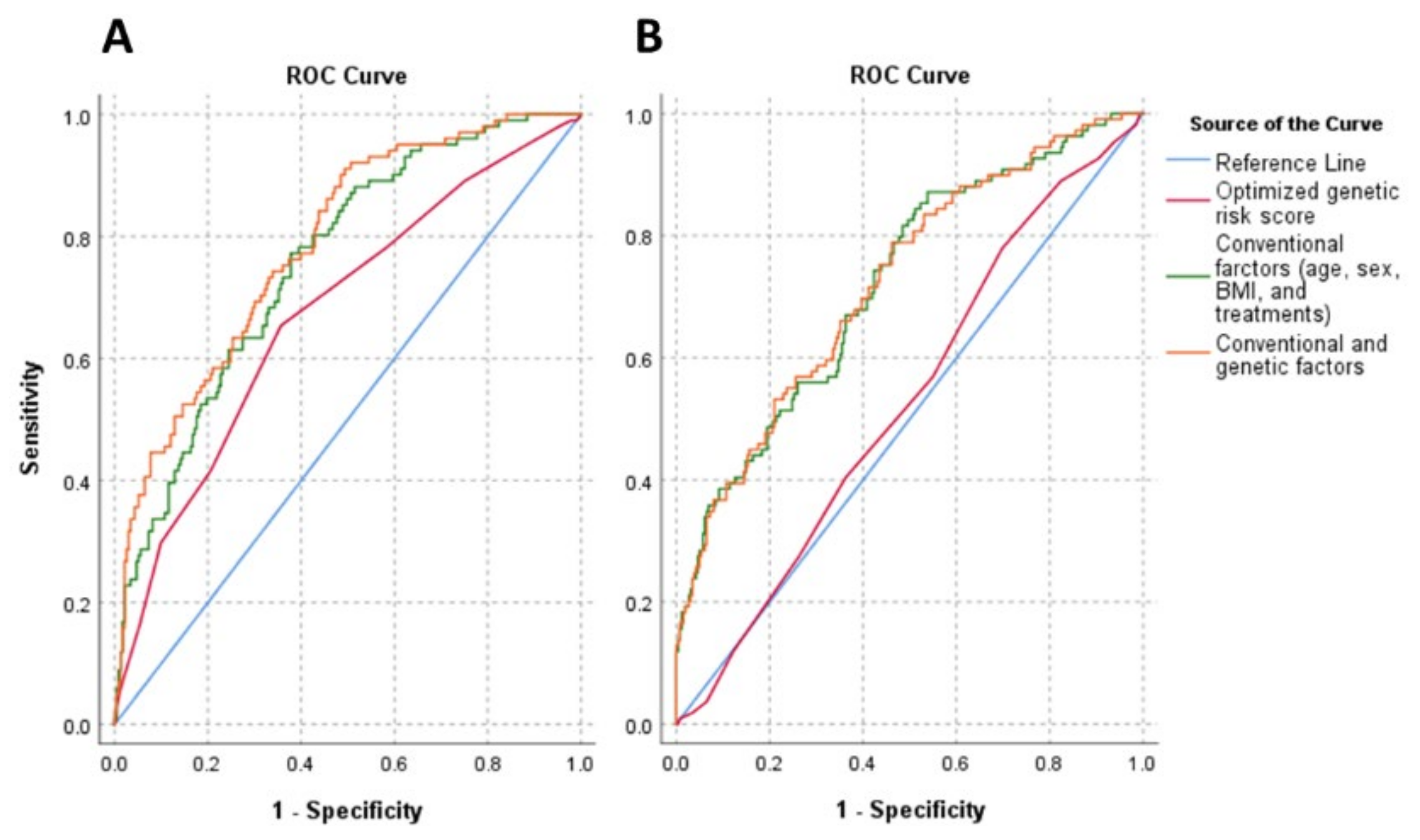
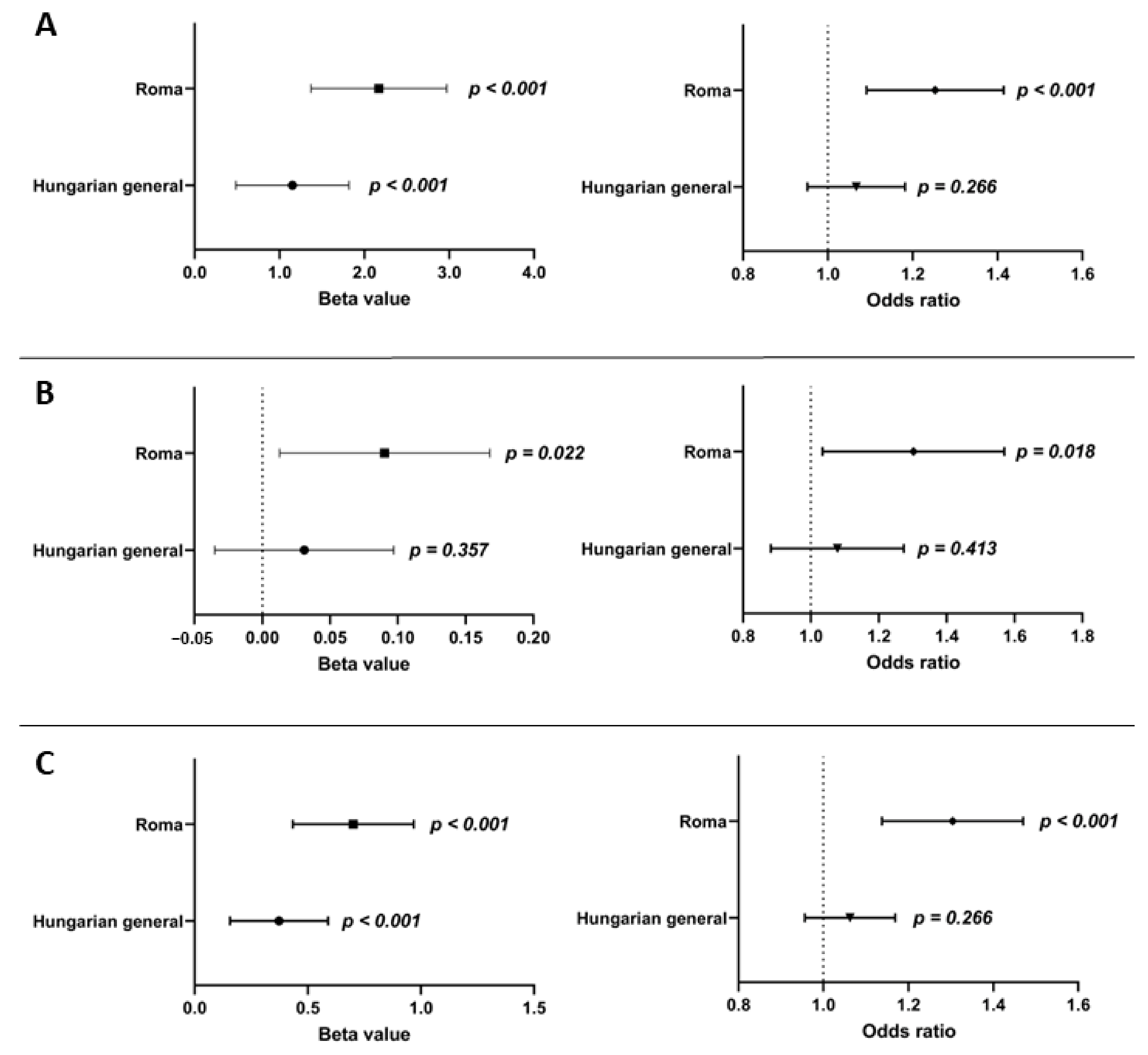
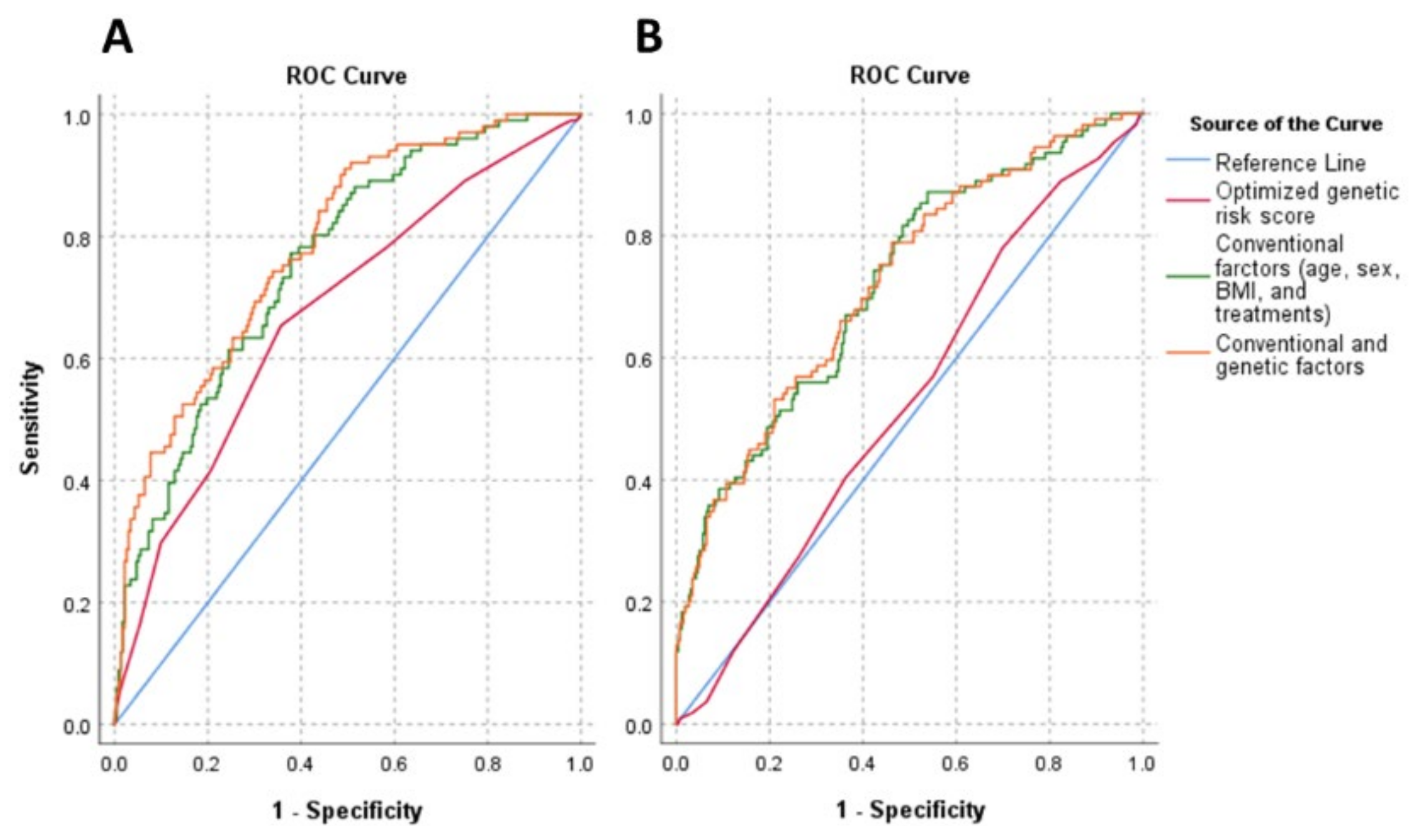
3.5. The Effect of Genetic (oGRS) and Conventional (Age, Gender, BMI, and Treatments) Risk Factors and the Combination of Them on Insulin Resistance in the Study Populations
