
Different Effects of Valproic Acid on SLC12A2, SLC12A5 and SLC5A8 Gene Expression in Pediatric Glioblastoma Cells as an Approach to Personalised Therapy


 , and
, and
Abstract
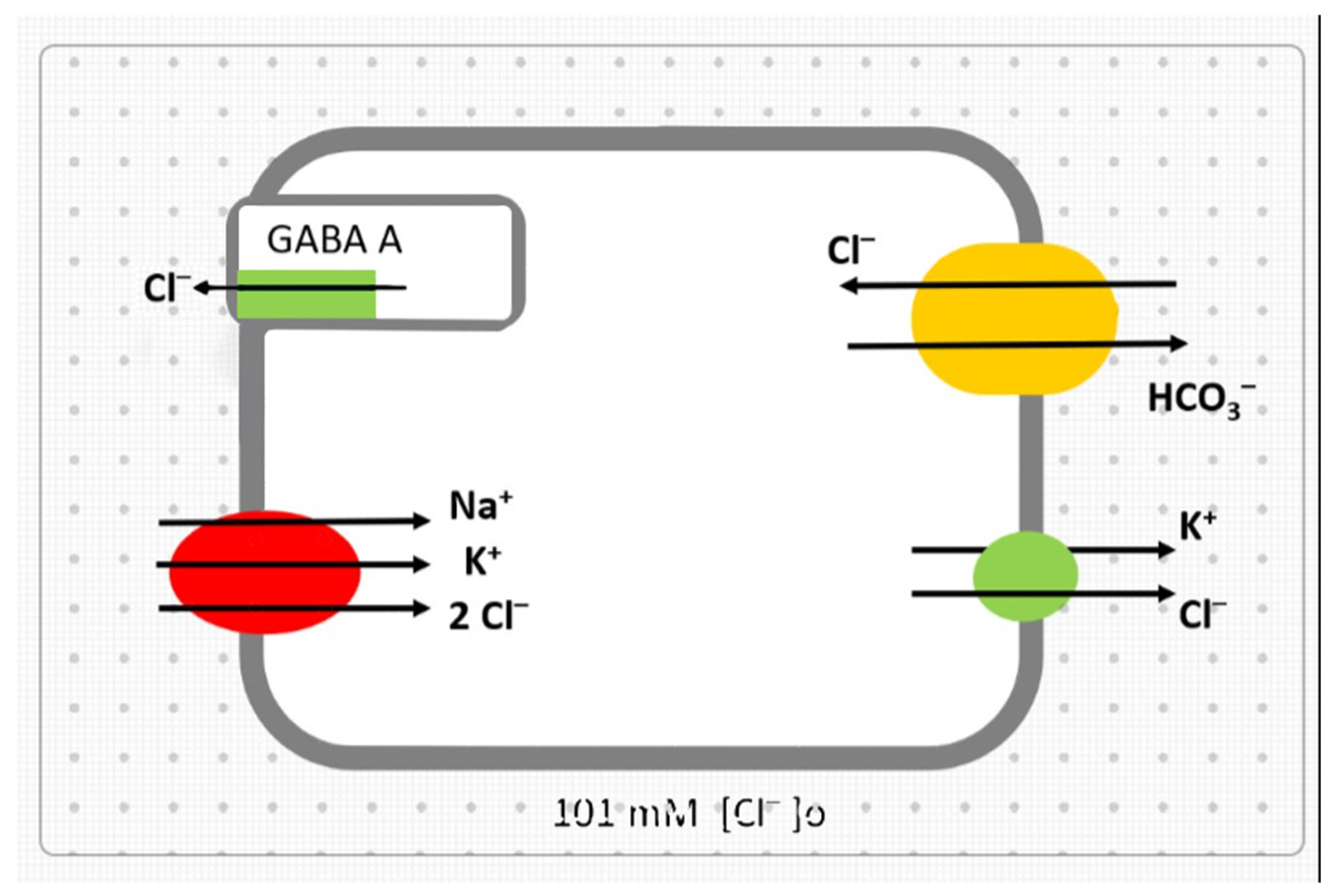
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Cell Culture
2.2. Extraction of RNA from PBT24 and SF8628 Cell Line Cells
2.3. Determination of the SLC12A2, SLC12A5 and SLC5A8 Expression in PBT24 and SF8628 Cell Line Cells
2.4. Statistical Analysis
3. Results
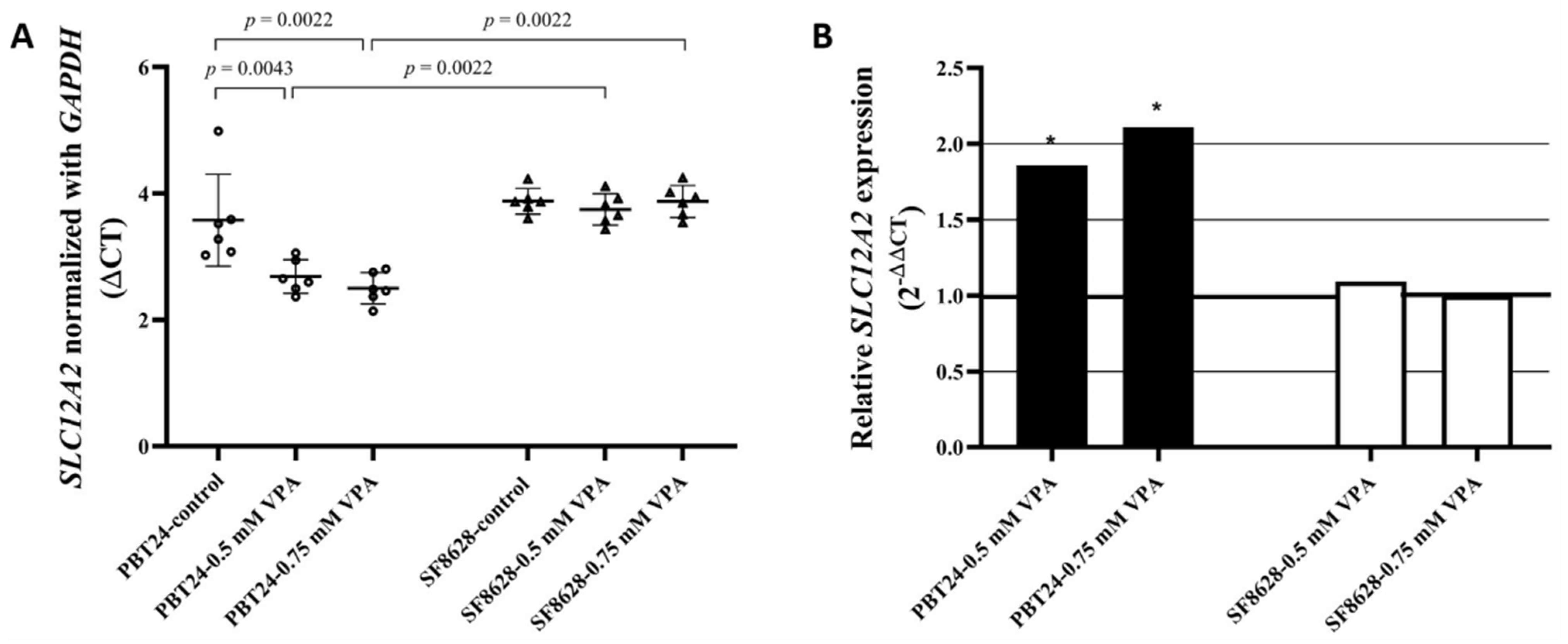
3.1. The Expression of SLC12A2 in PBT24 and SF8628 Cell Study Groups
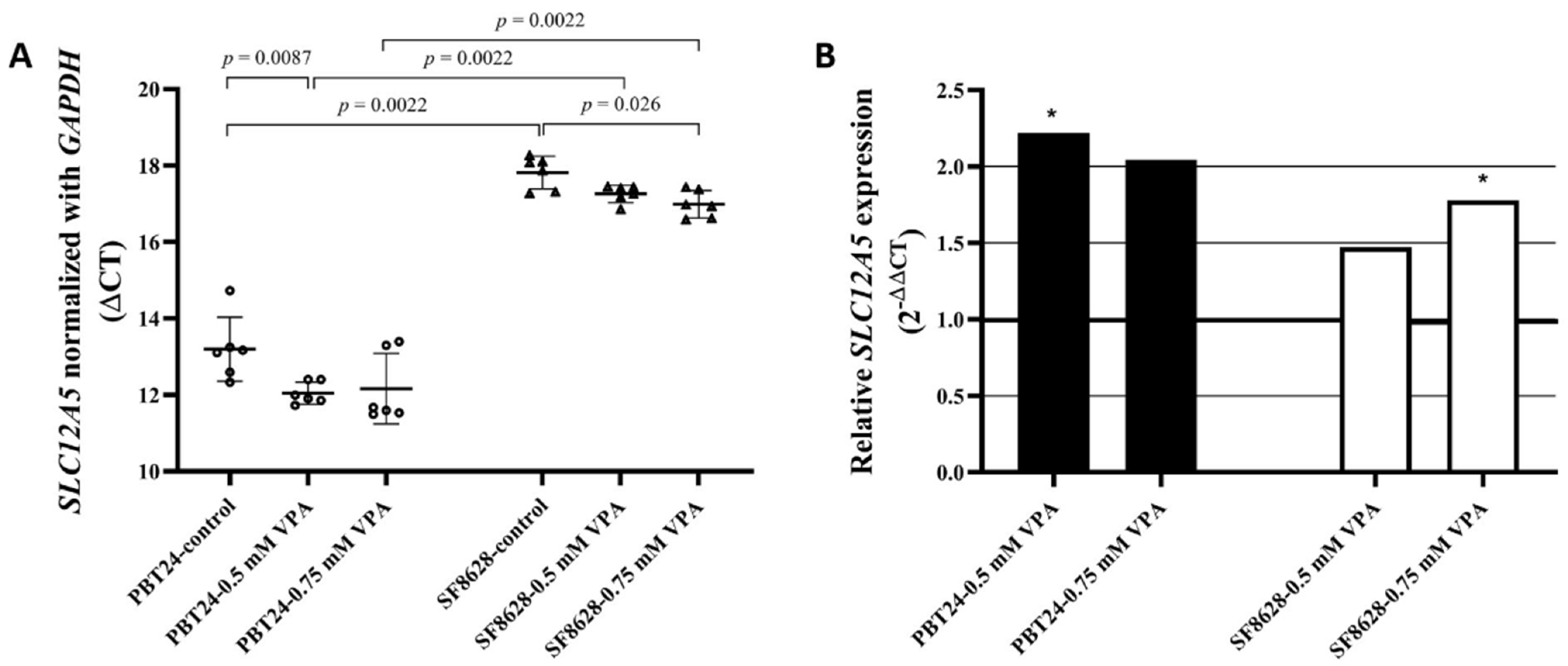
3.2. The Expression of SLC12A5 in PBT24 and SF8628 Cell Study Groups
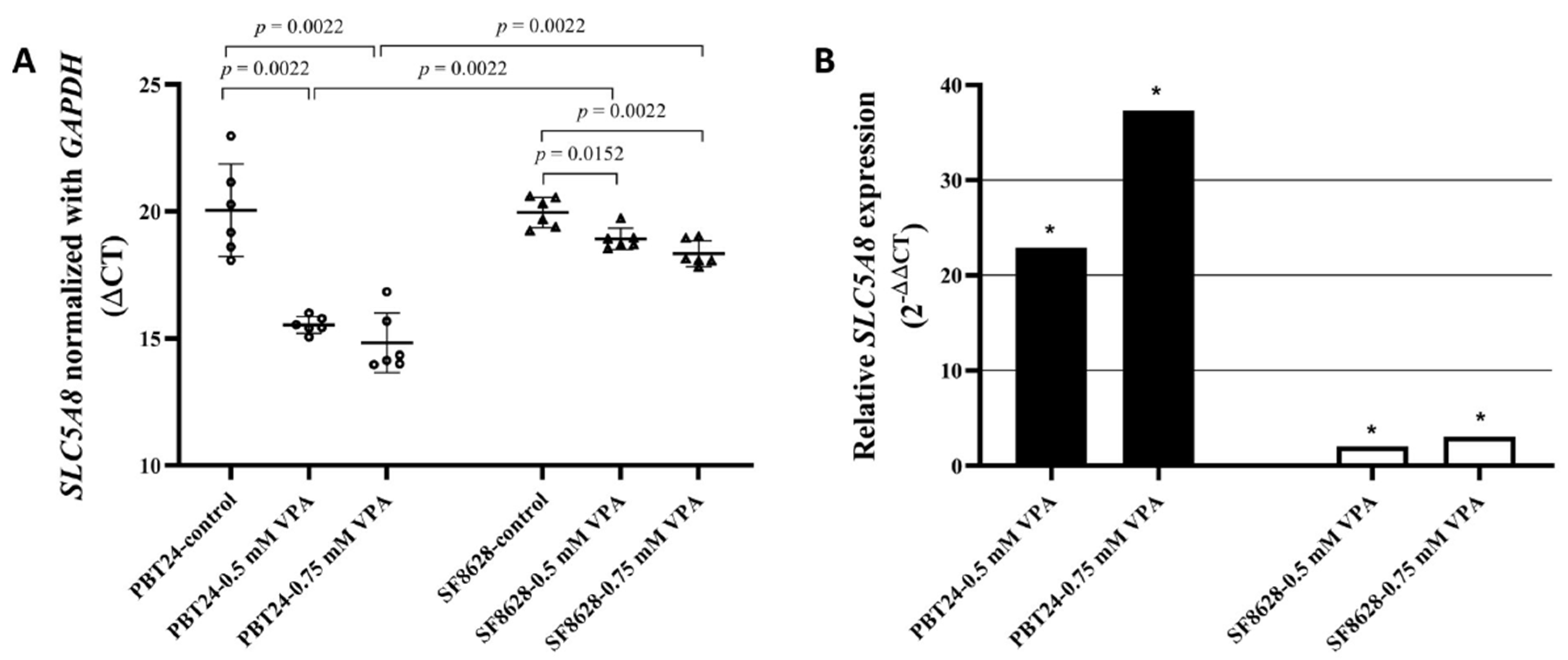
3.3. The Expression of SLC5A8 in PBT24 and SF8628 Cell Study Groups
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Njonkou, R.; Jackson, C.M.; Woodworth, G.F.; Hersh, D.S. Pediatric Glioblastoma: Mechanisms of Immune Evasion and Potential Therapeutic Opportunities. Cancer Immunol. Immunother. 2022. [Google Scholar] [CrossRef] [PubMed]
- Coleman, C.; Stoller, S.; Grotzer, M.; Stucklin, A.G.; Nazarian, J.; Mueller, S. Pediatric Hemispheric High-Grade Glioma: Targeting the Future. Cancer Metastasis Rev. 2020, 39, 245–260. [Google Scholar] [CrossRef]
- Jones, C.; Karajannis, M.A.; Jones, D.T.W.; Kieran, M.W.; Monje, M.; Baker, S.J.; Becher, O.J.; Cho, Y.-J.; Gupta, N.; Hawkins, C.; et al. Pediatric High-Grade Glioma: Biologically and Clinically in Need of New Thinking. Neuro Oncol. 2017, 19, 153–161. [Google Scholar] [CrossRef] [Green Version]
- Ene, C.I.; Holland, E.C. Personalized Medicine for Gliomas. Surg. Neurol. Int. 2015, 6, S89–S95. [Google Scholar] [CrossRef]
- Puiu, R.A.; Balaure, P.C.; Constantinescu, E.; Grumezescu, A.M.; Andronescu, E.; Oprea, O.-C.; Vasile, B.S.; Grumezescu, V.; Negut, I.; Nica, I.C.; et al. Anti-Cancer Nanopowders and MAPLE-Fabricated Thin Films Based on SPIONs Surface Modified with Paclitaxel Loaded β-Cyclodextrin. Pharmaceutics 2021, 13, 1356. [Google Scholar] [CrossRef] [PubMed]
- Fisher, J.L.; Jones, E.F.; Flanary, V.L.; Williams, A.S.; Ramsey, E.J.; Lasseigne, B.N. Considerations and Challenges for Sex-Aware Drug Repurposing. Biol. Sex Differ. 2022, 13, 13. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Chen, H.; Tan, Q.; Xie, C.; Zhan, W.; Sharma, A.; Sharma, H.S.; Zhang, Z. The Therapeutic and Neuroprotective Effects of an Antiepileptic Drug Valproic Acid in Glioma Patients. Prog. Brain Res. 2020, 258, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Daśko, M.; de Pascual-Teresa, B.; Ortín, I.; Ramos, A. HDAC Inhibitors: Innovative Strategies for Their Design and Applications. Molecules 2022, 27, 715. [Google Scholar] [CrossRef]
