
Identification of Novel Vascular Genes Downstream of Islet2 and Nr2f1b Transcription Factors

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Zebrafish Husbandry and Chemical Treatment
2.2. Morpholinos and Tol2 DNA Injection
2.3. Microarray Analysis
2.4. RNA Extraction and cDNA Preparation
2.5. Real-Time RT-PCR Analysis
2.6. Whole-Mount In Situ Hybridization and Frozen Sections
2.7. Image Acquisition and Processing
2.8. Protein Extraction and Western Blotting
2.9. Statistical Analyses
3. Results
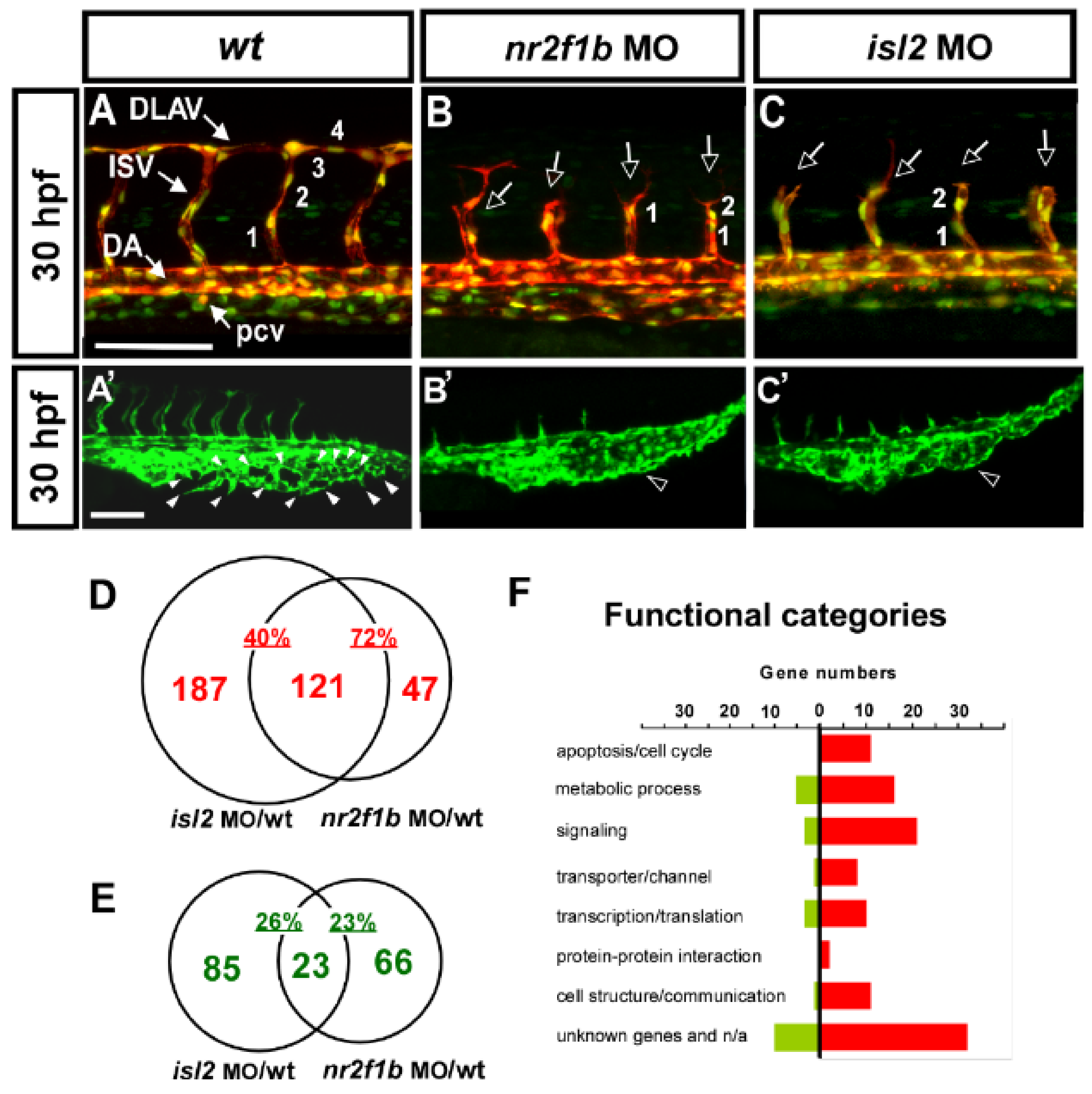
3.1. Microarray Analysis of nr2f1b and isl2 Downstream Targets

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genbank/Array ID | Gene Symbol a | Gene Name | isl2MO/wt b | nr2f1bMO/wt b |
|---|---|---|---|---|
| AI943081 | voltage-dependent calcium channel | 29.10 | 23.72 | |
| BC151913.1 | receptor activity | 14.18 | 19.74 | |
| BC066689.1 | cpn1 | carboxypeptidase N, polypeptide 1 | 12.40 | 10.85 |
| ENSDART00000073432 | RBL2 | retinoblastoma-like protein 2 | 11.31 | 11.88 |
| ZV700S00000110 | casp8 | caspase 8 | 10.15 | 10.17 |
| ZV700S00000432 | lnx1 | ligand of numb-protein X1 | 9.90 | 9.92 |
| ENSDART00000086206 | ATXN1 | ataxin-1/Spinocerebellar ataxia type 1 protein | 9.23 | 12.45 |
| ENSDART00000005808 | calponin-1-like | 8.49 | 9.00 | |
| ZV700S00000355 | phlda3 | pleckstrin homology-like domain, family A, member 3 | 8.25 | 8.03 |
| ENSDART00000016263 | probable G-protein-coupled receptor-like | 7.50 | 3.21 | |
| OTTDART00000022443 | fsta | follistatin a | 7.41 | 6.36 |
| TC254361 | bbc3 | bcl2 binding component 3 | 6.84 | 6.68 |
| OTTDART00000030315 | cdkn1a | cyclin-dependent kinase inhibitor 1A | 6.57 | 5.01 |
| OTTDART00000021597 | ftr82 | finTRIM family, member 82 | 6.50 | 7.25 |
| ENSDART00000097728 | similar to neurotoxin/C59 | 6.49 | 5.68 | |
| OTTDART00000027213 | mdm2 | murine double minute 2 homolog | 6.48 | 8.07 |
| OTTDART00000007800 | foxo3b | forkhead box O3b Transcription factor | 6.38 | 7.35 |
| ZV700S00006205 | RPS27L | ribosomal protein S27-like protein | 5.74 | 4.68 |
| OTTDART00000019467 | tp53 | tumor protein p53 | 5.74 | 5.44 |
| BC072702.1 | cyp3a65 | cytochrome P450, family 3, subfamily A, polypeptide 65 | 5.61 | 5.60 |
| OTTDART00000025224 | mmp2 | matrix metalloproteinase 2 | 5.22 | 5.99 |
| ZV700S00003980 | gadd45aa | growth arrest and DNA-damage-inducible, alpha, a | 5.07 | 4.50 |
| TC253576 | Beta-1,3-glucosyltransferase-like | 5.05 | 4.12 | |
| ENSDART00000034197 | SUSD5 | Sushi domain-containing protein 5 | 4.80 | 4.91 |
| ZV700S00006317 | igf2a | insulin-like growth factor 2a | 4.68 | 4.46 |
| OTTDART00000007033 | fos | v-fos FBJ murine osteosarcoma viral oncogene homolog | 4.54 | 4.82 |
| BC056806.1 | gtpbp1l | GTP-binding protein 1-like | 4.30 | 4.23 |
| BM571671 | protein p8 | 4.25 | 5.95 | |
| ENSDART00000081980 | LAG1 homolog, ceramide synthase 5-like | 4.24 | 4.26 | |
| TC250557 | slc4a2a | solute carrier family 4, anion exchanger, member 2a | 4.23 | 2.93 |
| OTTDART00000007990 | cx32.2 | connexin 32.2/cell junction | 4.17 | 6.50 |
| TC242899 | crfb1 | cytokine receptor family member b1 | 4.07 | 4.06 |