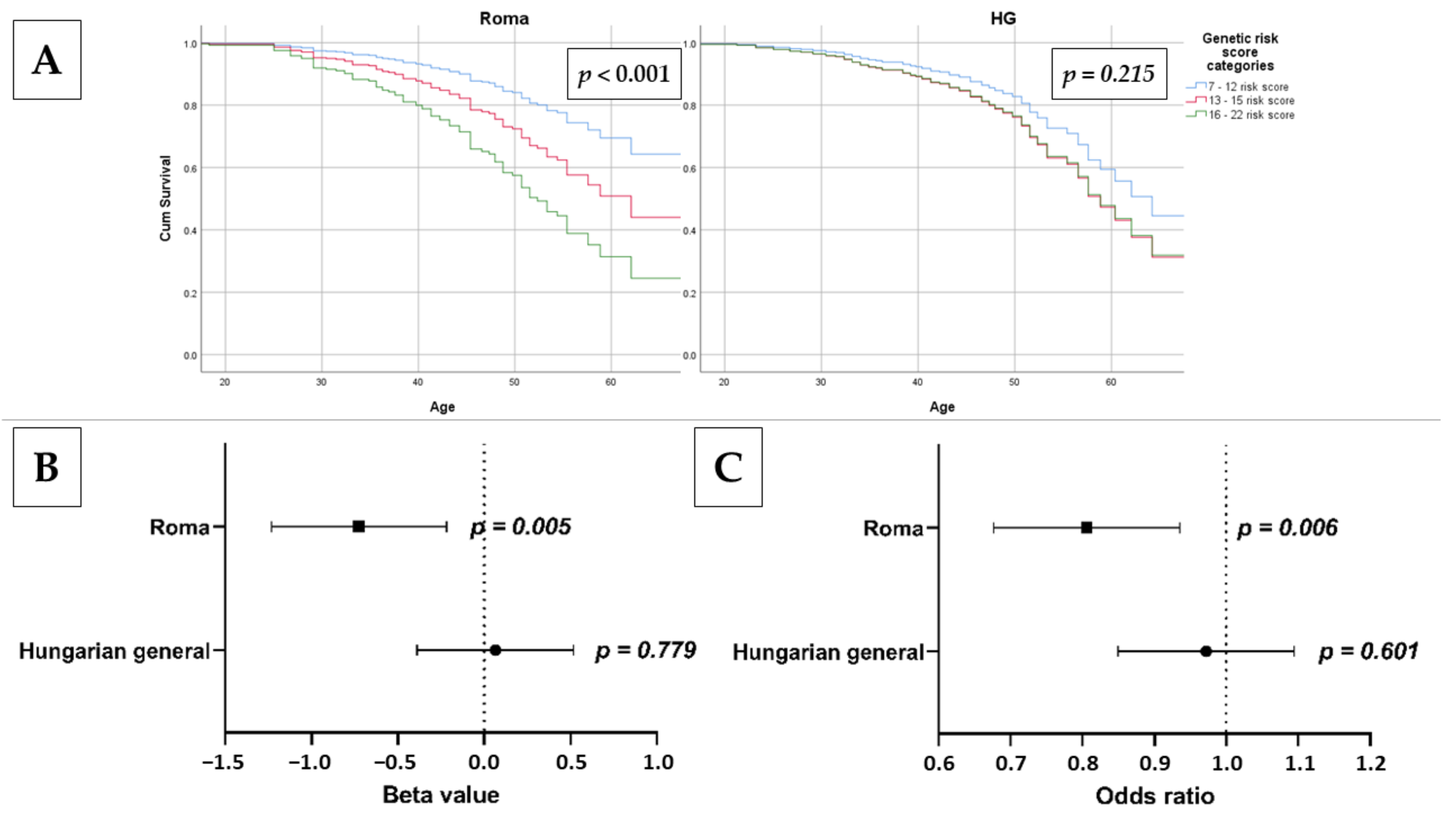
3.6. Association of Genetic Risk Groups with Insulin Resistance in the Study Populations
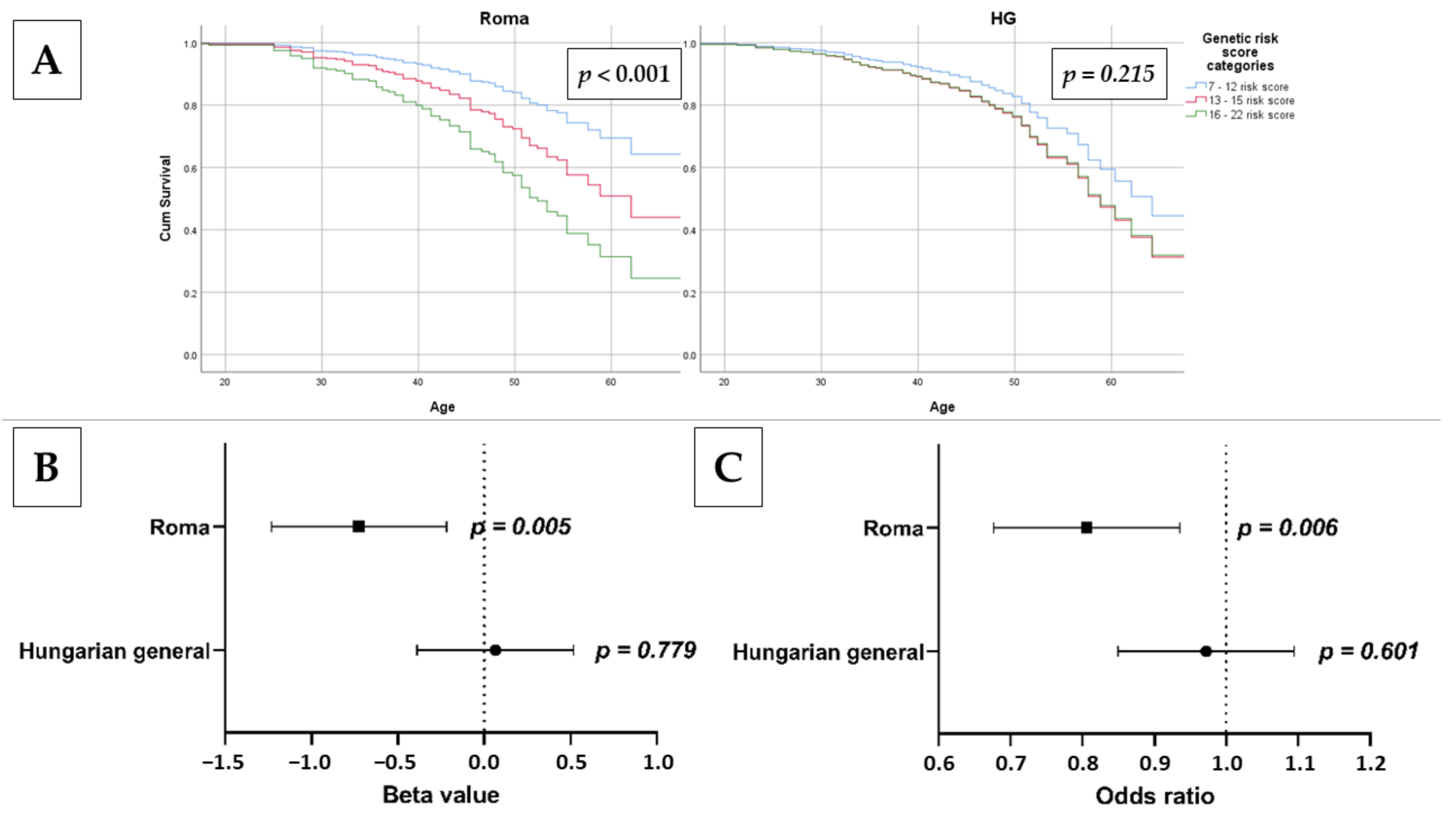
3.7. Association of oGRS with Early Onset of Insulin Resistance and Estimated Longevity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- International Diabetes Federation. IDF Diabetes Atlas 2021. Available online: https://diabetesatlas.org/idfawp/resource-files/2021/07/IDF_Atlas_10th_Edition_2021.pdf (accessed on 9 March 2022).
