
Validation of Non-Invasive Preimplantation Genetic Screening Using a Routine IVF Laboratory Workflow


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Embryo Management and Embryo Culture
2.3. Spent Culture Medium Collection
2.4. Whole-Genome Amplification and DNA Sequencing
2.5. Genetic Data Analysis
2.6. Time-Lapse System
2.7. Embryo Transfer
2.8. Statistical Analysis
3. Results
3.1. Characterization of Cell-Free DNA
3.2. Concordance of the Results from Nipgt-A and Their Corresponding Embryos/TE Biopsies
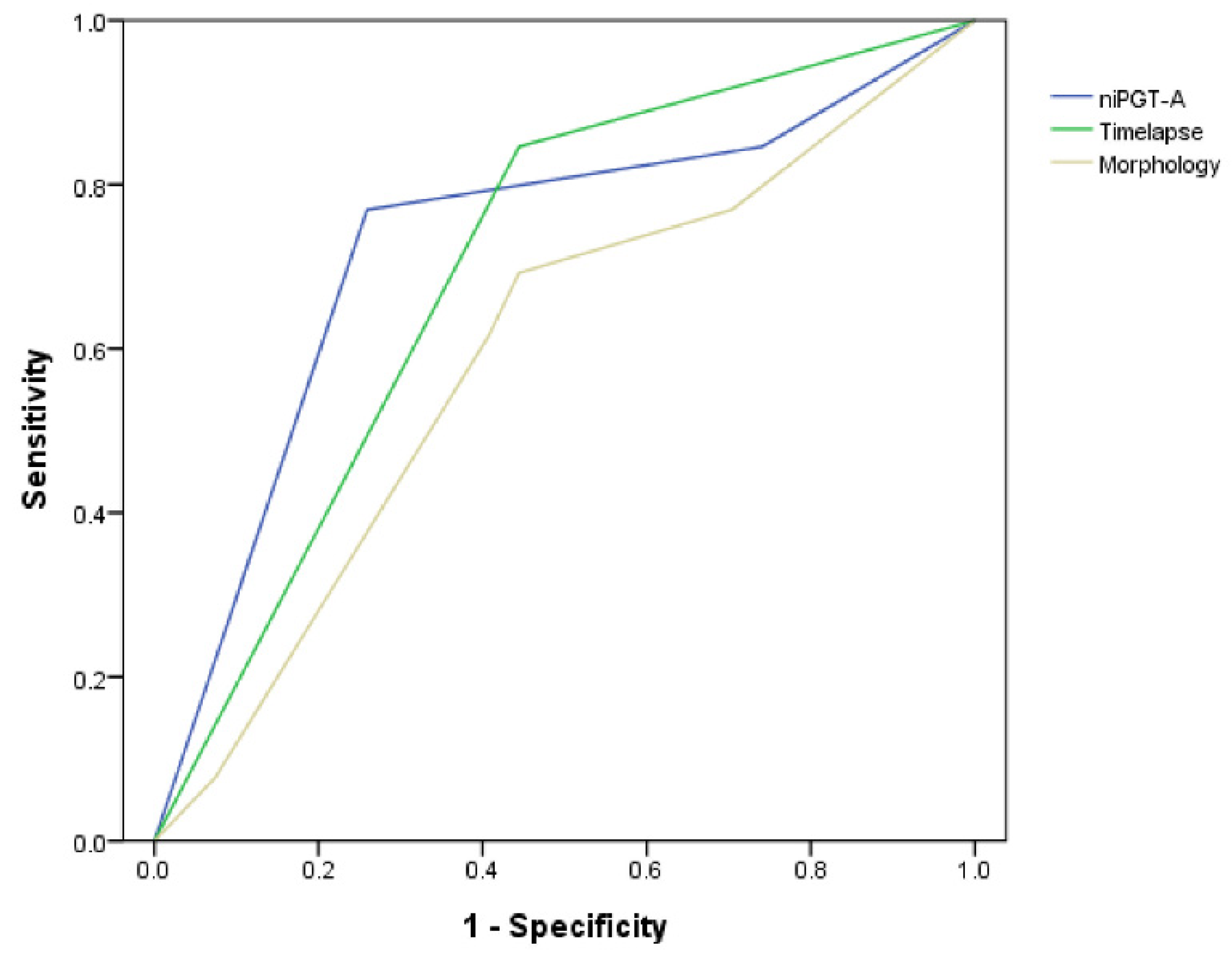
3.3. Embryo Section Algorithm Using NiPGT-A and/or Time-Lapse Morphokinetics
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sallam, H.N.; Sallam, N.H.; Sallam, S.H. Non-invasive methods for embryo selection. Facts Views Vis. ObGyn 2016, 8, 87–100. [Google Scholar] [PubMed]
- L’Heveder, A.; Jones, B.P.; Naja, R.; Serhal, P.; Nagi, J.B. Preimplantation Genetic Testing for Aneuploidy: Current Perspectives. Semin. Reprod. Med. 2021, 39, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Marin, D.; Xu, J.; Treff, N.R. Preimplantation genetic testing for aneuploidy: A review of published blastocyst reanalysis concordance data. Prenat. Diagn. 2021, 41, 545–553. [Google Scholar] [CrossRef] [PubMed]
- The use of preimplantation genetic testing for aneuploidy (PGT-A): A committee opinion. Fertil. Steril. 2018, 109, 429–436. [CrossRef] [Green Version]
- Vera-Rodriguez, M.; Rubio, C. Assessing the true incidence of mosaicism in preimplantation embryos. Fertil. Steril. 2017, 107, 1107–1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roseboom, T.J. Developmental plasticity and its relevance to assisted human reproduction. Hum. Reprod. 2018, 33, 546–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barker, D.J.P. Mothers, Babies and Health in Later Life, 2nd ed.; Churchill Livingstone: London, UK, 1998. [Google Scholar]
- Sanchez, T.; Seidler, E.A.; Gardner, D.K.; Needleman, D.; Sakkas, D. Will noninvasive methods surpass invasive for assessing gametes and embryos? Fertil. Steril. 2017, 108, 730–737. [Google Scholar] [CrossRef] [Green Version]
- Ajduk, A.; Zernicka-Goetz, M. Quality control of embryo development. Mol. Asp. Med. 2013, 34, 903–918. [Google Scholar] [CrossRef]
- Navarro-Sánchez, L.; García-Pascual, C.; Rubio, C.; Simón, C. Non-invasive preimplantation genetic testing for aneuploidies: An update. Reprod. BioMed. Online 2022, 44, 817–828. [Google Scholar] [CrossRef]
- Lan, K.C.; Huang, F.J.; Lin, Y.C.; Kung, F.T.; Hsieh, C.H.; Huang, H.W.; Tan, P.H.; Chang, S.Y. The predictive value of using a combined Z-score and day 3 embryo morphology score in the assessment of embryo survival on day 5. Hum. Reprod. 2003, 18, 1299–1306. [Google Scholar] [CrossRef]
- Good practice recommendations for the use of time-lapse technology. Hum. Reprod. Open 2020, 2020, hoaa008. [CrossRef]