- Wawruszak, A.; Halasa, M.; Okon, E.; Kukula-Koch, W.; Stepulak, A. Valproic Acid and Breast Cancer: State of the Art in 2021. Cancers 2021, 13, 3409. [Google Scholar] [CrossRef]
- Monga, V.; Swami, U.; Tanas, M.; Bossler, A.; Mott, S.L.; Smith, B.J.; Milhem, M. A Phase I/II Study Targeting Angiogenesis Using Bevacizumab Combined with Chemotherapy and a Histone Deacetylase Inhibitor (Valproic Acid) in Advanced Sarcomas. Cancers 2018, 10, 53. [Google Scholar] [CrossRef] [Green Version]
- Han, W.; Yu, F.; Wang, R.; Guan, W.; Zhi, F. Valproic Acid Sensitizes Glioma Cells to Luteolin through Induction of Apoptosis and Autophagy via Akt Signaling. Cell. Mol. Neurobiol. 2021, 41, 1625–1634. [Google Scholar] [CrossRef] [PubMed]
- Shao, C.; Wu, M.; Chen, F.; Li, C.; Xia, Y.; Chen, Z. Histone Deacetylase Inhibitor, 2-Propylpentanoic Acid, Increases the Chemosensitivity and Radiosensitivity of Human Glioma Cell Lines in Vitro. Chin. Med. J. 2012, 125, 4338–4343. [Google Scholar] [PubMed]
- Chinnaiyan, P.; Cerna, D.; Burgan, W.E.; Beam, K.; Williams, E.S.; Camphausen, K.; Tofilon, P.J. Postradiation Sensitization of the Histone Deacetylase Inhibitor Valproic Acid. Clin. Cancer Res. 2008, 14, 5410–5415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Nifterik, K.A.; Van den Berg, J.; Slotman, B.J.; Lafleur, M.V.M.; Sminia, P.; Stalpers, L.J.A. Valproic Acid Sensitizes Human Glioma Cells for Temozolomide and γ-Radiation. J. Neuro-Oncol. 2012, 107, 61–67. [Google Scholar] [CrossRef]
- Pont, L.M.E.B.; Naipal, K.; Kloezeman, J.J.; Venkatesan, S.; van den Bent, M.; van Gent, D.C.; Dirven, C.M.F.; Kanaar, R.; Lamfers, M.L.M.; Leenstra, S. DNA Damage Response and Anti-Apoptotic Proteins Predict Radiosensitization Efficacy of HDAC Inhibitors SAHA and LBH589 in Patient-Derived Glioblastoma Cells. Cancer Lett. 2015, 356, 525–535. [Google Scholar] [CrossRef] [PubMed]
- Su, J.M.-F.; Murray, J.C.; McNall-Knapp, R.Y.; Bowers, D.C.; Shah, S.; Adesina, A.M.; Paulino, A.C.; Jo, E.; Mo, Q.; Baxter, P.A.; et al. A Phase 2 Study of Valproic Acid and Radiation, Followed by Maintenance Valproic Acid and Bevacizumab in Children with Newly Diagnosed Diffuse Intrinsic Pontine Glioma or High-Grade Glioma. Pediatr. Blood Cancer 2020, 67, e28283. [Google Scholar] [CrossRef]
- Berendsen, S.; Broekman, M.; Seute, T.; Snijders, T.; van Es, C.; de Vos, F.; Regli, L.; Robe, P. Valproic Acid for the Treatment of Malignant Gliomas: Review of the Preclinical Rationale and Published Clinical Results. Expert Opin. Investig. Drugs 2012, 21, 1391–1415. [Google Scholar] [CrossRef]