| ENSDART00000097555 | PAPPA | Pappalysin-1 Precursor | 4.05 | 2.82 |
| AW826461 | sept8b | septin 8b Septin-like protein | 3.98 | 3.29 |
| ZV700S00005741 | pik3r1 | phosphoinositide-3-kinase regulatory subunit | 3.98 | 3.61 |
| TC245426 | ect2 | epithelial-cell-transforming sequence 2 oncogene | 3.90 | 3.71 |
| AI793818 | chrngl | cholinergic receptor, nicotinic, gamma like | 3.90 | 3.75 |
| NM_131862 | jag2 | jagged 2 | 3.86 | 4.95 |
| TC238338 | otop1 | otopetrin 1 | 3.85 | 3.23 |
| OTTDART00000029706 | similar to vertebrate doublecortin domain-containing 2, DCDC2 | 3.78 | 3.53 | |
| OTTDART00000025192 | f2 | coagulation factor II, thrombin/(prothrombin) | 3.73 | 3.58 |
| ZV700S00004623 | cdkal1 | CDK5 regulatory subunit-associated protein 1-like 1 | 3.72 | 2.89 |
| OTTDART00000017384 | aqp12 | aquaporin 12 | 3.68 | 3.90 |
| ZV700S00005526 | sesn3 | Sestrin 3 | 3.66 | 5.11 |
| ZV700S00000935 | hdac4 | histone deacetylase 4 chromatin modification | 3.65 | 3.01 |
| OTTDART00000010432 | ncl1 | nicalin | 3.52 | 3.28 |
| ZV700S00003982 | rlbp1b | retinaldehyde binding protein 1b | 3.42 | 3.09 |
| ZV700S00003517 | rhoub | ras homolog gene family, member Ub | 3.37 | 3.18 |
| ZV700S00000580 | ccng1 | cyclin G1 | 3.36 | 3.40 |
| BI984278_R; BI984278 | taf1a | TATA box binding protein (TBP)-associated factor | 3.32 | 2.70 |
| OTTDART00000027172 | cry-dash | cryptochrome DASH. (photolyase-like) | 3.26 | 3.53 |
| ZV700S00000920 | phlda2 | pleckstrin homology-like domain, family A, member 2 | 3.25 | 3.02 |
| TC249636 | C6 | complement component 6 | 3.23 | 2.78 |
| BQ419558 | nardilysin isoform b (N-arginine dibasic convertase) | 3.22 | 2.79 | |
| OTTDART00000030385 | qdpra | quinoid dihydropteridine reductase | 3.20 | 2.70 |
| ENSDART00000049419 | jagged2-like | 3.18 | 3.93 | |
| ZV700S00004928 | rasl11b | RAS-like, family 11, member B | 3.16 | 3.33 |
| ZV700S00003200 | fabp1b.1 | Fatty acid binding protein 1b | 3.12 | 2.89 |
| ENSDART00000059592 | MEP1B | Meprin A beta | 3.12 | 3.17 |
| ZV700S00002841 | scarb2 | scavenger receptor class B, member 2 | 2.72 | 3.40 |
| BC053278.1 | itm2bb | integral membrane protein 2b | 2.68 | 3.14 |
| BI980126 | SEC15L2 | SEC15-like 1 isoform a | 2.95 | 2.70 |
| BC153975 | rpz5 | Rapunzel 5 | 2.90 | 4.01 |
| BC064309.1 | tmem38a | transmembrane protein 38A/cation channel transport activity | 2.80 | 2.56 |
| ENSDART00000065340 | tlr9 | Toll-like receptor 9/innate immune response | 2.79 | 4.52 |
| OTTDART00000017062 | similar to vertebrate capicua homolog | 2.78 | 3.11 | |
| NM_001007784 | crygn1 | crystallin, gamma N1 | 2.72 | 3.33 |
| ENSDART00000080490 | similar to peroxidasin | 2.64 | 2.57 | |
| OTTDART00000025182 | slc38a3 | solute carrier family 38, member 3 | 2.54 | 2.71 |
| NM_131156 | krt5 | keratin 5 | 2.54 | 2.68 |
| Genbank/Array ID | Gene Symbol a | Gene Name | isl2MO/wt b | nr2f1bMO/wt b |
|---|---|---|---|---|
| ENSDART00000008778 | extracellular-glutamate-gated ion channel activity | 0.24 | 0.23 | |
| OTTDART00000007452 | eef1db | translation elongation factor-1, delta,b | 0.33 | 0.19 |
| OTTDART00000028819 | nsdhl | NAD(P) dependent steroid dehydrogenase-like | 0.33 | 0.31 |