- Petersen, B.M.; Boel, M.; Montag, M.; Gardner, D.K. Development of a generally applicable morphokinetic algorithm capable of predicting the implantation potential of embryos transferred on Day 3. Hum. Reprod. 2016, 31, 2231–2244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reignier, A.; Lammers, J.; Barriere, P.; Freour, T. Can time-lapse parameters predict embryo ploidy? A systematic review. Reprod. Biomed. Online 2018, 36, 380–387. [Google Scholar] [CrossRef] [Green Version]
- Desai, N.; Goldberg, J.M.; Austin, C.; Falcone, T. Are cleavage anomalies, multinucleation, or specific cell cycle kinetics observed with time-lapse imaging predictive of embryo developmental capacity or ploidy? Fertil. Steril. 2018, 109, 665–674. [Google Scholar] [CrossRef] [Green Version]
- Gardner, D.K.; Meseguer, M.; Rubio, C.; Treff, N.R. Diagnosis of human preimplantation embryo viability. Hum. Reprod. Update 2015, 21, 727–747. [Google Scholar] [CrossRef] [Green Version]
- Gombos, K.; Gálik, B.; Kalács, K.I.; Gödöny, K.; Várnagy, Á.; Alpár, D.; Bódis, J.; Gyenesei, A.; Kovács, G.L. NGS-Based Application for Routine Non-Invasive Pre-Implantation Genetic Assessment in IVF. Int. J. Mol. Sci. 2021, 22, 2443. [Google Scholar] [CrossRef] [PubMed]
- Alizadegan, A.; Dianat-Moghadam, H.; Shadman, N.; Nouri, M.; Hamdi, K.; Ghasemzadeh, A.; Akbarzadeh, M.; Sarvarian, P.; Mehdizadeh, A.; Dolati, S.; et al. Application of cell free DNA in ART. Placenta 2022, 120, 18–24. [Google Scholar] [CrossRef]
- Rubio, C.; Navarro-Sánchez, L.; García-Pascual, C.M.; Ocali, O.; Cimadomo, D.; Venier, W.; Barroso, G.; Kopcow, L.; Bahçeci, M.; Kulmann, M.I.R.; et al. Multicenter prospective study of concordance between embryonic cell-free DNA and trophectoderm biopsies from 1301 human blastocysts. Am. J. Obstet. Gynecol. 2020, 223, 751.e1–751.e3. [Google Scholar] [CrossRef]
- Yeung, Q.S.Y.; Zhang, Y.X.; Chung, J.P.W.; Lui, W.T.; Kwok, Y.K.Y.; Gui, B.; Kong, G.W.S.; Cao, Y.; Li, T.C.; Choy, K.W. A prospective study of non-invasive preimplantation genetic testing for aneuploidies (NiPGT-A) using next-generation sequencing (NGS) on spent culture media (SCM). J. Assist. Reprod. Genet. 2019, 36, 1609–1621. [Google Scholar] [CrossRef]
- Jiao, J.; Shi, B.; Sagnelli, M.; Yang, D.; Yao, Y.; Li, W.; Shao, L.; Lu, S.; Li, D.; Wang, X. Minimally invasive preimplantation genetic testing using blastocyst culture medium. Hum. Reprod. 2019, 34, 1369–1379. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Bogale, B.; Tang, Y.; Lu, S.; Xie, X.S.; Racowsky, C. Noninvasive preimplantation genetic testing for aneuploidy in spent medium may be more reliable than trophectoderm biopsy. Proc. Natl. Acad. Sci. USA 2019, 116, 14105–14112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vera-Rodriguez, M.; Diez-Juan, A.; Jimenez-Almazan, J.; Martinez, S.; Navarro, R.; Peinado, V.; Mercader, A.; Meseguer, M.; Blesa, D.; Moreno, I.; et al. Origin and composition of cell-free DNA in spent medium from human embryo culture during preimplantation development. Hum. Reprod. 2018, 33, 745–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.; Song, Z.; Yao, Y.; Huang, T.; Mao, R.; Huang, J.; Ma, Y.; Dong, X.; Huang, W.; Huang, J.; et al. Preimplantation Genetic Screening with Spent Culture Medium/Blastocoel Fluid for in Vitro Fertilization. Sci. Rep. 2018, 8, 9275. [Google Scholar] [CrossRef] [PubMed]