- Rost, D.; Kopplow, K.; Gehrke, S.; Mueller, S.; Friess, H.; Ittrich, C.; Mayer, D.; Stiehl, A. Gender-Specific Expression of Liver Organic Anion Transporters in Rat. Eur. J. Clin. Investig. 2005, 35, 635–643. [Google Scholar] [CrossRef]
- Smirnova, O.V. Sex differences in drug action: The role of multidrug-resistance proteins (MRPs). Fiziol Cheloveka 2012, 38, 124–136. [Google Scholar]
- Suzuki, T.; Zhao, Y.L.; Nadai, M.; Naruhashi, K.; Shimizu, A.; Takagi, K.; Takagi, K.; Hasegawa, T. Gender-Related Differences in Expression and Function of Hepatic P-Glycoprotein and Multidrug Resistance-Associated Protein (Mrp2) in Rats. Life Sci. 2006, 79, 455–461. [Google Scholar] [CrossRef]
- Ibarra, M.; Vázquez, M.; Fagiolino, P.; Derendorf, H. Sex Related Differences on Valproic Acid Pharmacokinetics after Oral Single Dose. J. Pharmacokinet. Pharmacodyn. 2013, 40, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Wong, H.; Xia, B.; Tong, V.; Kumar, S.; Kenny, J. Atypical Kinetics of Valproic Acid Glucuronidation In vitro and In vivo in Humans. Biol. Med. 2012, S11, 6. [Google Scholar] [CrossRef] [Green Version]
- Riva, G.; Cilibrasi, C.; Bazzoni, R.; Cadamuro, M.; Negroni, C.; Butta, V.; Strazzabosco, M.; Dalprà, L.; Lavitrano, M.; Bentivegna, A. Valproic Acid Inhibits Proliferation and Reduces Invasiveness in Glioma Stem Cells Through Wnt/β Catenin Signalling Activation. Genes 2018, 9, 522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okada, Y.; Maeno, E. Apoptosis, Cell Volume Regulation and Volume-Regulatory Chloride Channels. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2001, 130, 377–383. [Google Scholar] [CrossRef]
- Maeno, E.; Ishizaki, Y.; Kanaseki, T.; Hazama, A.; Okada, Y. Normotonic Cell Shrinkage Because of Disordered Volume Regulation is an Early Prerequisite to Apoptosis. Proc. Natl. Acad. Sci. USA 2000, 97, 9487–9492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bortner, C.D.; Cidlowski, J.A. Cell Shrinkage and Monovalent Cation Fluxes: Role in Apoptosis. Arch. Biochem. Biophys. 2007, 462, 176–188. [Google Scholar] [CrossRef] [Green Version]
- Garzon-Muvdi, T.; Schiapparelli, P.; ap Rhys, C.; Guerrero-Cazares, H.; Smith, C.; Kim, D.-H.; Kone, L.; Farber, H.; Lee, D.Y.; An, S.S.; et al. Regulation of Brain Tumor Dispersal by NKCC1 through a Novel Role in Focal Adhesion Regulation. PLoS Biol. 2012, 10, e1001320. [Google Scholar] [CrossRef] [Green Version]
- Pallud, J.; Le Van Quyen, M.; Bielle, F.; Pellegrino, C.; Varlet, P.; Cresto, N.; Baulac, M.; Duyckaerts, C.; Kourdougli, N.; Chazal, G.; et al. Cortical GABAergic Excitation Contributes to Epileptic Activities around Human Glioma. Sci. Transl. Med. 2014, 6, 244ra89. [Google Scholar] [CrossRef] [Green Version]