| NM_213309 | stap2b | signal transducing adaptor family member 2b | 0.34 | 0.32 |
| NM_001002580 | pmepa1 | prostate transmembrane protein, androgen-induced 1 | 0.36 | 0.37 |
| ENSDART00000066927 | N/A | 0.37 | 0.41 | |
| BC085576 | dhrs13a.1 | oxidoreductase activity | 0.37 | 0.39 |
| NM_001082856 | rorb | RAR-related orphan receptor B (NR1F2) | 0.37 | 0.29 |
| AL922441 | N/A | 0.37 | 0.38 | |
| CD605872_R | DNA helicase B | 0.38 | 0.37 | |
| ENSDART00000027268 | otpa | orthopedia homolog a | 0.39 | 0.37 |
| ENSDART00000074537 | shisa9b | protein shisa-9B | 0.39 | 0.30 |
| ENSDART00000077671 | similar to Aldehyde dehydrogenase 9 family, member A1 like 1 | 0.39 | 0.38 | |
| NM_001025174 | fam110b | family with sequence similarity 110, member B | 0.42 | 0.42 |
| TC246456 | metallocarboxypeptidase activity | 0.42 | 0.29 | |
| ENSDART00000111589 | hapln1a | hyaluronan and proteoglycan link protein 1a | 0.43 | 0.45 |
| ENSDART00000097296 | similar to alpha-2,3-sialyltransferase ST3Gal I-r2 | 0.43 | 0.36 | |
| ENSDART00000058866 | nes | Nestin | 0.43 | 0.36 |
| CR545462 | WASF1 | similar to vertebrate WAS protein family, member 1 | 0.44 | 0.34 |
| ZV700S00001176 | ccn1 | cyclin I | 0.45 | 0.47 |
| NM_001003834 | top2a | DNA topoisomerase (DNA) II alpha | 0.45 | 0.35 |
| BC164898.1 | her4.1 | hairy-related 4.1 | 0.46 | 0.42 |
| NM_205585 | sfrp1a | secreted frizzled-related protein 1a | 0.46 | 0.43 |
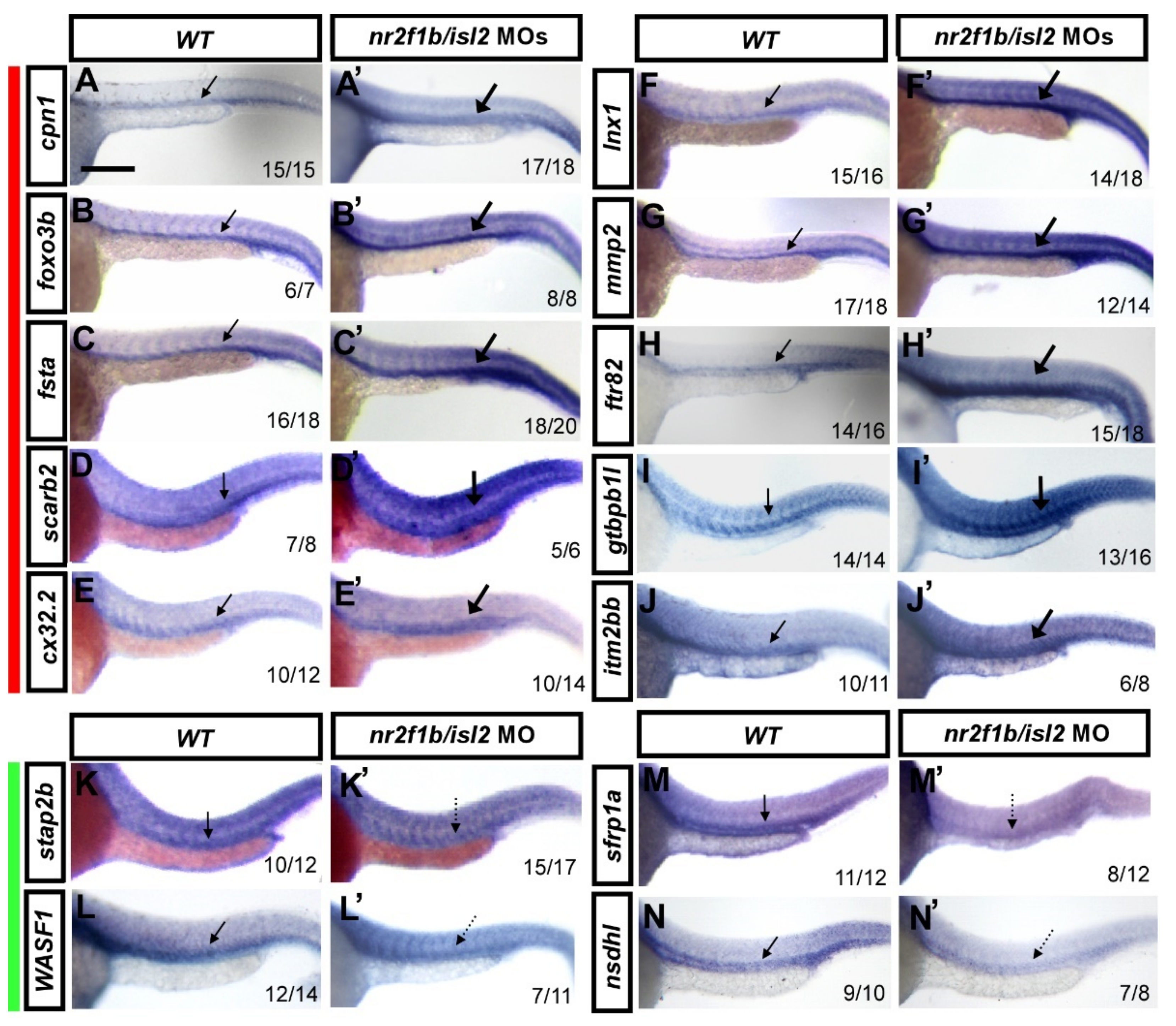
3.2. Verification of Expression Pattern of Several Potential Targets
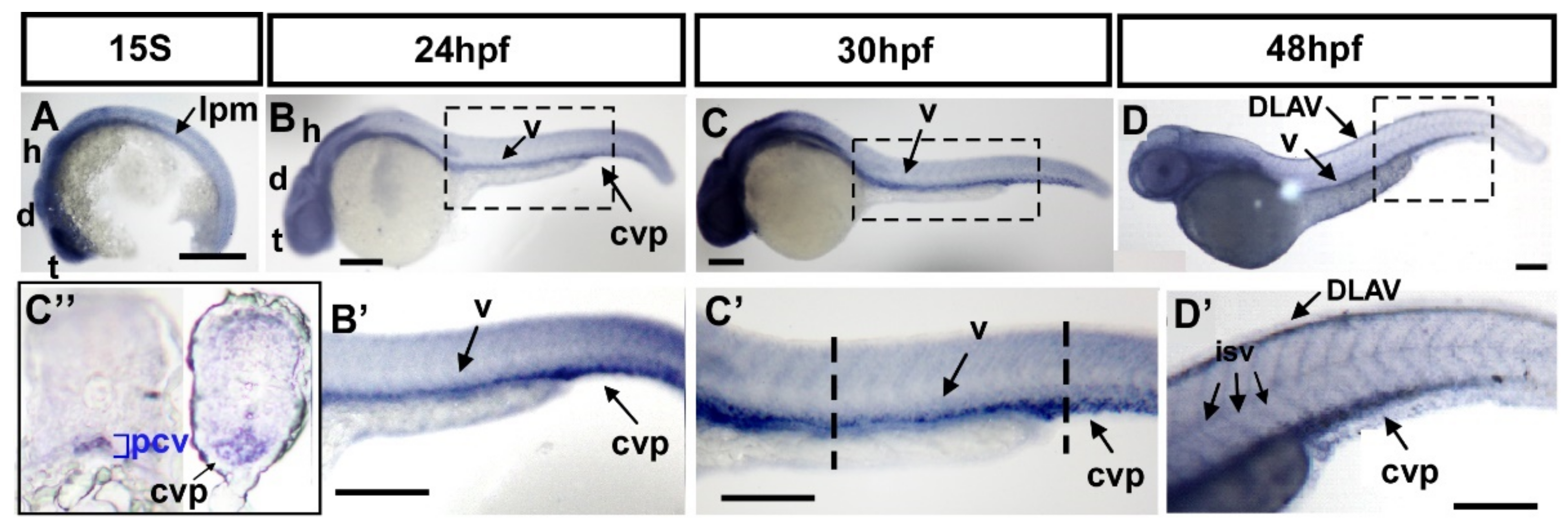
3.3. fsta Is Highly Conserved and Is Expressed in the Vasculature during Development
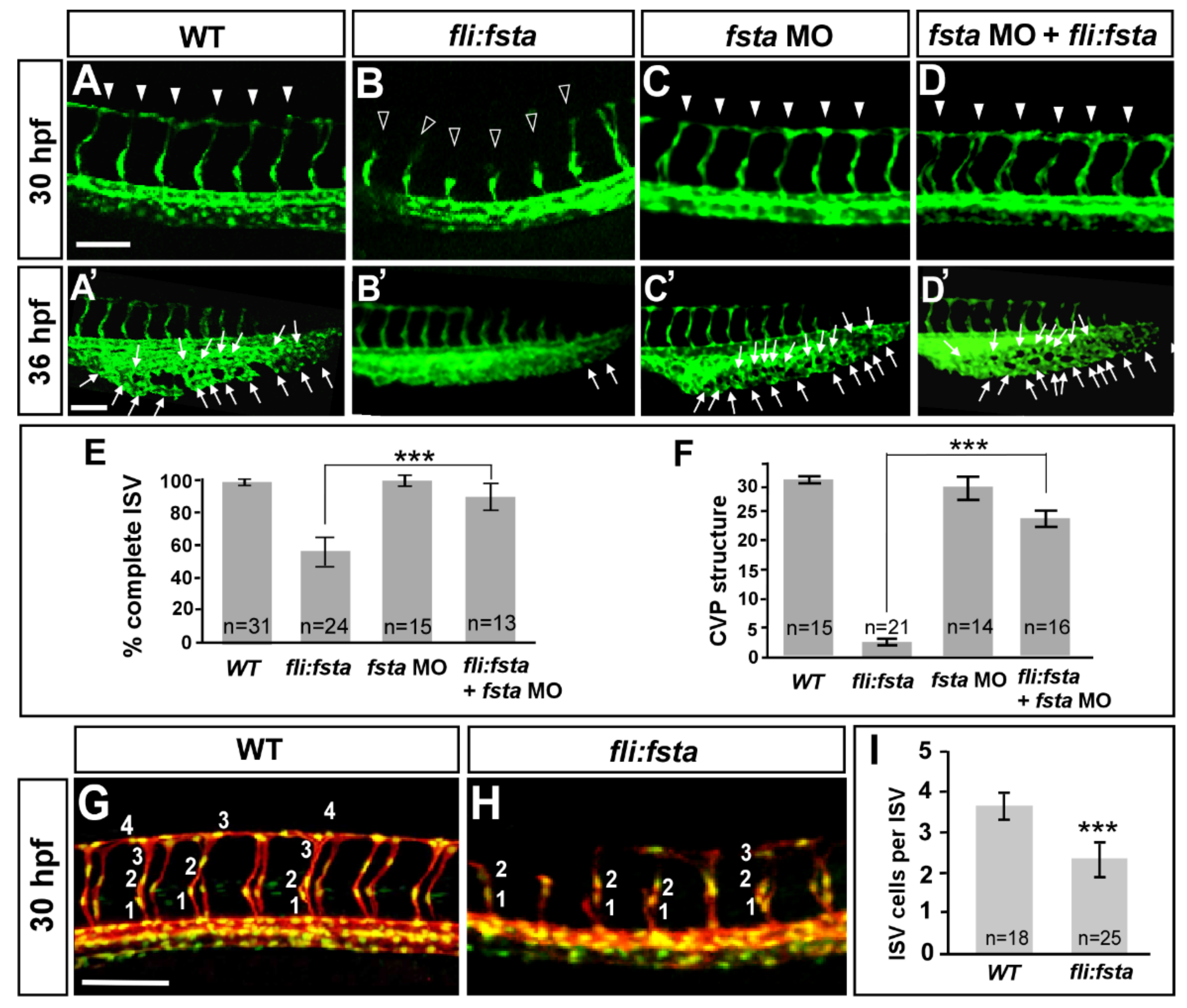
3.4. Overexpression of the fsta Gene Causes Defects in Vascular Development
3.5. Specificity of fsta Effects
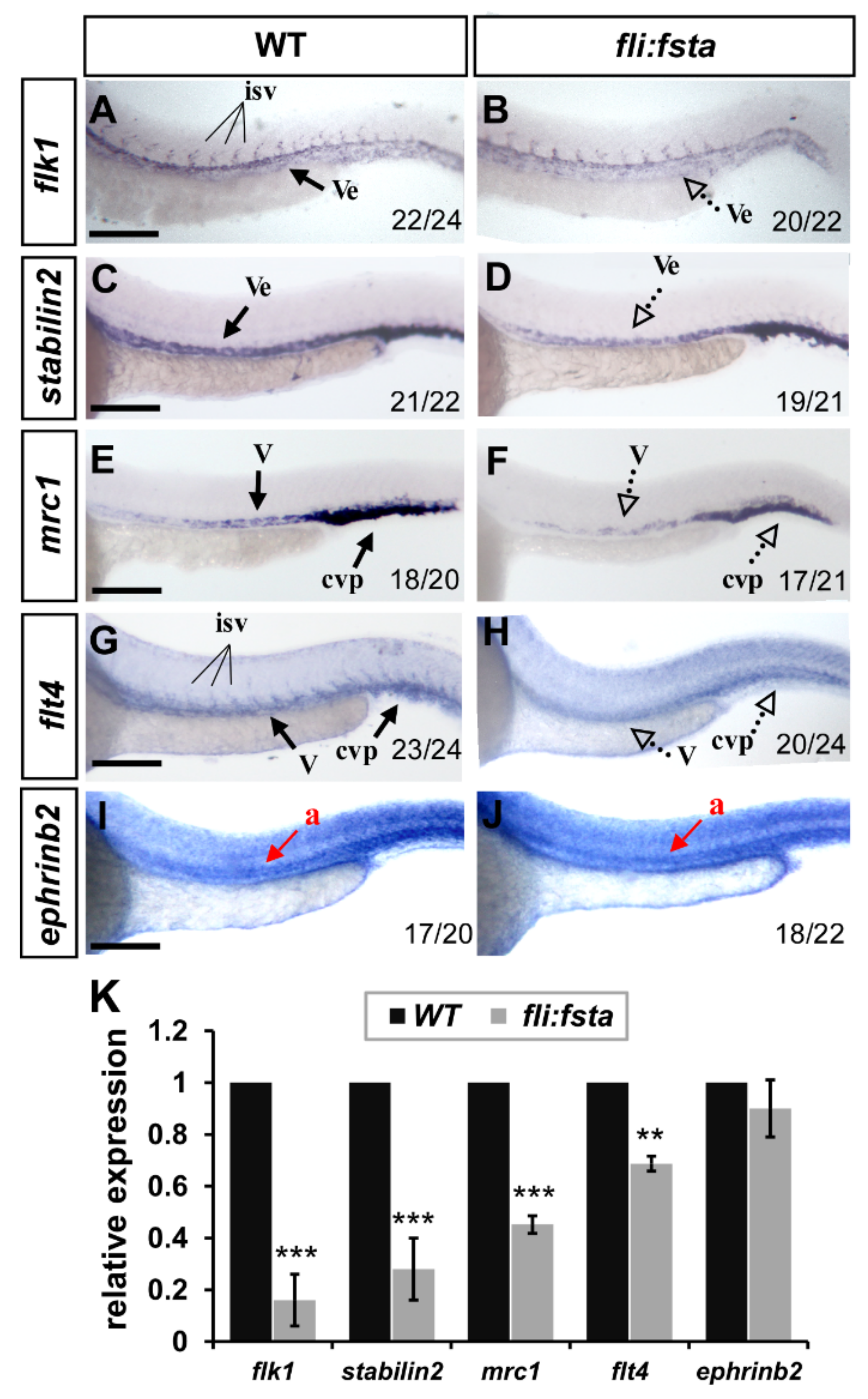
3.6. Overexpression of fsta Remodeled the Expression of Vascular Markers
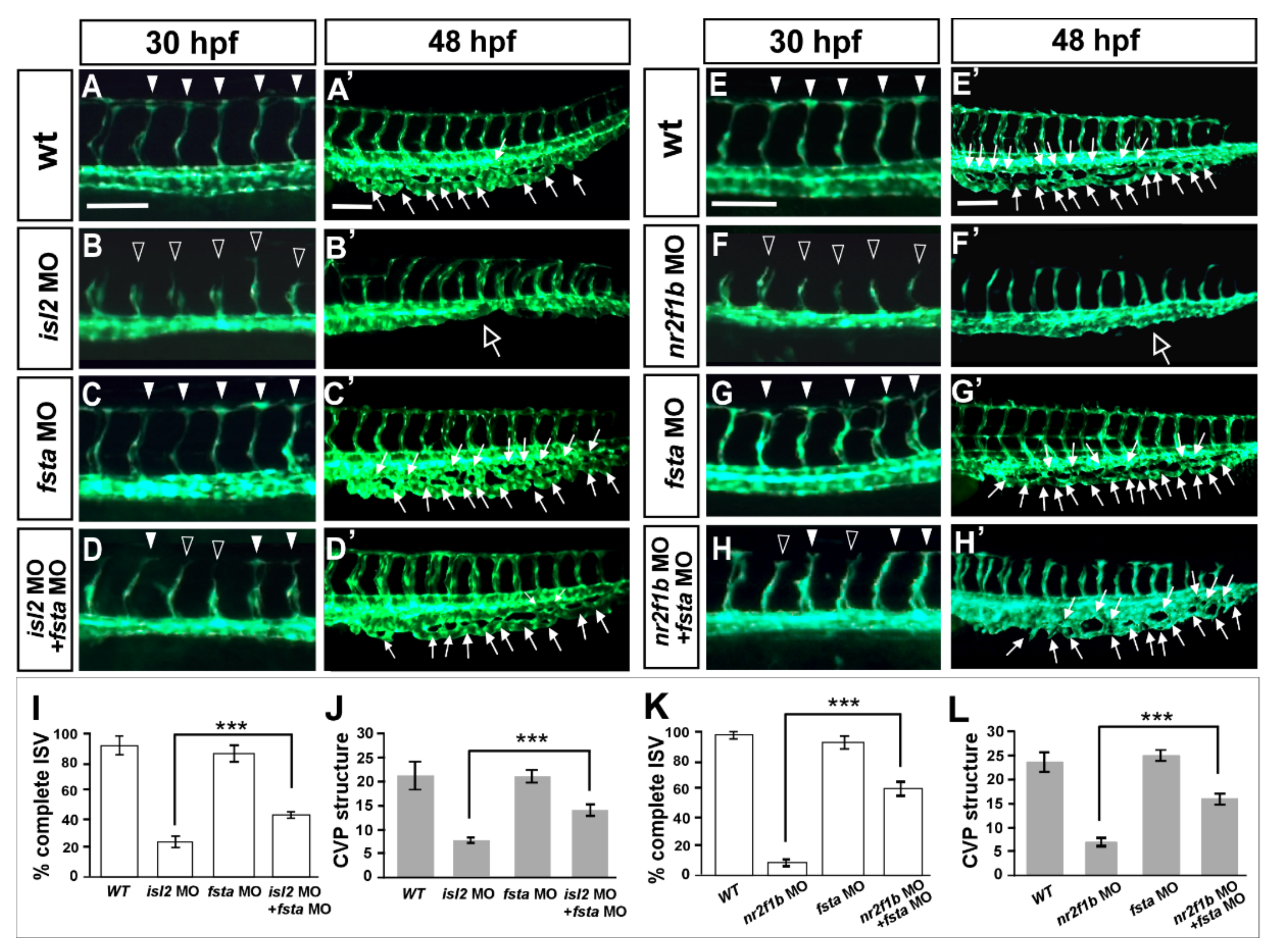
3.7. Knockdown of fsta Rescues the Vascular Defects in isl2 or nr2f1b Morphants
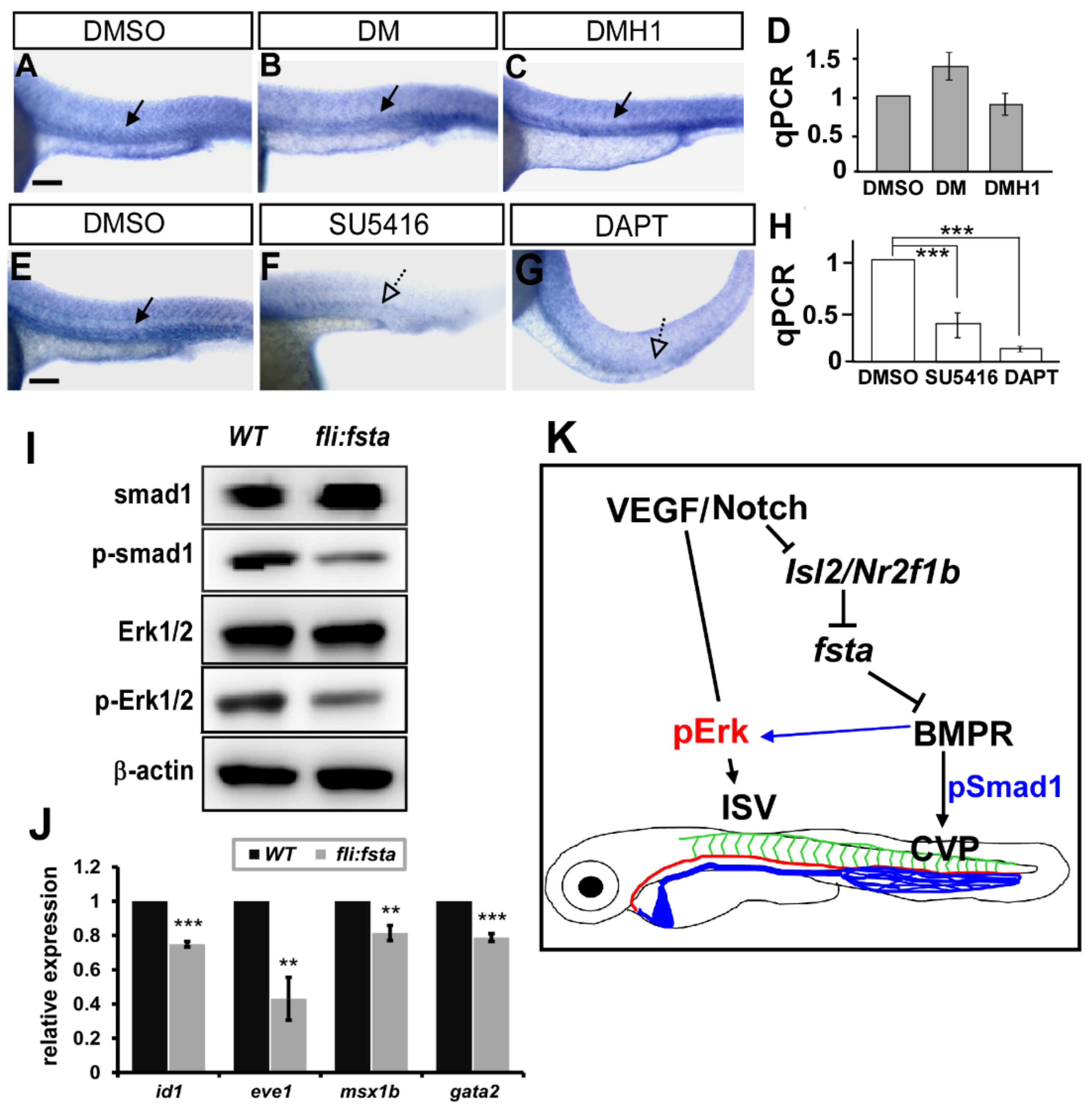
3.8. Follistatin a Regulates BMP Signals to Control Vascular Patterning
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhong, T.P. Zebrafish genetics and formation of embryonic vasculature. Curr. Top. Dev. Biol. 2005, 71, 53–81. [Google Scholar] [CrossRef] [PubMed]
- Lamont, R.E.; Childs, S. MAPping out arteries and veins. Sci. STKE 2006, 2006, pe39. [Google Scholar] [CrossRef] [PubMed]
- Conway, E.M.; Collen, D.; Carmeliet, P. Molecular mechanisms of blood vessel growth. Cardiovasc. Res. 2001, 49, 507–521. [Google Scholar] [CrossRef] [Green Version]
- Adams, R.H.; Alitalo, K. Molecular regulation of angiogenesis and lymphangiogenesis. Nat. Rev. Mol. Cell Biol. 2007, 8, 464–478. [Google Scholar] [CrossRef] [PubMed]
- Herbert, S.P.; Stainier, D.Y. Molecular control of endothelial cell behaviour during blood vessel morphogenesis. Nat. Rev. Mol. Cell Biol. 2011, 12, 551–564. [Google Scholar] [CrossRef] [Green Version]
- Childs, S.; Chen, J.-N.; Garrity, D.; Fishman, M. Patterning of angiogenesis in the zebrafish embryo. Development 2002, 129, 973–982. [Google Scholar] [CrossRef]