- Murray, C.J.L.; Aravkin, A.Y.; Zheng, P.; Abbafati, C.; Abbas, K.M.; Abbasi-Kangevari, M.; Abd-Allah, F.; Abdelalim, A.; Abdollahi, M.; Abdollahpour, I.; et al. Global burden of 87 risk factors in 204 countries and territories, 1990–2019: A systematic analysis for the Global Burden of Disease Study 2019. Lancet 2020, 396, 1223–1249. [Google Scholar] [CrossRef]
- Saeedi, P.; Petersohn, I.; Salpea, P.; Malanda, B.; Karuranga, S.; Unwin, N.; Colagiuri, S.; Guariguata, L.; Motala, A.A.; Ogurtsova, K.; et al. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas, 9th edition. Diabetes Res. Clin. Pr. 2019, 157, 107843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, X.; Xu, Y.; Pan, X.; Xu, J.; Ding, Y.; Sun, X.; Song, X.; Ren, Y.; Shan, P.F. Global, regional, and national burden and trend of diabetes in 195 countries and territories: An analysis from 1990 to 2025. Sci. Rep. 2020, 10, 14790. [Google Scholar] [CrossRef] [PubMed]
- Karachaliou, F.; Simatos, G.; Simatou, A. The Challenges in the Development of Diabetes Prevention and Care Models in Low-Income Settings. Front. Endocrinol. 2020, 11, 518. [Google Scholar] [CrossRef]
- Global Health Estimates: Deaths by Cause, Age, Sex and Country, 2000–2012; World Health Organization: Geneva, Switzerland, 2015.
- Xu, Z.Y.; Yu, D.; Yin, X.Y.; Zheng, F.P.; Li, H. Socioeconomic status is associated with global diabetes prevalence. Oncotarget 2017, 8, 44434–44439. [Google Scholar] [CrossRef] [Green Version]
- Pham, T.M.; Carpenter, J.R.; Morris, T.P.; Sharma, M.; Petersen, I. Ethnic Differences in the Prevalence of Type 2 Diabetes Diagnoses in the UK: Cross-Sectional Analysis of the Health Improvement Network Primary Care Database. Clin. Epidemiol 2019, 11, 1081–1088. [Google Scholar] [CrossRef] [Green Version]
- Wondmkun, Y.T. Obesity, Insulin Resistance, and Type 2 Diabetes: Associations and Therapeutic Implications. Diabetes Metab. Syndr. Obes. Targets Ther. 2020, 13, 3611–3616. [Google Scholar] [CrossRef]
- Czech, M.P. Insulin action and resistance in obesity and type 2 diabetes. Nat. Med. 2017, 23, 804–814. [Google Scholar] [CrossRef]
- Ormazabal, V.; Nair, S.; Elfeky, O.; Aguayo, C.; Salomon, C.; Zuniga, F.A. Association between insulin resistance and the development of cardiovascular disease. Cardiovasc. Diabetol. 2018, 17, 122. [Google Scholar] [CrossRef]
- Paneni, F.; Costantino, S.; Cosentino, F. Insulin resistance, diabetes, and cardiovascular risk. Curr. Atheroscler. Rep. 2014, 16, 419. [Google Scholar] [CrossRef]
- Paolisso, G.; Barbieri, M.; Rizzo, M.R.; Carella, C.; Rotondi, M.; Bonafè, M.; Franceschi, C.; Rose, G.; De Benedictis, G. Low insulin resistance and preserved β-cell function contribute to human longevity but are not associated with TH-INS genes. Exp. Gerontol. 2001, 37, 149–156. [Google Scholar] [CrossRef]
- Wijsman, C.A.; Rozing, M.P.; Streefland, T.C.; le Cessie, S.; Mooijaart, S.P.; Slagboom, P.E.; Westendorp, R.G.; Pijl, H.; van Heemst, D.; Leiden Longevity Study Group. Familial longevity is marked by enhanced insulin sensitivity. Aging Cell 2011, 10, 114–121. [Google Scholar] [CrossRef]
- Avogaro, A.; de Kreutzenberg, S.V.; Fadini, G.P. Insulin signaling and life span. Pflug. Arch. 2010, 459, 301–314. [Google Scholar] [CrossRef] [PubMed]
- Hectors, T.L.M.; Vanparys, C.; Van Gaal, L.F.; Jorens, P.G.; Covaci, A.; Blust, R. Insulin Resistance and Environmental Pollutants: Experimental Evidence and Future Perspectives. Environ. Health Persp. 2013, 121, 1273–1281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Y.; Jiang, C.D.; Chang, A.M.; Shi, Y.; Gao, J.J.; Zhu, L.L.; Zhang, Z. Interactions among insulin resistance, inflammation factors, obesity-related gene polymorphisms, environmental risk factors, and diet in the development of gestational diabetes mellitus. J. Matern.-Fetal Neonatal Med. 2019, 32, 339–347. [Google Scholar] [CrossRef]
- Kyrou, I.; Tsigos, C.; Mavrogianni, C.; Cardon, G.; Van Stappen, V.; Latomme, J.; Kivela, J.; Wikstrom, K.; Tsochev, K.; Nanasi, A.; et al. Sociodemographic and lifestyle-related risk factors for identifying vulnerable groups for type 2 diabetes: A narrative review with emphasis on data from Europe. BMC Endocr. Disord. 2020, 20, 134. [Google Scholar] [CrossRef] [Green Version]