- Aronica, E.; Boer, K.; Redeker, S.; Spliet, W.G.M.; van Rijen, P.C.; Troost, D.; Gorter, J.A. Differential Expression Patterns of Chloride Transporters, Na+-K+-2Cl--Cotransporter and K+-Cl--Cotransporter, in Epilepsy-Associated Malformations of Cortical Development. Neuroscience 2007, 145, 185–196. [Google Scholar] [CrossRef]
- Thangaraju, M.; Carswell, K.N.; Prasad, P.D.; Ganapathy, V. Colon Cancer Cells Maintain Low Levels of Pyruvate to Avoid Cell Death Caused by Inhibition of HDAC1/HDAC3. Biochem. J. 2009, 417, 379–389. [Google Scholar] [CrossRef]
- Cong, D.; Zhu, W.; Kuo, J.S.; Hu, S.; Sun, D. Ion Transporters in Brain Tumors. Curr. Med. Chem. 2015, 22, 1171–1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, B.R.; Cuddapah, V.A.; Watkins, S.; Rohn, K.J.; Dy, T.E.; Sontheimer, H. With-No-Lysine Kinase 3 (WNK3) Stimulates Glioma Invasion by Regulating Cell Volume. Am. J. Physiol. Cell Physiol. 2011, 301, C1150–C1160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, B.R.; Sontheimer, H. Inhibition of the Sodium-Potassium-Chloride Cotransporter Isoform-1 Reduces Glioma Invasion. Cancer Res. 2010, 70, 5597–5606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juknevičienė, M.; Balnytė, I.; Valančiūtė, A.; Lesauskaitė, V.; Stanevičiūtė, J.; Curkūnavičiūtė, R.; Stakišaitis, D. Valproic Acid Inhibits NA-K-2CL Cotransporter RNA Expression in Male But Not in Female Rat Thymocytes. Dose Response 2019, 17, 1559325819852444. [Google Scholar] [CrossRef] [PubMed]
- Hong, C.; Maunakea, A.; Jun, P.; Bollen, A.W.; Hodgson, J.G.; Goldenberg, D.D.; Weiss, W.A.; Costello, J.F. Shared Epigenetic Mechanisms in Human and Mouse Gliomas Inactivate Expression of the Growth Suppressor SLC5A8. Cancer Res. 2005, 65, 3617–3623. [Google Scholar] [CrossRef] [Green Version]
- Gopal, E.; Fei, Y.-J.; Miyauchi, S.; Zhuang, L.; Prasad, P.D.; Ganapathy, V. Sodium-Coupled and Electrogenic Transport of B-Complex Vitamin Nicotinic Acid by Slc5a8, a Member of the Na/Glucose Co-Transporter Gene Family. Biochem. J. 2005, 388, 309–316. [Google Scholar] [CrossRef] [Green Version]
- Babu, E.; Ramachandran, S.; CoothanKandaswamy, V.; Elangovan, S.; Prasad, P.D.; Ganapathy, V.; Thangaraju, M. Role of SLC5A8, a Plasma Membrane Transporter and a Tumor Suppressor, in the Antitumor Activity of Dichloroacetate. Oncogene 2011, 30, 4026–4037. [Google Scholar] [CrossRef] [Green Version]