- Kuznyetsov, V.; Madjunkova, S.; Antes, R.; Abramov, R.; Motamedi, G.; Ibarrientos, Z.; Librach, C. Evaluation of a novel non-invasive preimplantation genetic screening approach. PLoS ONE 2018, 13, e0197262. [Google Scholar] [CrossRef]
- Ho, J.R.; Arrach, N.; Rhodes-Long, K.; Ahmady, A.; Ingles, S.; Chung, K.; Bendikson, K.A.; Paulson, R.J.; McGinnis, L.K. Pushing the limits of detection: Investigation of cell-free DNA for aneuploidy screening in embryos. Fertil. Steril. 2018, 110, 467–475 e462. [Google Scholar] [CrossRef]
- Xu, J.; Fang, R.; Chen, L.; Chen, D.; Xiao, J.P.; Yang, W.; Wang, H.; Song, X.; Ma, T.; Bo, S.; et al. Noninvasive chromosome screening of human embryos by genome sequencing of embryo culture medium for in vitro fertilization. Proc. Natl. Acad. Sci. USA 2016, 113, 11907–11912. [Google Scholar] [CrossRef] [Green Version]
- Shamonki, M.I.; Jin, H.; Haimowitz, Z.; Liu, L. Proof of concept: Preimplantation genetic screening without embryo biopsy through analysis of cell-free DNA in spent embryo culture media. Fertil. Steril. 2016, 106, 1312–1318. [Google Scholar] [CrossRef] [Green Version]
- Palini, S.; Galluzzi, L.; De Stefani, S.; Bianchi, M.; Wells, D.; Magnani, M.; Bulletti, C. Genomic DNA in human blastocoele fluid. Reprod. Biomed. Online 2013, 26, 603–610. [Google Scholar] [CrossRef] [Green Version]
- Shitara, A.; Takahashi, K.; Goto, M.; Takahashi, H.; Iwasawa, T.; Onodera, Y.; Makino, K.; Miura, H.; Shirasawa, H.; Sato, W.; et al. Cell-free DNA in spent culture medium effectively reflects the chromosomal status of embryos following culturing beyond implantation compared to trophectoderm biopsy. PLoS ONE 2021, 16, e0246438. [Google Scholar] [CrossRef]
- Chen, J.; Jia, L.; Li, T.; Guo, Y.; He, S.; Zhang, Z.; Su, W.; Zhang, S.; Fang, C. Diagnostic efficiency of blastocyst culture medium in noninvasive preimplantation genetic testing. F&S Rep. 2021, 2, 88–94. [Google Scholar] [CrossRef]
- Rubio, C.; Simon, C. Noninvasive preimplantation genetic testing for aneuploidy: Is the glass half-empty or half-full? Fertil. Steril. 2021, 115, 1426–1427. [Google Scholar] [CrossRef] [PubMed]
- Brouillet, S.; Martinez, G.; Coutton, C.; Hamamah, S. Is cell-free DNA in spent embryo culture medium an alternative to embryo biopsy for preimplantation genetic testing? A systematic review. Reprod. Biomed. Online 2020, 40, 779–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, B.; Zhang, H.; Xie, J.; Wei, Y.; Zhang, C.; Meng, L. Validation of preimplantation genetic tests for aneuploidy (PGT-A) with DNA from spent culture media (SCM): Concordance assessment and implication. Reprod. Biol. Endocrinol. 2021, 19, 41. [Google Scholar] [CrossRef] [PubMed]