- Gratas-Delamarche, A.; Derbre, F.; Vincent, S.; Cillard, J. Physical inactivity, insulin resistance, and the oxidative-inflammatory loop. Free Radic. Res. 2014, 48, 93–108. [Google Scholar] [CrossRef]
- Roberts, C.K.; Hevener, A.L.; Barnard, R.J. Metabolic syndrome and insulin resistance: Underlying causes and modification by exercise training. Compr. Physiol. 2013, 3, 1–58. [Google Scholar] [CrossRef] [Green Version]
- Fedewa, M.V.; Gist, N.H.; Evans, E.M.; Dishman, R.K. Exercise and Insulin Resistance in Youth: A Meta-Analysis. Pediatrics 2014, 133, E163–E174. [Google Scholar] [CrossRef] [Green Version]
- Kavadar, G.; Demircioglu, D.T.; Ozgonenel, L.; Emre, T.Y. The relationship between vitamin D status, physical activity and insulin resistance in overweight and obese subjects. Bosnian J. Basic Med. 2015, 15, 62–66. [Google Scholar] [CrossRef] [Green Version]
- Shou, J.; Chen, P.J.; Xiao, W.H. Mechanism of increased risk of insulin resistance in aging skeletal muscle. Diabetol. Metab. Syndr. 2020, 12, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piko, P.; Werissa, N.A.; Fiatal, S.; Sandor, J.; Adany, R. Impact of Genetic Factors on the Age of Onset for Type 2 Diabetes Mellitus in Addition to the Conventional Risk Factors. J. Pers. Med. 2020, 11, 6. [Google Scholar] [CrossRef] [PubMed]
- Malone, J.I.; Hansen, B.C. Does obesity cause type 2 diabetes mellitus (T2DM)? Or is it the opposite? Pediatr. Diabetes 2019, 20, 5–9. [Google Scholar] [CrossRef] [Green Version]
- Leitner, D.R.; Fruhbeck, G.; Yumuk, V.; Schindler, K.; Micic, D.; Woodward, E.; Toplak, H. Obesity and Type 2 Diabetes: Two Diseases with a Need for Combined Treatment Strategies—EASO Can Lead the Way. Obes. Facts 2017, 10, 483–492. [Google Scholar] [CrossRef]
- Artese, A.; Stamford, B.A.; Moffatt, R.J. Cigarette Smoking: An Accessory to the Development of Insulin Resistance. Am. J. Lifestyle Med. 2019, 13, 602–605. [Google Scholar] [CrossRef] [PubMed]
- Maddatu, J.; Anderson-Baucum, E.; Evans-Molina, C. Smoking and the risk of type 2 diabetes. Transl. Res. 2017, 184, 101–107. [Google Scholar] [CrossRef]
- Castro-Quezada, I.; Flores-Guillen, E.; Nunez-Ortega, P.E.; Irecta-Najera, C.A.; Sanchez-Chino, X.M.; Mendez-Flores, O.G.; Olivo-Vidal, Z.E.; Garcia-Miranda, R.; Solis-Hernandez, R.; Ochoa-Diaz-Lopez, H. Dietary Carbohydrates and Insulin Resistance in Adolescents from Marginalized Areas of Chiapas, Mexico. Nutrients 2019, 11, 3066. [Google Scholar] [CrossRef] [Green Version]
- Hojlund, K. Metabolism and insulin signaling in common metabolic disorders and inherited insulin resistance. Dan. Med. J. 2014, 61, B4890. [Google Scholar]
- Brown, A.E.; Walker, M. Genetics of Insulin Resistance and the Metabolic Syndrome. Curr. Cardiol. Rep. 2016, 18, 75. [Google Scholar] [CrossRef] [Green Version]
- Kodama, K.; Tojjar, D.; Yamada, S.; Toda, K.; Patel, C.J.; Butte, A.J. Ethnic differences in the relationship between insulin sensitivity and insulin response: A systematic review and meta-analysis. Diabetes Care 2013, 36, 1789–1796. [Google Scholar] [CrossRef] [Green Version]
- Lotta, L.A.; Gulati, P.; Day, F.R.; Payne, F.; Ongen, H.; Van De Bunt, M.; Gaulton, K.J.; Eicher, J.D.; Sharp, S.J.; Rolfe, E.D.L. Integrative genomic analysis implicates limited peripheral adipose storage capacity in the pathogenesis of human insulin resistance. Nat. Genet. 2017, 49, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Manning, A.K.; Hivert, M.F.; Scott, R.A.; Grimsby, J.L.; Bouatia-Naji, N.; Chen, H.; Rybin, D.; Liu, C.T.; Bielak, L.F.; Prokopenko, I.; et al. A genome-wide approach accounting for body mass index identifies genetic variants influencing fasting glycemic traits and insulin resistance. Nat. Genet. 2012, 44, 659–669. [Google Scholar] [CrossRef] [PubMed]
- Stancakova, A.; Kuulasmaa, T.; Kuusisto, J.; Mohlke, K.L.; Collins, F.S.; Boehnke, M.; Laakso, M. Genetic risk scores in the prediction of plasma glucose, impaired insulin secretion, insulin resistance and incident type 2 diabetes in the METSIM study. Diabetologia 2017, 60, 1722–1730. [Google Scholar] [CrossRef] [PubMed]
- EU Roma Strategic Framework for Equality, Inclusion and Participation for 2020–2030; European Commission, European Committee of the Regions: Brussels, Belgium, 2020.