- Travaglini, L.; Vian, L.; Billi, M.; Grignani, F.; Nervi, C. Epigenetic Reprogramming of Breast Cancer Cells by Valproic Acid Occurs Regardless of Estrogen Receptor Status. Int. J. Biochem. Cell Biol. 2009, 41, 225–234. [Google Scholar] [CrossRef]
- Milutinovic, S.; D’Alessio, A.C.; Detich, N.; Szyf, M. Valproate Induces Widespread Epigenetic Reprogramming Which Involves Demethylation of Specific Genes. Carcinogenesis 2007, 28, 560–571. [Google Scholar] [CrossRef] [Green Version]
- Veronezi, G.M.B.; Felisbino, M.B.; Gatti, M.S.V.; Mello, M.L.S.; de Vidal, B.C. DNA Methylation Changes in Valproic Acid-Treated HeLa Cells as Assessed by Image Analysis, Immunofluorescence and Vibrational Microspectroscopy. PLoS ONE 2017, 12, e0170740. [Google Scholar] [CrossRef]
- Kaji, I.; Iwanaga, T.; Watanabe, M.; Guth, P.H.; Engel, E.; Kaunitz, J.D.; Akiba, Y. SCFA Transport in Rat Duodenum. Am. J. Physiol. Gastrointest Liver Physiol. 2015, 308, G188–G197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omer, S.; Koumangoye, R.; Delpire, E. A Mutation in the Na-K-2Cl Cotransporter-1 Leads to Changes in Cellular Metabolism. J. Cell. Physiol. 2020, 235, 7239–7250. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Velez, N.; Marigil, M.; García-Moure, M.; Gonzalez-Huarriz, M.; Aristu, J.J.; Ramos-García, L.-I.; Tejada, S.; Díez-Valle, R.; Patiño-García, A.; Becher, O.J.; et al. Delta-24-RGD Combined with Radiotherapy Exerts a Potent Antitumor Effect in Diffuse Intrinsic Pontine Glioma and Pediatric High Grade Glioma Models. Acta Neuropathol. Commun. 2019, 7, 64. [Google Scholar] [CrossRef] [Green Version]
- Sigma-Aldrich. SF8628 Human DIPG H3.3-K27M Cell Line SF8628 Pediatric Diffuse Intrinsic Pontine Glioma (DIPG) Cell Line Harbors the Histone H3.3 Lys 27-to-Methionine (K27M) Mutation and Can Support Research and Drug Development Efforts Targeting DIPG. Available online: http://www.sigmaaldrich.com/ (accessed on 8 December 2021).
- Mueller, S.; Hashizume, R.; Yang, X.; Kolkowitz, I.; Olow, A.K.; Phillips, J.; Smirnov, I.; Tom, M.W.; Prados, M.D.; James, C.D.; et al. Targeting Wee1 for the Treatment of Pediatric High-Grade Gliomas. Neuro Oncol. 2014, 16, 352–360. [Google Scholar] [CrossRef] [PubMed]
- Coulter, D.W.; Walko, C.; Patel, J.; Moats-Staats, B.M.; McFadden, A.; Smith, S.V.; Khan, W.A.; Bridges, A.S.; Deal, A.M.; Oesterheld, J.; et al. Valproic Acid Reduces the Tolerability of Temsirolimus in Children with Solid Tumors. Anticancer Drugs 2013, 24, 415–421. [Google Scholar] [CrossRef] [Green Version]