- Orvieto, R.; Aizer, A.; Gleicher, N. Is there still a rationale for non-invasive PGT-A by analysis of cell-free DNA released by human embryos into culture medium? Hum. Reprod. 2021, 36, 1186–1190. [Google Scholar] [CrossRef]
- Tsai, Y.R.; Huang, F.J.; Lin, P.Y.; Kung, F.T.; Lin, Y.J.; Lin, Y.C.; Lan, K.C. Progesterone elevation on the day of human chorionic gonadotropin administration is not the only factor determining outcomes of in vitro fertilization. Fertil. Steril. 2015, 103, 106–111. [Google Scholar] [CrossRef]
- Tsai, N.C.; Su, Y.T.; Lin, Y.J.; Chiang, H.J.; Huang, F.J.; Kung, F.T.; Lan, K.C. Developmental potential of surplus morulas with delayed and/or incomplete compaction after freezing-thawing procedures. Reprod. Biol. Endocrinol. 2019, 17, 87. [Google Scholar] [CrossRef]
- Gardner, D.K.; Schoolcraft, W.B. Culture and transfer of human blastocysts. Curr. Opin. Obstet. Gynecol. 1999, 11, 307–311. [Google Scholar] [CrossRef]
- Wu, H.Y.; Lin, Y.C.; Lan, K.C. Twin live births following transfer using eight-cell cleavage stage embryos on Day 4 with developmental arrest. Taiwan J. Obstet. Gynecol. 2011, 50, 381–384. [Google Scholar] [CrossRef] [Green Version]
- Mukaida, T.; Oka, C.; Goto, T.; Takahashi, K. Artificial shrinkage of blastocoeles using either a micro-needle or a laser pulse prior to the cooling steps of vitrification improves survival rate and pregnancy outcome of vitrified human blastocysts. Human Reprod. 2006, 21, 3246–3252. [Google Scholar] [CrossRef] [Green Version]
- Meseguer, M.; Herrero, J.; Tejera, A.; Hilligsoe, K.M.; Ramsing, N.B.; Remohi, J. The use of morphokinetics as a predictor of embryo implantation. Hum. Reprod. 2011, 26, 2658–2671. [Google Scholar] [CrossRef] [Green Version]
- Campbell, A.; Fishel, S.; Bowman, N.; Duffy, S.; Sedler, M.; Hickman, C.F. Modelling a risk classification of aneuploidy in human embryos using non-invasive morphokinetics. Reprod. Biomed. Online 2013, 26, 477–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chamayou, S.; Patrizio, P.; Storaci, G.; Tomaselli, V.; Alecci, C.; Ragolia, C.; Crescenzo, C.; Guglielmino, A. The use of morphokinetic parameters to select all embryos with full capacity to implant. J. Assist. Reprod. Genet. 2013, 30, 703–710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basile, N.; Nogales Mdel, C.; Bronet, F.; Florensa, M.; Riqueiros, M.; Rodrigo, L.; García-Velasco, J.; Meseguer, M. Increasing the probability of selecting chromosomally normal embryos by time-lapse morphokinetics analysis. Fertil. Steril. 2014, 101, 699–704. [Google Scholar] [CrossRef] [PubMed]
- Rubio, I.; Kuhlmann, R.; Agerholm, I.; Kirk, J.; Herrero, J.; Escribá, M.J.; Bellver, J.; Meseguer, M. Limited implantation success of direct-cleaved human zygotes: A time-lapse study. Fertil. Steril. 2012, 98, 1458–1463. [Google Scholar] [CrossRef] [PubMed]