- Melegh, B.I.; Banfai, Z.; Hadzsiev, K.; Miseta, A.; Melegh, B. Refining the South Asian Origin of the Romani people. BMC Genet. 2017, 18, 82. [Google Scholar] [CrossRef] [Green Version]
- Sudzinova, A.; Nagyova, I.; Studencan, M.; Rosenberger, J.; Skodova, Z.; Vargova, H.; Middel, B.; Reijneveld, S.A.; van Dijk, J.P. Roma coronary heart disease patients have more medical risk factors and greater severity of coronary heart disease than non-Roma. Int. J. Public Health 2013, 58, 409–415. [Google Scholar] [CrossRef] [Green Version]
- Kosa, Z.; Moravcsik-Kornyicki, A.; Dioszegi, J.; Roberts, B.; Szabo, Z.; Sandor, J.; Adany, R. Prevalence of metabolic syndrome among Roma: A comparative health examination survey in Hungary. Eur. J. Public Health 2015, 25, 299–304. [Google Scholar] [CrossRef] [Green Version]
- Adany, R. Roma health is global ill health. Eur. J. Public Health 2014, 24, 702–703. [Google Scholar] [CrossRef] [Green Version]
- Tudor, M. Closing the Life Expectancy Gap of Roma in Europe; European Public Health Alliance: Brussels, Belgium, 2019. [Google Scholar]
- Piko, P.; Fiatal, S.; Kosa, Z.; Sandor, J.; Adany, R. Genetic factors exist behind the high prevalence of reduced high-density lipoprotein cholesterol levels in the Roma population. Atherosclerosis 2017, 263, 119–126. [Google Scholar] [CrossRef]
- Piko, P.; Fiatal, S.; Kosa, Z.; Sandor, J.; Adany, R. Data to genetic risk assessment on high-density cholesterol level associated polymorphisms in Hungarian general and Roma populations. Data Brief 2017, 14, 354–359. [Google Scholar] [CrossRef]
- Nagy, K.; Fiatal, S.; Sandor, J.; Adany, R. Distinct Penetrance of Obesity-Associated Susceptibility Alleles in the Hungarian General and Roma Populations. Obes. Facts 2017, 10, 444–457. [Google Scholar] [CrossRef] [Green Version]
- Ádány, R.; Pikó, P.; Fiatal, S.; Kósa, Z.; Sándor, J.; Bíró, É.; Kósa, K.; Paragh, G.; Bácsné Bába, É.; Veres-Balajti, I. Prevalence of insulin resistance in the Hungarian general and Roma populations as defined by using data generated in a complex health (interview and examination) survey. Int. J. Environ. Res. Public Health 2020, 17, 4833. [Google Scholar] [CrossRef] [PubMed]
- Werissa, N.A.; Piko, P.; Fiatal, S.; Kosa, Z.; Sandor, J.; Adany, R. SNP-Based Genetic Risk Score Modeling Suggests No Increased Genetic Susceptibility of the Roma Population to Type 2 Diabetes Mellitus. Genes 2019, 10, 942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziaee, A.; Esmailzadehha, N.; Oveisi, S.; Ghorbani, A.; Ghanei, L. The threshold value of homeostasis model assessment for insulin resistance in Qazvin Metabolic Diseases Study (QMDS): Assessment of metabolic syndrome. JRHS J. Res. Health Sci. 2015, 15, 94–100. [Google Scholar]
- Friedrich, N.; Thuesen, B.; Jorgensen, T.; Juul, A.; Spielhagen, C.; Wallaschofksi, H.; Linneberg, A. The association between IGF-I and insulin resistance: A general population study in Danish adults. Diabetes Care 2012, 35, 768–773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeFronzo, R.A.; Tobin, J.D.; Andres, R. Glucose clamp technique: A method for quantifying insulin secretion and resistance. Am. J. Physiol. 1979, 237, E214–E223. [Google Scholar] [CrossRef]
- Sanchez-Garcia, A.; Rodriguez-Gutierrez, R.; Mancillas-Adame, L.; Gonzalez-Nava, V.; Diaz Gonzalez-Colmenero, A.; Solis, R.C.; Alvarez-Villalobos, N.A.; Gonzalez-Gonzalez, J.G. Diagnostic Accuracy of the Triglyceride and Glucose Index for Insulin Resistance: A Systematic Review. Int. J. Endocrinol. 2020, 2020, 4678526. [Google Scholar] [CrossRef]
- Horakova, D.; Stepanek, L.; Janout, V.; Janoutova, J.; Pastucha, D.; Kollarova, H.; Petrakova, A.; Stepanek, L.; Husar, R.; Martinik, K. Optimal Homeostasis Model Assessment of Insulin Resistance (HOMA-IR) Cut-Offs: A Cross-Sectional Study in the Czech Population. Medicina 2019, 55, 158. [Google Scholar] [CrossRef] [Green Version]
- Patarrão, R.S.; Lautt, W.W.; Macedo, M.P. Assessment of methods and indexes of insulin sensitivity. Rev. Port. De Endocrinol. Diabetes E Metab. 2014, 9, 65–73. [Google Scholar] [CrossRef] [Green Version]
- Singh, Y.; Garg, M.K.; Tandon, N.; Marwaha, R.K. A Study of Insulin Resistance by HOMA-IR and its Cut-off Value to Identify Metabolic Syndrome in Urban Indian Adolescents. J. Clin. Res. Pediatr. Endocrinol. 2013, 5, 245–251. [Google Scholar] [CrossRef]
- Tang, Q.; Li, X.; Song, P.; Xu, L. Optimal cut-off values for the homeostasis model assessment of insulin resistance (HOMA-IR) and pre-diabetes screening: Developments in research and prospects for the future. Drug Discov. Ther. 2015, 9, 380–385. [Google Scholar] [CrossRef] [Green Version]
- Salanti, G.; Southam, L.; Altshuler, D.; Ardlie, K.; Barroso, I.; Boehnke, M.; Cornelis, M.C.; Frayling, T.M.; Grallert, H.; Grarup, N.; et al. Underlying genetic models of inheritance in established type 2 diabetes associations. Am. J. Epidemiol. 2009, 170, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Templeton, G.F. A two-step approach for transforming continuous variables to normal: Implications and recommendations for IS research. Commun. Assoc. Inf. Syst. 2011, 28, 4. [Google Scholar] [CrossRef] [Green Version]