- Lawson, N.D.; Weinstein, B.M. In vivo imaging of embryonic vascular development using transgenic zebrafish. Dev. Biol. 2002, 248, 307–318. [Google Scholar] [CrossRef] [Green Version]
- Ellertsdottir, E.; Lenard, A.; Blum, Y.; Krudewig, A.; Herwig, L.; Affolter, M.; Belting, H.G. Vascular morphogenesis in the zebrafish embryo. Dev. Biol. 2010, 341, 56–65. [Google Scholar] [CrossRef] [Green Version]
- Schuermann, A.; Helker, C.S.; Herzog, W. Angiogenesis in zebrafish. Semin. Cell Dev. Biol. 2014, 31, 106–114. [Google Scholar] [CrossRef]
- Goi, M.; Childs, S.J. Patterning mechanisms of the sub-intestinal venous plexus in zebrafish. Dev. Biol. 2016, 409, 114–128. [Google Scholar] [CrossRef] [Green Version]
- Herpers, R.; van de Kamp, E.; Duckers, H.J.; Schulte-Merker, S. Redundant roles for sox7 and sox18 in arteriovenous specification in zebrafish. Circ. Res. 2008, 102, 12–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kume, T. Specification of arterial, venous, and lymphatic endothelial cells during embryonic development. Histol. Histopathol. 2010, 25, 637–646. [Google Scholar] [PubMed]
- Lawson, N.D.; Scheer, N.; Pham, V.N.; Kim, C.H.; Chitnis, A.B.; Campos-Ortega, J.A.; Weinstein, B.M. Notch signaling is required for arterial-venous differentiation during embryonic vascular development. Development 2001, 128, 3675–3683. [Google Scholar] [CrossRef] [PubMed]
- Zhong, T.; Childs, S.; Liu, J.; Fishman, M. Gridlock signaling pathway fashions the first embryonic artery. Nature 2001, 414, 216–220. [Google Scholar] [CrossRef]
- Lawson, N.D.; Vogel, A.M.; Weinstein, B.M. sonic hedgehog and vascular endothelial growth factor act upstream of the Notch pathway during arterial endothelial differentiation. Dev. Cell 2002, 3, 127–136. [Google Scholar] [CrossRef] [Green Version]
- Swift, M.R.; Weinstein, B.M. Arterial-venous specification during development. Circ. Res. 2009, 104, 576–588. [Google Scholar] [CrossRef] [Green Version]
- Torres-Vazquez, J.; Kamei, M.; Weinstein, B.M. Molecular distinction between arteries and veins. Cell Tissue Res. 2003, 314, 43–59. [Google Scholar] [CrossRef]
- You, L.R.; Lin, F.J.; Lee, C.T.; DeMayo, F.J.; Tsai, M.J.; Tsai, S.Y. Suppression of Notch signalling by the COUP-TFII transcription factor regulates vein identity. Nature 2005, 435, 98–104. [Google Scholar] [CrossRef]
- Gerhardt, H.; Golding, M.; Fruttiger, M.; Ruhrberg, C.; Lundkvist, A.; Abramsson, A.; Jeltsch, M.; Mitchell, C.; Alitalo, K.; Shima, D.; et al. VEGF guides angiogenic sprouting utilizing endothelial tip cell filopodia. J. Cell Biol. 2003, 161, 1163–1177. [Google Scholar] [CrossRef]
- Torres-Vazquez, J.; Gitler, A.D.; Fraser, S.D.; Berk, J.D.; Van, N.P.; Fishman, M.C.; Childs, S.; Epstein, J.A.; Weinstein, B.M. Semaphorin-plexin signaling guides patterning of the developing vasculature. Dev. Cell 2004, 7, 117–123. [Google Scholar] [CrossRef] [Green Version]
- Siekmann, A.F.; Lawson, N.D. Notch signalling limits angiogenic cell behaviour in developing zebrafish arteries. Nature 2007, 445, 781–784. [Google Scholar] [CrossRef] [PubMed]