- Lledo, B.; Morales, R.; Ortiz, J.A.; Rodriguez-Arnedo, A.; Ten, J.; Castillo, J.C.; Bernabeu, A.; Llacer, J.; Bernabeu, R. Consistent results of non-invasive PGT-A of human embryos using two different techniques for chromosomal analysis. Reprod. Biomed. Online 2021, 42, 555–563. [Google Scholar] [CrossRef]
- Franasiak, J.M.; Forman, E.J.; Hong, K.H.; Werner, M.D.; Upham, K.M.; Treff, N.R.; Scott, R.T., Jr. The nature of aneuploidy with increasing age of the female partner: A review of 15,169 consecutive trophectoderm biopsies evaluated with comprehensive chromosomal screening. Fertil. Steril. 2014, 101, 656–663.e1. [Google Scholar] [CrossRef]
- Orvieto, R.; Shimon, C.; Rienstein, S.; Jonish-Grossman, A.; Shani, H.; Aizer, A. Do human embryos have the ability of self-correction? Reprod. Biol. Endocrinol. 2020, 18, 98. [Google Scholar] [CrossRef]
- Bolton, H.; Graham, S.J.L.; Van der Aa, N.; Kumar, P.; Theunis, K.; Fernandez Gallardo, E.; Voet, T.; Zernicka-Goetz, M. Mouse model of chromosome mosaicism reveals lineage-specific depletion of aneuploid cells and normal developmental potential. Nat. Commun. 2016, 7, 11165. [Google Scholar] [CrossRef] [Green Version]
- Singla, S.; Iwamoto-Stohl, L.K.; Zhu, M.; Zernicka-Goetz, M. Autophagy-mediated apoptosis eliminates aneuploid cells in a mouse model of chromosome mosaicism. Nat. Commun. 2020, 11, 2958. [Google Scholar] [CrossRef]
- Hammond, E.R.; Shelling, A.N.; Cree, L.M. Nuclear and mitochondrial DNA in blastocoele fluid and embryo culture medium: Evidence and potential clinical use. Hum. Reprod. 2016, 31, 1653–1661. [Google Scholar] [CrossRef] [Green Version]
- Stigliani, S.; Anserini, P.; Venturini, P.L.; Scaruffi, P. Mitochondrial DNA content in embryo culture medium is significantly associated with human embryo fragmentation. Hum. Reprod. 2013, 28, 2652–2660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Li, N.; Wang, L.; Sun, H.; Ma, M.; Wang, H.; Xu, X.; Zhang, W.; Liu, Y.; Cram, D.S.; et al. Molecular analysis of DNA in blastocoele fluid using next-generation sequencing. J. Assist. Reprod. Genet. 2016, 33, 637–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammond, E.R.; McGillivray, B.C.; Wicker, S.M.; Peek, J.C.; Shelling, A.N.; Stone, P.; Chamley, L.W.; Cree, L.M. Characterizing nuclear and mitochondrial DNA in spent embryo culture media: Genetic contamination identified. Fertil. Steril. 2017, 107, 220–228.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Munck, N.; El Khatib, I.; Abdala, A.; El-Damen, A.; Bayram, A.; Arnanz, A.; Melado, L.; Lawrenz, B.; Fatemi, H.M. Intracytoplasmic sperm injection is not superior to conventional IVF in couples with non-male factor infertility and preimplantation genetic testing for aneuploidies (PGT-A). Hum. Reprod. 2020, 35, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Liang, B.; Li, H.; He, Q.; Li, H.; Kong, L.; Xuan, L.; Xia, Y.; Shen, J.; Mao, Y.; Li, Y.; et al. Enrichment of the fetal fraction in non-invasive prenatal screening reduces maternal background interference. Sci. Rep. 2018, 8, 17675. [Google Scholar] [CrossRef]