- Chopra, A.K. Metabolic syndrome or insulin resistance: Evolution, controversies and association with cardiovascular disease risk. Indian J. Clin. Cardiol. 2020, 1, 77–85. [Google Scholar] [CrossRef]
- Saklayen, M.G. The Global Epidemic of the Metabolic Syndrome. Curr. Hypertens. Rep. 2018, 20, 12. [Google Scholar] [CrossRef] [Green Version]
- Moore, J.X.; Chaudhary, N.; Akinyemiju, T. Metabolic Syndrome Prevalence by Race/Ethnicity and Sex in the United States, National Health and Nutrition Examination Survey, 1988–2012. Prev. Chronic Dis. 2017, 14, E24. [Google Scholar] [CrossRef] [Green Version]
- Aguilar, M.; Bhuket, T.; Torres, S.; Liu, B.; Wong, R.J. Prevalence of the metabolic syndrome in the United States, 2003–2012. JAMA 2015, 313, 1973–1974. [Google Scholar] [CrossRef]
- Piko, P.; Dioszegi, J.; Sandor, J.; Adany, R. Changes in the Prevalence of Metabolic Syndrome and Its Components as Well as in Relevant Preventive Medication between 2006 and 2018 in the Northeast Hungarian Population. J. Pers. Med. 2021, 11, 52. [Google Scholar] [CrossRef]
- Piko, P.; Dioszegi, J.; Kosa, Z.; Sandor, J.; Moizs, M.; Adany, R. Changes in the Prevalence of Metabolic Syndrome, Its Components, and Relevant Preventive Medication between 2011 and 2018 in the Northeast Hungarian Roma Population. J. Pers. Med. 2021, 11, 595. [Google Scholar] [CrossRef]
- Krajcovicova-Kudlackova, M.; Blazicek, P.; Spustova, V.; Valachovicova, M.; Ginter, E. Cardiovascular risk factors in young Gypsy population. Bratisl Lek Listy 2004, 105, 256–259. [Google Scholar]
- Graae, A.S.; Hollensted, M.; Kloppenborg, J.T.; Mahendran, Y.; Schnurr, T.M.; Appel, E.V.R.; Rask, J.; Nielsen, T.R.H.; Johansen, M.O.; Linneberg, A.; et al. An adult-based insulin resistance genetic risk score associates with insulin resistance, metabolic traits and altered fat distribution in Danish children and adolescents who are overweight or obese. Diabetologia 2018, 61, 1769–1779. [Google Scholar] [CrossRef] [Green Version]
- Tancredi, M.; Rosengren, A.; Svensson, A.M.; Kosiborod, M.; Pivodic, A.; Gudbjoornsdottir, S.; Wedel, H.; Clements, M.; Dahlqvist, S.; Lind, M. Excess Mortality among Persons with Type 2 Diabetes. N. Engl. J. Med. 2015, 373, 1720–1732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aktas, G.; Alcelik, A.; Ozlu, T.; Tosun, M.; Tekce, B.K.; Savli, H.; Tekce, H.; Dikbas, O. Association between Omentin Levels and Insulin Resistance in Pregnancy. Exp. Clin. Endocr. Diabetes 2014, 122, 163–166. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Diaz, A.; Arana-Martinez, J.C.; Carbo, R.; Espinosa-Cervantes, R.; Sanchez-Munoz, F. Omentin: Role in insulin resistance, inflammation and cardiovascular protection. Arch. Cardiol. Mex. 2016, 86, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Parida, S.; Siddharth, S.; Sharma, D. Role of Omentin in Obesity Paradox in Lung Cancer. Cancers 2021, 13, 275. [Google Scholar] [CrossRef] [PubMed]
- Kocak, M.Z.; Aktas, G.; Atak, B.M.; Duman, T.T.; Yis, O.M.; Erkus, E.; Savli, H. Is Neuregulin-4 a predictive marker of microvascular complications in type 2 diabetes mellitus? Eur. J. Clin. Invest. 2020, 50, e13206. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.Y.; Chan, L.; Zhou, S.W. Omentin: Linking Metabolic Syndrome and Cardiovascular Disease. Curr. Vasc. Pharmacol. 2014, 12, 136–143. [Google Scholar] [CrossRef]
- Proctor, M.J.; McMillan, D.C.; Horgan, P.G.; Fletcher, C.D.; Talwar, D.; Morrison, D.S. Systemic Inflammation Predicts All-Cause Mortality: A Glasgow Inflammation Outcome Study. PLoS ONE 2015, 10, e0116206. [Google Scholar] [CrossRef] [Green Version]
- Kanaya, A.M.; Herrington, D.; Vittinghoff, E.; Ewing, S.K.; Liu, K.; Blaha, M.J.; Dave, S.S.; Qureshi, F.; Kandula, N.R. Understanding the high prevalence of diabetes in U.S. south Asians compared with four racial/ethnic groups: The MASALA and MESA studies. Diabetes Care 2014, 37, 1621–1628. [Google Scholar] [CrossRef] [Green Version]
- European Union Agency for Fundamental Rights. Roma and Travelers in Six Countries; Publications Office of the European Union: Luxembourg, 2020. [Google Scholar]
- Gan, W.; Walters, R.G.; Holmes, M.V.; Bragg, F.; Millwood, I.Y.; Banasik, K.; Chen, Y.; Du, H.; Iona, A.; Mahajan, A.; et al. Evaluation of type 2 diabetes genetic risk variants in Chinese adults: Findings from 93,000 individuals from the China Kadoorie Biobank. Diabetologia 2016, 59, 1446–1457. [Google Scholar] [CrossRef] [Green Version]
- Walford, G.A.; Porneala, B.C.; Dauriz, M.; Vassy, J.L.; Cheng, S.; Rhee, E.P.; Wang, T.J.; Meigs, J.B.; Gerszten, R.E.; Florez, J.C. Metabolite traits and genetic risk provide complementary information for the prediction of future type 2 diabetes. Diabetes Care 2014, 37, 2508–2514. [Google Scholar] [CrossRef] [Green Version]
- Merzah, M.; Kosa, Z.; Sandor, J.; Natae, S.; Piko, P.; Adany, R.; Fiatal, S. Roma Socioeconomic Status Has a Higher Impact on Smoking Behaviour than Genetic Susceptibility. Int. J. Environ. Res. Public Health 2021, 18, 3206. [Google Scholar] [CrossRef]