- Wiley, D.; Kim, J.; Hao, J.; Hong, C.; Bautch, V.; Jin, S. Distinct signalling pathways regulate sprouting angiogenesis from the dorsal aorta and the axial vein. Nat. Cell Biol. 2011, 13, 686–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamik, A.; Wang, B.; Jain, M.K. Transcriptional regulators of angiogenesis. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 1936–1947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, C.; Kim, T.M.; Malik, A.B. Transcriptional regulation of endothelial cell and vascular development. Circ. Res. 2013, 112, 1380–1400. [Google Scholar] [CrossRef]
- Pereira, F.A.; Qiu, Y.; Zhou, G.; Tsai, M.J.; Tsai, S.Y. The orphan nuclear receptor COUP-TFII is required for angiogenesis and heart development. Genes Dev. 1999, 13, 1037–1049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aranguren, X.L.; Beerens, M.; Vandevelde, W.; Dewerchin, M.; Carmeliet, P.; Luttun, A. Transcription factor COUP-TFII is indispensable for venous and lymphatic development in zebrafish and Xenopus laevis. Biochem. Biophys. Res. Commun. 2011, 410, 121–126. [Google Scholar] [CrossRef]
- Swift, M.R.; Pham, V.N.; Castranova, D.; Bell, K.; Poole, R.J.; Weinstein, B.M. SoxF factors and Notch regulate nr2f2 gene expression during venous differentiation in zebrafish. Dev. Biol. 2014, 390, 116–125. [Google Scholar] [CrossRef] [Green Version]
- Li, R.F.; Wu, T.Y.; Mou, Y.Z.; Wang, Y.S.; Chen, C.L.; Wu, C.Y. Nr2f1b control venous specification and angiogenic patterning during zebrafish vascular development. J. Biomed. Sci. 2015, 22, 104. [Google Scholar] [CrossRef] [Green Version]
- Lamont, R.E.; Wu, C.Y.; Ryu, J.R.; Vu, W.; Davari, P.; Sobering, R.E.; Kennedy, R.M.; Munsie, N.M.; Childs, S.J. The LIM-homeodomain transcription factor Islet2a promotes angioblast migration. Dev. Biol. 2016, 414, 181–192. [Google Scholar] [CrossRef]
- Iemura, S.; Yamamoto, T.S.; Takagi, C.; Uchiyama, H.; Natsume, T.; Shimasaki, S.; Sugino, H.; Ueno, N. Direct binding of follistatin to a complex of bone-morphogenetic protein and its receptor inhibits ventral and epidermal cell fates in early Xenopus embryo. Proc. Natl. Acad. Sci. USA 1998, 95, 9337–9342. [Google Scholar] [CrossRef] [Green Version]
- Jiang, N.; Jin, X.; He, J.; Yin, Z. The roles of follistatin 1 in regulation of zebrafish fecundity and sexual differentiation. Biol. Reprod. 2012, 87, 54. [Google Scholar] [CrossRef] [PubMed]
- Winbanks, C.E.; Weeks, K.L.; Thomson, R.E.; Sepulveda, P.V.; Beyer, C.; Qian, H.; Chen, J.L.; Allen, J.M.; Lancaster, G.I.; Febbraio, M.A.; et al. Follistatin-mediated skeletal muscle hypertrophy is regulated by Smad3 and mTOR independently of myostatin. J. Cell Biol. 2012, 197, 997–1008. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Li, Y.; Lu, A.; Gharaibeh, B.; Ma, J.; Kobayashi, T.; Quintero, A.J.; Huard, J. Follistatin improves skeletal muscle healing after injury and disease through an interaction with muscle regeneration, angiogenesis, and fibrosis. Am. J. Pathol. 2011, 179, 915–930. [Google Scholar] [CrossRef] [PubMed]