- Mei, S.; Feng, W.; Zhu, L.; Yu, Y.; Yang, W.; Gao, B.; Wu, X.; Zhao, Z.; Fang, F. Genetic Polymorphisms and Valproic Acid Plasma Concentration in Children with Epilepsy on Valproic Acid Monotherapy. Seizure-Eur. J. Epilepsy 2017, 51, 22–26. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Torrisi, F.; Alberghina, C.; D’Aprile, S.; Pavone, A.M.; Longhitano, L.; Giallongo, S.; Tibullo, D.; Di Rosa, M.; Zappalà, A.; Cammarata, F.P.; et al. The Hallmarks of Glioblastoma: Heterogeneity, Intercellular Crosstalk and Molecular Signature of Invasiveness and Progression. Biomedicines 2022, 10, 806. [Google Scholar] [CrossRef]
- Damanskienė, E.; Balnytė, I.; Valančiūtė, A.; Alonso, M.M.; Preikšaitis, A.; Stakišaitis, D. The Different Temozolomide Effects on Tumorigenesis Mechanisms of Pediatric Glioblastoma PBT24 and SF8628 Cell Tumor in CAM Model and on Cells In Vitro. Int. J. Mol. Sci. 2022, 23, 2001. [Google Scholar] [CrossRef]
- Algharabil, J.; Kintner, D.B.; Wang, Q.; Begum, G.; Clark, P.A.; Yang, S.-S.; Lin, S.-H.; Kahle, K.T.; Kuo, J.S.; Sun, D. Inhibition of Na(+)-K(+)-2Cl(-) Cotransporter Isoform 1 Accelerates Temozolomide-Mediated Apoptosis in Glioblastoma Cancer Cells. Cell. Physiol. Biochem. 2012, 30, 33–48. [Google Scholar] [CrossRef]
- Gagnon, K.B.E.; England, R.; Delpire, E. Characterization of SPAK and OSR1, Regulatory Kinases of the Na-K-2Cl Cotransporter. Mol. Cell. Biol. 2006, 26, 689–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahle, K.T.; Rinehart, J.; Lifton, R.P. Phosphoregulation of the Na-K-2Cl and K-Cl Cotransporters by the WNK Kinases. Biochim. Biophys. Acta 2010, 1802, 1150–1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bortner, C.D.; Sifre, M.I.; Cidlowski, J.A. Cationic Gradient Reversal and Cytoskeleton-Independent Volume Regulatory Pathways Define an Early Stage of Apoptosis. J. Biol. Chem. 2008, 283, 7219–7229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bortner, C.D.; Hughes, F.M.; Cidlowski, J.A. A Primary Role for K+ and Na+ Efflux in the Activation of Apoptosis. J. Biol. Chem. 1997, 272, 32436–32442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, J.; Okabe, A.; Toyoda, H.; Kilb, W.; Luhmann, H.J.; Fukuda, A. Cl- Uptake Promoting Depolarizing GABA Actions in Immature Rat Neocortical Neurones Is Mediated by NKCC1. J. Physiol. 2004, 557, 829–841. [Google Scholar] [CrossRef]
- Chen, H.; Sun, D. The Role of Na-K-Cl Co-Transporter in Cerebral Ischemia. Neurol. Res. 2005, 27, 280–286. [Google Scholar] [CrossRef]
- Ben-Ari, Y. Excitatory Actions of Gaba during Development: The Nature of the Nurture. Nat. Rev. Neurosci. 2002, 3, 728–739. [Google Scholar] [CrossRef]
- Owens, D.F.; Kriegstein, A.R. Is There More to GABA than Synaptic Inhibition? Nat. Rev. Neurosci. 2002, 3, 715–727. [Google Scholar] [CrossRef]