- Dioszegi, J.; Fiatal, S.; Toth, R.; Moravcsik-Kornyicki, A.; Kosa, Z.; Sandor, J.; McKee, M.; Adany, R. Distribution Characteristics and Combined Effect of Polymorphisms Affecting Alcohol Consumption Behaviour in the Hungarian General and Roma Populations. Alcohol Alcohol. 2017, 52, 104–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiatal, S.; Piko, P.; Kosa, Z.; Sandor, J.; Adany, R. Disparities in smoking habits in Hungarian Roma and general populations. Eur. J. Public Health 2019, 29, 388. [Google Scholar] [CrossRef]
- Llanaj, E.; Vincze, F.; Kosa, Z.; Sandor, J.; Dioszegi, J.; Adany, R. Dietary Profile and Nutritional Status of the Roma Population Living in Segregated Colonies in Northeast Hungary. Nutrients 2020, 12, 2836. [Google Scholar] [CrossRef] [PubMed]
- Llanaj, E.; Vincze, F.; Kosa, Z.; Bardos, H.; Dioszegi, J.; Sandor, J.; Adany, R. Deteriorated Dietary Patterns with Regards to Health and Environmental Sustainability among Hungarian Roma Are Not Differentiated from Those of the General Population. Nutrients 2021, 13, 721. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Roma (n = 334) | Hungarian General (n = 372) | ||
|---|---|---|---|
| Mean (SD) | p value | ||
| Age (years) | 42.60 (12.18) | 44.20 (12.12) | 0.114 |
| BMI (kg/m2) | 27.54 (6.81) | 27.21 (5.39) | 0.901 |
| Fasting glucose (mmol/L) | 5.10 (1.56) | 5.27 (1.93) | 0.102 |
| Fasting insulin (mU/L) | 17.13 (21.02) | 15.46 (16.43) | 0.891 |
| HOMA—IR | 4.45 (7.00) | 3.99 (5.54) | 0.978 |
| Prevalence in % (SE) | p value | ||
| Female | 74.6 (2.4) | 54.8 (2.6) | 0.003 |
| Antihypertensive treatment | 30.2 (2.5) | 28.5 (2.3) | 0.877 |
| Lipid-lowering treatment | 12.0 (1.8) | 6.7 (1.3) | 0.228 |
| Antidiabetic treatment | 11.1 (1.7) | 5.9 (1.2) | 0.245 |
| Elevated fasting glucose (≥7 mmol/L) | 9.0 (1.6) | 8.1 (1.4) | 0.670 |
| Elevated fasting insulin (>20 mU/L) | 22.5 (2.3) | 20.4 (2.1) | 0.499 |
| Elevated HOMA—IR (>3.63) | 30.2 (2.5) | 29.6 (2.4) | 0.846 |
| No. | SNP | Risk Allele | Genetic Model | Beta (95% CI) | p Value |
|---|---|---|---|---|---|
| 1 | rs7961581 | T | codominant | 1.042 (0.406–1.679) | 0.001 |
| 2 | rs1801282 | C | dominant | 2.084 (0.574–3.594) | 0.007 |
| 3 | rs6822892 | G | dominant | 0.492 (0.041–0.943) | 0.033 |
| 4 | rs13266634 | T | recessive | 0.933 (0.070–1.797) | 0.034 |
| 5 | rs4430796 | A | codominant | 0.574 (0.023–1.125) | 0.041 |
| 6 | rs10010131 | A | codominant | 0.522 (−0.073–1.117) | 0.085 |
| 7 | rs7903146 | T | dominant | 0.349 (−0.058–0.756) | 0.093 |
| 8 | rs5219 | T | dominant | 0.335 (−0.073–0.743) | 0.107 |
| 9 | rs731839 | G | recessive | 0.502 (−0.139–1.143) | 0.124 |
| 10 | rs459193 | A | recessive | 0.622 (−0.228–1.471) | 0.151 |
| 11 | rs4402960 | T | dominant | 0.301 (−0.120–0.722) | 0.160 |
| 12 | rs308971 | A | dominant | 1.030 (−0.480–2.541) | 0.181 |
| 13 | rs10811661 | T | recessive | 0.296 (−0.149–0.741) | 0.192 |
| 14 | rs4607103 | T | codominant | 0.393 (−0.213–0.999) | 0.204 |
| 15 | rs7754840 | G | dominant | 0.514 (−0.279–1.307) | 0.204 |
| 16 | rs3822072 | A | recessive | 0.306 (−0.167–0.778) | 0.205 |
| 17 | rs7578597 | C | codominant | 0.593 (−0.473–1.660) | 0.275 |
| 18 | rs1111875 | T | recessive | 0.294 (−0.299–0.888) | 0.331 |
| 19 | rs2943645 | T | recessive | 0.200 (−0.208–0.609) | 0.335 |
| 20 | rs780094 | C | codominant | 0.240 (−0.320–0.799) | 0.401 |
| 21 | rs4846565 | G | recessive | 0.163 (−0.246–0.572) | 0.433 |
| 22 | rs10195252 | T | recessive | 0.155 (−0.258–0.567) | 0.462 |
| 23 | rs10923931 | T | dominant | 0.188 (−0.319–0.695) | 0.467 |
| 24 | rs8050136 | C | dominant | 0.164 (−0.360–0.687) | 0.540 |
| 25 | rs4865796 | G | recessive | 0.188 (−0.546–0.922) | 0.615 |
| 26 | rs864745 | C | recessive | 0.109 (−0.380–0.597) | 0.662 |
| 27 | rs564398 | C | codominant | 0.127 (−0.469–0.723) | 0.676 |
| 28 | rs2745353 | T | dominant | 0.041 (−0.422–0.505) | 0.861 |
| 29 | rs2237892 | T | dominant | 0.045 (−0.663–0.753) | 0.901 |
| Low Risk (oGRS: 12 or Less) | Medium Risk (oGRS: 13–15) | High Risk (oGRS: 16 and More) | p for Trend | |
|---|---|---|---|---|
| Prevalence of IR in % (n) | ||||
| Roma | 15.9 (11) | 27.4 (48) | 46.7 (42) | <0.001 |
| Hungarian general | 23.3 (24) | 32.5 (55) | 31.0 (31) | 0.224 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piko, P.; Werissa, N.A.; Adany, R. Genetic Susceptibility to Insulin Resistance and Its Association with Estimated Longevity in the Hungarian General and Roma Populations. Biomedicines 2022, 10, 1703. https://doi.org/10.3390/biomedicines10071703
Piko P, Werissa NA, Adany R. Genetic Susceptibility to Insulin Resistance and Its Association with Estimated Longevity in the Hungarian General and Roma Populations. Biomedicines. 2022; 10(7):1703. https://doi.org/10.3390/biomedicines10071703
Chicago/Turabian StylePiko, Peter, Nardos Abebe Werissa, and Roza Adany. 2022. "Genetic Susceptibility to Insulin Resistance and Its Association with Estimated Longevity in the Hungarian General and Roma Populations" Biomedicines 10, no. 7: 1703. https://doi.org/10.3390/biomedicines10071703
APA StylePiko, P., Werissa, N. A., & Adany, R. (2022). Genetic Susceptibility to Insulin Resistance and Its Association with Estimated Longevity in the Hungarian General and Roma Populations. Biomedicines, 10(7), 1703. https://doi.org/10.3390/biomedicines10071703