- Hu, P.; Liang, D.; Chen, Y.; Lin, Y.; Qiao, F.; Li, H.; Wang, T.; Peng, C.; Luo, D.; Liu, H.; et al. An enrichment method to increase cell-free fetal DNA fraction and significantly reduce false negatives and test failures for non-invasive prenatal screening: A feasibility study. J. Transl. Med. 2019, 17, 124. [Google Scholar] [CrossRef]
- Qiao, L.; Yu, B.; Liang, Y.; Zhang, C.; Wu, X.; Xue, Y.; Shen, C.; He, Q.; Lu, J.; Xiang, J.; et al. Sequencing shorter cfDNA fragments improves the fetal DNA fraction in noninvasive prenatal testing. Am. J. Obstet. Gynecol. 2019, 221, 345.e1–345.e11. [Google Scholar] [CrossRef]
- Huang, B.; Tan, W.; Li, Z.; Jin, L. An artificial intelligence model (euploid prediction algorithm) can predict embryo ploidy status based on time-lapse data. Reprod. Biol. Endocrinol. 2021, 19, 185. [Google Scholar] [CrossRef]
- Chen, L.; Li, W.; Liu, Y.; Peng, Z.; Cai, L.; Zhang, N.; Xu, J.; Wang, L.; Teng, X.; Yao, Y.; et al. Non-invasive embryo selection strategy for clinical IVF to avoid wastage of potentially competent embryos. Reprod. BioMed. Online 2022. [Google Scholar] [CrossRef]
- Chen, L.; Sun, Q.; Xu, J.; Fu, H.; Liu, Y.; Yao, Y.; Lu, S.; Yao, B. A Non-invasive Chromosome Screening Strategy for Prioritizing in vitro Fertilization Embryos for Implantation. Front. Cell Dev. Biol. 2021, 9, 708322. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Variables | IVF, n = 75 | ICSI, n = 43 | p |
|---|---|---|---|
| Age, year | 34.5 (4.9) | 35.4 (4.8) | 0.35 |
| Qubit, ng/uL | 2.1 (1.7) | 1.8 (1.3) | 0.26 |
| Alive embryo on D5, n (%) | 69 (92.0%) | 38 (88.4%) | 0.53 |
| Collected D6, n (%) | 12 (16.0%) | 8 (19.0%) | 0.44 |
| QC failed, n (%) | 9 (12.0%) | 3 (7.0%) | 0.29 |
| Size peak ≥ 2, n (%) | 28 (37.3%) | 19 (44.2%) | 0.56 |
| Contamination 1 | |||
| Global | 16 (21.3%) | 8 (18.6%) | 0.40 |
| Maternal | 14 (18.7%) | 8 (18.6%) | NS |
| Paternal | 2 (2.7%) | 0 (0.0%) | NS |
| Age | Insemination Method | Collection Day | Blastocyst Morphology Grade | Qubit_SCM (ng/uL) | Karyotype_ SCM | Qubit_TE/ Whole Embryo (ng/uL) | Karyotype_ TE/Whole Embryo | Ploidy Concordance 1 |
|---|---|---|---|---|---|---|---|---|
| 39 | ICSI | 6 | 5BB | 4.86 | 45, X | 26.40 | 47, XY, +1 (×3, mos, ~70%), +16 (×3) | Y |
| 39 | ICSI | 5 | 5BB | 2.74 | 46, XX | 14.50 | 46, XX | Y |
| 39 | ICSI | 5 | 5AB | 2.18 | 46, XX | 21.40 | 46, XY | Y |
| 40 | IVF | 6 | 5AA | 0.88 | 45, X, −X (×1) | 25.80 | 48, XX, +21 (×3), +22 (×3) | Y |
| 40 | IVF | 6 | 5AB | 1.50 | 46, XX | 22.20 | 45, XY, −15 (×1) | N |
| 40 | IVF | 6 | 5BB | 1.42 | 46, XX | 22.40 | 45, X, −X (×1), +2 (×3), −14 (×1) | N |
| 38 | IVF | 5 | 5BB | 2.14 | 45, XY, −13 (×1) | 8.24 | 46, XY, +10 (×3, mos, ~50%) | N |
| 38 | IVF | 6 | 5AA | 4.32 | 46, XX | 20.40 | 46, XX | Y |