- Di Cristo, G.; Awad, P.N.; Hamidi, S.; Avoli, M. KCC2, Epileptiform Synchronization, and Epileptic Disorders. Prog. Neurobiol. 2018, 162, 1–16. [Google Scholar] [CrossRef]
- Su, J.M.; Li, X.-N.; Thompson, P.; Ou, C.-N.; Ingle, A.M.; Russell, H.; Lau, C.C.; Adamson, P.C.; Blaney, S.M. Phase 1 Study of Valproic Acid in Pediatric Patients with Refractory Solid or CNS Tumors: A Children’s Oncology Group Report. Clin. Cancer Res. 2011, 17, 589–597. [Google Scholar] [CrossRef] [Green Version]
- Wolff, J.E.A.; Kramm, C.; Kortmann, R.-D.; Pietsch, T.; Rutkowski, S.; Jorch, N.; Gnekow, A.; Driever, P.H. Valproic Acid Was Well Tolerated in Heavily Pretreated Pediatric Patients with High-Grade Glioma. J. Neuro-Oncol. 2008, 90, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Witt, O.; Schweigerer, L.; Driever, P.H.; Wolff, J.; Pekrun, A. Valproic Acid Treatment of Glioblastoma Multiforme in a Child. Pediatr. Blood Cancer 2004, 43, 181. [Google Scholar] [CrossRef] [PubMed]
- Happold, C.; Gorlia, T.; Chinot, O.; Gilbert, M.; Nabors, L.B.; Wick, W.; Pugh, S.L.; Hegi, M.; Cloughesy, T.; Roth, P.; et al. 26LBA Does Valproic Acid Improve Survival in Glioblastoma? A Meta-Analysis of Randomized Trials in Newly Diagnosed Glioblastoma. Eur. J. Cancer 2015, 51, S723. [Google Scholar] [CrossRef]
- Thangaraju, M.; Ananth, S.; Martin, P.M.; Roon, P.; Smith, S.B.; Sterneck, E.; Prasad, P.D.; Ganapathy, V. C/Ebpdelta Null Mouse as a Model for the Double Knock-out of Slc5a8 and Slc5a12 in Kidney. J. Biol. Chem. 2006, 281, 26769–26773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frank, H.; Gröger, N.; Diener, M.; Becker, C.; Braun, T.; Boettger, T. Lactaturia and Loss of Sodium-Dependent Lactate Uptake in the Colon of SLC5A8-Deficient Mice. J. Biol. Chem. 2008, 283, 24729–24737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mihaylova, M.M.; Vasquez, D.S.; Ravnskjaer, K.; Denechaud, P.-D.; Yu, R.T.; Alvarez, J.G.; Downes, M.; Evans, R.M.; Montminy, M.; Shaw, R.J. Class IIa Histone Deacetylases Are Hormone-Activated Regulators of FOXO and Mammalian Glucose Homeostasis. Cell 2011, 145, 607–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.; Warrington, N.M.; Taylor, S.J.; Whitmire, P.; Carrasco, E.; Singleton, K.W.; Wu, N.; Lathia, J.D.; Berens, M.E.; Kim, A.H.; et al. Sex Differences in GBM Revealed by Analysis of Patient Imaging, Transcriptome, and Survival Data. Sci. Transl. Med. 2019, 11, eaao5253. [Google Scholar] [CrossRef] [Green Version]
- Learn Science at Scitable. The Role of Methylation in Gene Expression. Available online: https://www.nature.com/scitable/topicpage/the-role-of-methylation-in-gene-expression-1070/ (accessed on 19 April 2022).