- Moser, M.; Patterson, C. Bone morphogenetic proteins and vascular differentiation: BMPing up vasculogenesis. Thromb. Haemost. 2005, 94, 713–718. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Miralles, I.; Schisler, J.C.; Patterson, C. New insights into bone morphogenetic protein signaling: Focus on angiogenesis. Curr. Opin. Hematol. 2009, 16, 195–201. [Google Scholar] [CrossRef] [Green Version]
- Cai, J.; Pardali, E.; Sanchez-Duffhues, G.; ten Dijke, P. BMP signaling in vascular diseases. FEBS Lett. 2012, 586, 1993–2002. [Google Scholar] [CrossRef]
- Kim, J.D.; Kang, H.; Larrivee, B.; Lee, M.Y.; Mettlen, M.; Schmid, S.L.; Roman, B.L.; Qyang, Y.; Eichmann, A.; Jin, S.W. Context-dependent proangiogenic function of bone morphogenetic protein signaling is mediated by disabled homolog 2. Dev. Cell 2012, 23, 441–448. [Google Scholar] [CrossRef] [Green Version]
- Kashiwada, T.; Fukuhara, S.; Terai, K.; Tanaka, T.; Wakayama, Y.; Ando, K.; Nakajima, H.; Fukui, H.; Yuge, S.; Saito, Y.; et al. beta-Catenin-dependent transcription is central to Bmp-mediated formation of venous vessels. Development 2015, 142, 497–509. [Google Scholar] [CrossRef] [Green Version]
- Westerfield, M. The Zebrafish Book: A Guide for the Laboratory Use of Zebrafish (Danio Rerio); Institute of Neuroscience, University of Oregon: Eugene, OR, USA, 1995. [Google Scholar]
- Roman, B.L.; Pham, V.N.; Lawson, N.D.; Kulik, M.; Childs, S.; Lekven, A.C.; Garrity, D.M.; Moon, R.T.; Fishman, M.C.; Lechleider, R.J.; et al. Disruption of acvrl1 increases endothelial cell number in zebrafish cranial vessels. Development 2002, 129, 3009–3019. [Google Scholar] [CrossRef]
- Choi, J.; Dong, L.; Ahn, J.; Dao, D.; Hammerschmidt, M.; Chen, J.N. FoxH1 negatively modulates flk1 gene expression and vascular formation in zebrafish. Dev. Biol. 2007, 304, 735–744. [Google Scholar] [CrossRef] [Green Version]
- Proulx, K.; Lu, A.; Sumanas, S. Cranial vasculature in zebrafish forms by angioblast cluster-derived angiogenesis. Dev. Biol. 2010, 348, 34–46. [Google Scholar] [CrossRef] [PubMed]
- Erickson, T.; French, C.R.; Waskiewicz, A.J. Meis1 specifies positional information in the retina and tectum to organize the zebrafish visual system. Neural Dev. 2010, 5, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, B.J.; Chiu, C.C.; Chen, C.L.; Wang, W.D.; Wang, J.H.; Wen, Z.H.; Liu, W.; Chang, H.W.; Wu, C.Y. Nuclear receptor subfamily 2 group F member 1a (nr2f1a) is required for vascular development in zebrafish. PLoS ONE 2014, 9, e105939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolstad, B.M.; Irizarry, R.A.; Astrand, M.; Speed, T.P. A comparison of normalization methods for high density oligonucleotide array data based on variance and bias. Bioinformatics 2003, 19, 185–193. [Google Scholar] [CrossRef] [Green Version]
- Irizarry, R.A.; Bolstad, B.M.; Collin, F.; Cope, L.M.; Hobbs, B.; Speed, T.P. Summaries of Affymetrix GeneChip probe level data. Nucleic Acids Res. 2003, 31, e15. [Google Scholar] [CrossRef]
- Irizarry, R.A.; Hobbs, B.; Collin, F.; Beazer-Barclay, Y.D.; Antonellis, K.J.; Scherf, U.; Speed, T.P. Exploration, normalization, and summaries of high density oligonucleotide array probe level data. Biostatistics 2003, 4, 249–264. [Google Scholar] [CrossRef] [Green Version]