| 38 | IVF | 5 | 5AB | 1.67 | 43, X, −X (×1), −10 (×1), −20 (×1) | 23.00 | 47, XX, +21 (×3) | Y |
| 38 | IVF | 5 | 5BB | 1.99 | 47, XX, +14 (×3) | 24.40 | 46, XX | N |
| 39 | ICSI | 6 | 5CC | 1.96 | 49, X, +12 (×3), +17 (×4), +22 (×3) | high | 48XY, +3 (×3), +22 (×3) | Y |
| 39 | ICSI | 5 | 5AA | 1.19 | 46, XX | high | 47XX, +3 (×3) | N |
| 39 | ICSI | 6 | 5AB | 0.86 | 46, XX | high | XY, Mosaic partial duplication of chromosome 14, 50% | N |
| 32 | ICSI | 5 | 5AB | 0.45 | 46, XX | high | 46, XX | Y |
| 32 | ICSI | 6 | 5AB | 4.10 | 46, XX | high | 46, XX | Y |
| 32 | ICSI | 6 | 5BC | 3.86 | 46, XX | high | XX, Multiple chromosomal abnormalities | N |
| 41 | ICSI | 6 | 5CC | 1.92 | 46, XX, −2 (×1), −5 (×1), +7 (×3), +14 (×3) | high | XX, Monosomy 2 & 5; Trisomy 7; Mosaic trisomy 14 | Y |
| 41 | ICSI | 6 | 5BB | 0.82 | 46, XX | high | 47, XY, +20 (×3) | N |
| 40 | IVF | 5 | 5AA | 0.48 | QC fail | 14.80 | 45, XY, −22 (×1) | N/A |
| 40 | IVF | 5 | 5AB | 0.45 | QC fail | 13.00 | 47, XY, +22 (×3) | N/A |
| 40 | IVF | 5 | 5AB | 0.69 | 46, XY | 13.60 | 46, XY, −16 (×1), +17 (×3) | N |
| 40 | IVF | 5 | 5BB | 0.38 | QC fail | 15.40 | 46, XY, +16 (×3), −22 (×1) | N/A |
| 40 | IVF | 5 | 5BB | 0.17 | QC fail | 15.10 | 45, XX, −10 (×1) | N/A |
| 40 | IVF | 5 | 5AB | 1.96 | 46, XX | 11.30 | 46, XX | Y |
| 40 | IVF | 5 | 5AB | 1.81 | 46, XX | 11.00 | 46, XY | Y |
| 40 | IVF | 5 | 5AA | 3.14 | 46, XX | 9.58 | 46, XX | Y |
| 40 | IVF | 5 | 5AB | 1.03 | 47, XX, +16 (×3) | 17.40 | 45, XX, −16 (×1) | Y |
| 40 | IVF | 5 | 5BA | 1.61 | 46, XX | 8.74 | 46, XX | Y |
| 40 | IVF | 5 | 5BB | 0.59 | QC fail | 10.80 | 48, XX, +6 (×3), +15 (×3) | N/A |
| 39 | ICSI | 3 | Arrest | 1.48 | 44, XY, −2 (×1), −18 (×1) | 22.40 | 51, XX, +10 (×3), +18 (×3), +20 (×3), +22 (×4) | Y |
| 39 | ICSI | 3 | Arrest | 3.70 | 48, XX, +1 (×3), +18 (×3) | 26.60 | 47, XX, +18 (×3) | Y |
| 39 | ICSI | 3 | Arrest | 1.53 | 45, X | 26.00 | 47, XXY, +X (×2) | Y |
| 40 | IVF | 3 | Arrest | 1.05 | 47, XXY, +X (×2) | 33.40 | 48, XXY, +X (×2), +13 (×3) | Y |
| 40 | IVF | 3 | Arrest | 2.32 | 46, XX | 25.80 | 46, XY | Y |
| 40 | IVF | 3 | Arrest | 1.65 | 46, XX, −8 (×1), +13 (×3) | 21.80 | 47, XXY, +X (×2), +11 (×3), +15 (×3), −21 (×1) | Y |
| 40 | IVF | 3 | Arrest | 0.69 | QC fail | 29.00 | 46, XY | N/A |
| 40 | IVF | 3 | Arrest | 0.12 | QC fail | 6.44 | 46, XX | N/A |
| 40 | IVF | 3 | Arrest | 1.57 | QC fail | 26.60 | 47, XXY, +X (×2) | N/A |
| 40 | IVF | 3 | Arrest | 0.55 | QC fail | 24.00 | 50, X, −Y(×0), Multiple chromosomal abnormalities | N/A |
| 40 | IVF | 3 | Arrest | 3.48 | 46, XX | 26.40 | 44, XX, −14 (×1), −19 (×1) | N |
| No. | Day | Blastocyst Grading | TE PGT-A | niPGT-A (SCM) | CC2 (T3–T2) | CC3 (T5–T3) | T5–T2 | S2 (T4–T3) | Reverse Cleavage |
|---|---|---|---|---|---|---|---|---|---|
| 1 | D5 | 5AB | Euploidy | Euploidy | |||||
| 2 | D6 | 5AB | Euploidy | Euploidy | |||||
| 3 | D6 | 5AA | Euploidy | Euploidy | |||||