- de Sousa Abreu, R.; Penalva, L.O.; Marcotte, E.M.; Vogel, C. Global Signatures of Protein and MRNA Expression Levels. Mol. Biosyst. 2009, 5, 1512–1526. [Google Scholar] [CrossRef] [Green Version]
- Vogel, C.; Marcotte, E.M. Insights into the Regulation of Protein Abundance from Proteomic and Transcriptomic Analyses. Nat. Rev. Genet. 2012, 13, 227–232. [Google Scholar] [CrossRef]
- Koussounadis, A.; Langdon, S.P.; Um, I.H.; Harrison, D.J.; Smith, V.A. Relationship between Differentially Expressed MRNA and MRNA-Protein Correlations in a Xenograft Model System. Sci. Rep. 2015, 5, 10775. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Group | n | CT Mean | ΔCT Mean ± SD | ΔΔCT | |
|---|---|---|---|---|---|
| SLC12A2 | GAPDH | ||||
| PBT24-control | 6 | 22.951 | 19.372 | 3.579 ± 0.73 | |
| 0.5-mM VPA | 6 | 21.934 | 19.249 | 2.685 ± 0.27 a | −0.894 |
| 0.75-mM VPA | 6 | 21.847 | 19.346 | 2.501 ± 0.25 b | −1.077 |
| SF8628-control | 6 | 22.894 | 19.017 | 3.876 ± 0.21 | |
| 0.5-mM VPA | 6 | 23.340 | 19.592 | 3.748 ± 0.25 c | −0.128 |
| 0.75-mM VPA | 6 | 23.584 | 19.709 | 3.875 ± 0.25 d | −0.002 |
| Study Group | n | CT Mean | ΔCT Mean ± SD | ΔΔCT | |
|---|---|---|---|---|---|
| SLC12A5 | GAPDH | ||||
| PBT24-control | 6 | 32.564 | 19.372 | 13.191 ± 0.83 | |
| 0.5-mM VPA | 6 | 31.290 | 19.249 | 12.041 ± 0.29 a | −1.150 |
| 0.75-mM VPA | 6 | 31.505 | 19.346 | 12.159 ± 0.92 | −1.032 |
| SF8628-control | 6 | 36.831 | 19.017 | 17.814 ± 0.43 b | |
| 0.5-mM VPA | 6 | 36.848 | 19.592 | 17.256 ± 0.23 c | −0.558 |
| 0.75-mM VPA | 6 | 36.691 | 19.709 | 16.982 ± 0.36 d,e | −0.832 |
| Study Group | n | CT Mean | ΔCT Mean ± SD | ΔΔCT | |
|---|---|---|---|---|---|
| SLC5A8 | GAPDH | ||||
| PBT24-control | 6 | 39.416 | 19.372 | 20.044 ± 1.82 | |
| 0.5-mM VPA | 6 | 34.775 | 19.249 | 15.526 ± 0.33 a | −4.518 |
| 0.75-mM VPA | 6 | 34.170 | 19.346 | 14.824 ± 1.17 b | −5.220 |
| SF8628-control | 6 | 38.972 | 19.017 | 19.955 ± 0.59 | |
| 0.5-mM VPA | 6 | 38.507 | 19.592 | 18.915 ± 0.42 c,d | −1.040 |
| 0.75-mM VPA | 6 | 38.044 | 19.709 | 18.335 ± 0.51 e,f | −1.620 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Damanskienė, E.; Balnytė, I.; Valančiūtė, A.; Alonso, M.M.; Stakišaitis, D. Different Effects of Valproic Acid on SLC12A2, SLC12A5 and SLC5A8 Gene Expression in Pediatric Glioblastoma Cells as an Approach to Personalised Therapy. Biomedicines 2022, 10, 968. https://doi.org/10.3390/biomedicines10050968
Damanskienė E, Balnytė I, Valančiūtė A, Alonso MM, Stakišaitis D. Different Effects of Valproic Acid on SLC12A2, SLC12A5 and SLC5A8 Gene Expression in Pediatric Glioblastoma Cells as an Approach to Personalised Therapy. Biomedicines. 2022; 10(5):968. https://doi.org/10.3390/biomedicines10050968
Chicago/Turabian StyleDamanskienė, Eligija, Ingrida Balnytė, Angelija Valančiūtė, Marta Marija Alonso, and Donatas Stakišaitis. 2022. "Different Effects of Valproic Acid on SLC12A2, SLC12A5 and SLC5A8 Gene Expression in Pediatric Glioblastoma Cells as an Approach to Personalised Therapy" Biomedicines 10, no. 5: 968. https://doi.org/10.3390/biomedicines10050968
APA StyleDamanskienė, E., Balnytė, I., Valančiūtė, A., Alonso, M. M., & Stakišaitis, D. (2022). Different Effects of Valproic Acid on SLC12A2, SLC12A5 and SLC5A8 Gene Expression in Pediatric Glioblastoma Cells as an Approach to Personalised Therapy. Biomedicines, 10(5), 968. https://doi.org/10.3390/biomedicines10050968