- Smyth, G.K.; Michaud, J.; Scott, H.S. Use of within-array replicate spots for assessing differential expression in microarray experiments. Bioinformatics 2005, 21, 2067–2075. [Google Scholar] [CrossRef]
- Benjamini, Y.; Drai, D.; Elmer, G.; Kafkafi, N.; Golani, I. Controlling the false discovery rate in behavior genetics research. Behav. Brain Res. 2001, 125, 279–284. [Google Scholar] [CrossRef] [Green Version]
- Thisse, C.; Thisse, B. High-resolution in situ hybridization to whole-mount zebrafish embryos. Nat. Protoc. 2008, 3, 59–69. [Google Scholar] [CrossRef]
- Fukuhara, S.; Fukui, H.; Wakayama, Y.; Ando, K.; Nakajima, H.; Mochizuki, N. Looking back and moving forward: Recent advances in understanding of cardiovascular development by imaging of zebrafish. Dev. Growth Differ. 2015, 57, 333–340. [Google Scholar] [CrossRef] [Green Version]
- Betz, C.; Lenard, A.; Belting, H.G.; Affolter, M. Cell behaviors and dynamics during angiogenesis. Development 2016, 143, 2249–2260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sumanas, S.; Jorniak, T.; Lin, S. Identification of novel vascular endothelial-specific genes by the microarray analysis of the zebrafish cloche mutants. Blood 2005, 106, 534–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez, G.A.; Veldman, M.B.; Zhao, Y.; Burgess, S.; Lin, S. Discovery and characterization of novel vascular and hematopoietic genes downstream of etsrp in zebrafish. PLoS ONE 2009, 4, e4994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, K.S.; Proulx, K.; Rost, M.S.; Sumanas, S. Identification of vasculature-specific genes by microarray analysis of Etsrp/Etv2 overexpressing zebrafish embryos. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2009, 238, 1836–1850. [Google Scholar] [CrossRef]
- Arnold, C.R.; Lamont, R.E.; Walker, J.T.; Spice, P.J.; Chan, C.K.; Ho, C.Y.; Childs, S.J. Comparative analysis of genes regulated by Dzip1/iguana and hedgehog in zebrafish. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2015, 244, 211–223. [Google Scholar] [CrossRef]
- Watanabe, M.; Yoshida, K.; Hida, M.; Kato, H.; Uchida, K.; Yamaguchi, R.; Tateyama, S.; Sugano, S. Cloning, expression analysis, and chromosomal mapping of GTPBP2, a novel member of the G protein family. Gene 2000, 256, 51–58. [Google Scholar] [CrossRef]
- Takai, Y.; Sasaki, T.; Matozaki, T. Small GTP-binding proteins. Physiol. Rev. 2001, 81, 153–208. [Google Scholar] [CrossRef] [PubMed]
- Lakshmikanthan, S.; Sobczak, M.; Chun, C.; Henschel, A.; Dargatz, J.; Ramchandran, R.; Chrzanowska-Wodnicka, M. Rap1 promotes VEGFR2 activation and angiogenesis by a mechanism involving integrin alphavbeta(3). Blood 2011, 118, 2015–2026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kather, J.N.; Kroll, J. Rho guanine exchange factors in blood vessels: Fine-tuners of angiogenesis and vascular function. Exp. Cell Res. 2013, 319, 1289–1297. [Google Scholar] [CrossRef] [PubMed]