| 4 | D5 | 5AB | Euploidy | Euploidy | |||||
| 5 | D5 | 5AA | Euploidy | Euploidy | |||||
| 6 | D5 | 5BB | Euploidy | Euploidy | |||||
| 7 | D5 | 5AB | Euploidy | Euploidy | |||||
| 8 | D5 | 5AB | Euploidy | Euploidy | |||||
| 9 | D5 | 5BA | Euploidy | Euploidy | |||||
| 10 | D5 | 5BB | Euploidy | Aneuploidy | |||||
| 11 | D5 | 5BB | Mosaicism | Aneuploidy | |||||
| 12 | D5 | 5AA | Aneuploidy | Euploidy | |||||
| 13 | D6 | 5BC | Aneuploidy | Euploidy | |||||
| 14 | D6 | 5BB | Aneuploidy | Euploidy | |||||
| 15 | D5 | 5AB | Aneuploidy | Euploidy | |||||
| 16 | D5 | 5AB | Aneuploidy | Aneuploidy | |||||
| 17 | D6 | 5BB | Mosaicism | Aneuploidy | |||||
| 18 | D6 | 5AA | Aneuploidy | Aneuploidy | |||||
| 19 | D6 | 5AB | Mosaicism | Euploidy | |||||
| 20 | D6 | 5CC | Mosaicism | Aneuploidy | |||||
| 21 | D5 | 5AB | Aneuploidy | Aneuploidy | |||||
| 22 | D6 | 5BB | Aneuploidy | Euploidy | |||||
| 23 | D5 | 5AB | Aneuploidy | Aneuploidy | |||||
| 24 | D5 | 5BB | Aneuploidy | QC fail | |||||
| 25 | D5 | 5AA | Aneuploidy | QC fail | |||||
| 26 | D5 | 5AB | Aneuploidy | QC fail | |||||
| 27 | D5 | 5BB | Aneuploidy | QC fail | |||||
| 28 | D5 | 5BB | Aneuploidy | QC fail | |||||
| 29 | D6 | 5CC | Aneuploidy | Aneuploidy | |||||
| 30 | Arrest | Euploidy | QC fail | ||||||
| 31 | Arrest | Euploidy | QC fail | ||||||
| 32 | Arrest | Euploidy | Euploidy | ||||||
| 33 | Arrest | Aneuploidy | Aneuploidy | ||||||
| 34 | Arrest | Aneuploidy | Aneuploidy | ||||||
| 35 | Arrest | Aneuploidy | Aneuploidy | ||||||
| 36 | Arrest | Aneuploidy | Aneuploidy | ||||||
| 37 | Arrest | Aneuploidy | Aneuploidy | ||||||
| 38 | Arrest | Aneuploidy | QC fail | ||||||
| 39 | Arrest | Aneuploidy | Euploidy | ||||||
| 40 | Arrest | Aneuploidy | QC fail |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsai, N.-C.; Chang, Y.-C.; Su, Y.-R.; Lin, Y.-C.; Weng, P.-L.; Cheng, Y.-H.; Li, Y.-L.; Lan, K.-C. Validation of Non-Invasive Preimplantation Genetic Screening Using a Routine IVF Laboratory Workflow. Biomedicines 2022, 10, 1386. https://doi.org/10.3390/biomedicines10061386
Tsai N-C, Chang Y-C, Su Y-R, Lin Y-C, Weng P-L, Cheng Y-H, Li Y-L, Lan K-C. Validation of Non-Invasive Preimplantation Genetic Screening Using a Routine IVF Laboratory Workflow. Biomedicines. 2022; 10(6):1386. https://doi.org/10.3390/biomedicines10061386
Chicago/Turabian StyleTsai, Ni-Chin, Yun-Chiao Chang, Yi-Ru Su, Yi-Chi Lin, Pei-Ling Weng, Yin-Hua Cheng, Yi-Ling Li, and Kuo-Chung Lan. 2022. "Validation of Non-Invasive Preimplantation Genetic Screening Using a Routine IVF Laboratory Workflow" Biomedicines 10, no. 6: 1386. https://doi.org/10.3390/biomedicines10061386
APA StyleTsai, N.-C., Chang, Y.-C., Su, Y.-R., Lin, Y.-C., Weng, P.-L., Cheng, Y.-H., Li, Y.-L., & Lan, K.-C. (2022). Validation of Non-Invasive Preimplantation Genetic Screening Using a Routine IVF Laboratory Workflow. Biomedicines, 10(6), 1386. https://doi.org/10.3390/biomedicines10061386