- Chetty, C.; Lakka, S.S.; Bhoopathi, P.; Rao, J.S. MMP-2 alters VEGF expression via alphaVbeta3 integrin-mediated PI3K/AKT signaling in A549 lung cancer cells. Int. J. Cancer 2010, 127, 1081–1095. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.L.; Lin, S.W.; Fang, H.C.; Chou, K.J.; Bee, Y.S.; Chu, T.H.; Chang, M.C.; Weng, W.T.; Wu, C.Y.; Cho, C.L.; et al. A novel poly-naphthol compound ST104P suppresses angiogenesis by attenuating matrix metalloproteinase-2 expression in endothelial cells. Int. J. Mol. Sci. 2014, 15, 16611–16627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Uhlirova, M.; Bohmann, D. Spatial restriction of FGF signaling by a matrix metalloprotease controls branching morphogenesis. Dev Cell 2010, 18, 157–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potente, M.; Urbich, C.; Sasaki, K.; Hofmann, W.K.; Heeschen, C.; Aicher, A.; Kollipara, R.; DePinho, R.A.; Zeiher, A.M.; Dimmeler, S. Involvement of Foxo transcription factors in angiogenesis and postnatal neovascularization. J. Clin. Investig. 2005, 115, 2382–2392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.Y.; Fox, C.S.; North, T.E.; Goessling, W. Functional validation of GWAS gene candidates for abnormal liver function during zebrafish liver development. Dis. Models Mech. 2013, 6, 1271–1278. [Google Scholar] [CrossRef] [Green Version]
- Davis, D.A.; Singer, K.E.; De La Luz Sierra, M.; Narazaki, M.; Yang, F.; Fales, H.M.; Yarchoan, R.; Tosato, G. Identification of carboxypeptidase N as an enzyme responsible for C-terminal cleavage of stromal cell-derived factor-1alpha in the circulation. Blood 2005, 105, 4561–4568. [Google Scholar] [CrossRef]
- Weiss, A.; Baumgartner, M.; Radziwill, G.; Dennler, J.; Moelling, K. c-Src is a PDZ interaction partner and substrate of the E3 ubiquitin ligase Ligand-of-Numb protein X1. FEBS Lett. 2007, 581, 5131–5136. [Google Scholar] [CrossRef] [Green Version]
- Rehn, K.; Wong, K.S.; Balciunas, D.; Sumanas, S. Zebrafish enhancer trap line recapitulates embryonic aquaporin 1a expression pattern in vascular endothelial cells. Int. J. Dev. Biol. 2011, 55, 613–618. [Google Scholar] [CrossRef]
- Nicoli, S.; Knyphausen, C.P.; Zhu, L.J.; Lakshmanan, A.; Lawson, N.D. miR-221 is required for endothelial tip cell behaviors during vascular development. Dev. Cell 2012, 22, 418–429. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, R.-F.; Wang, Y.-S.; Lu, F.-I.; Huang, Y.-S.; Chiu, C.-C.; Tai, M.-H.; Wu, C.-Y. Identification of Novel Vascular Genes Downstream of Islet2 and Nr2f1b Transcription Factors. Biomedicines 2022, 10, 1261. https://doi.org/10.3390/biomedicines10061261
Li R-F, Wang Y-S, Lu F-I, Huang Y-S, Chiu C-C, Tai M-H, Wu C-Y. Identification of Novel Vascular Genes Downstream of Islet2 and Nr2f1b Transcription Factors. Biomedicines. 2022; 10(6):1261. https://doi.org/10.3390/biomedicines10061261
Chicago/Turabian StyleLi, Ru-Fang, Yi-Shan Wang, Fu-I Lu, Yi-Shan Huang, Chien-Chih Chiu, Ming-Hong Tai, and Chang-Yi Wu. 2022. "Identification of Novel Vascular Genes Downstream of Islet2 and Nr2f1b Transcription Factors" Biomedicines 10, no. 6: 1261. https://doi.org/10.3390/biomedicines10061261
APA StyleLi, R.-F., Wang, Y.-S., Lu, F.-I., Huang, Y.-S., Chiu, C.-C., Tai, M.-H., & Wu, C.-Y. (2022). Identification of Novel Vascular Genes Downstream of Islet2 and Nr2f1b Transcription Factors. Biomedicines, 10(6), 1261. https://doi.org/10.3390/biomedicines10061261