
Breast Milk from Non-Obese Women with a High Omega-6 to Omega-3 Fatty Acid Ratio, but Not from Women with Obesity, Increases Lipogenic Gene Expression in 3T3-L1 Preadipocytes, Suggesting Adipocyte Dysfunction

 , , , ,
, , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Collection of BM Samples
2.3. Preparation of BM Whey
2.4. Fatty Acid Analysis of BM Samples
2.5. Treatment of 3T3-L1 Preadipocytes with BM
2.6. Oil Red O Staining
2.7. RNA Extraction and Real-Time Quantitative Polymerase Chain Reaction (qPCR)
2.8. Statistical Analysis
3. Results
3.1. Participant Characteristics
3.2. Effects of Maternal Obesity on the Fatty Acid Composition of BM
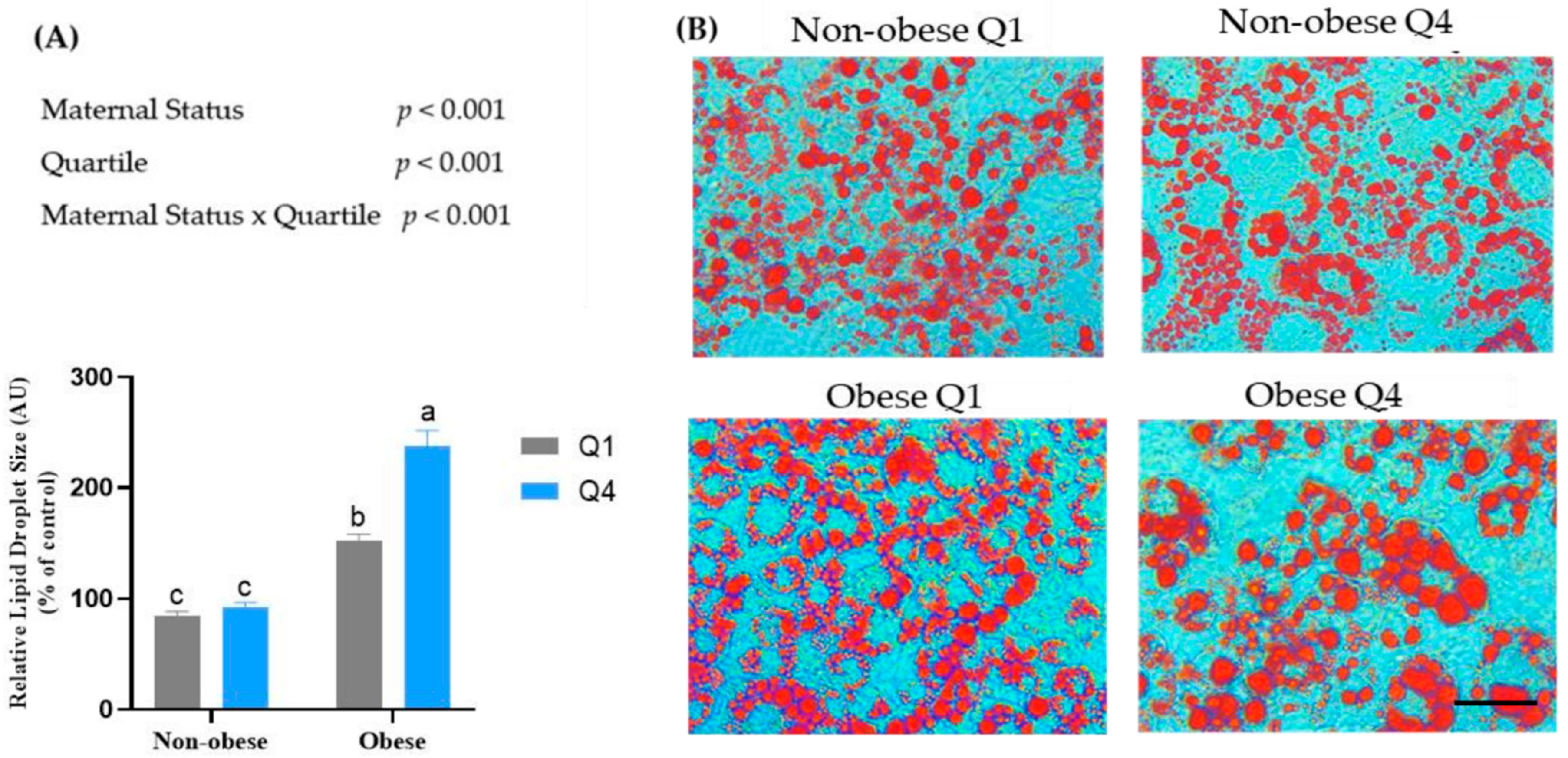
3.3. Maternal BMI and BM n-6:n-3 PUFA Ratio Affects Lipid Accumulation in Mature 3T3-L1 Adipocytes
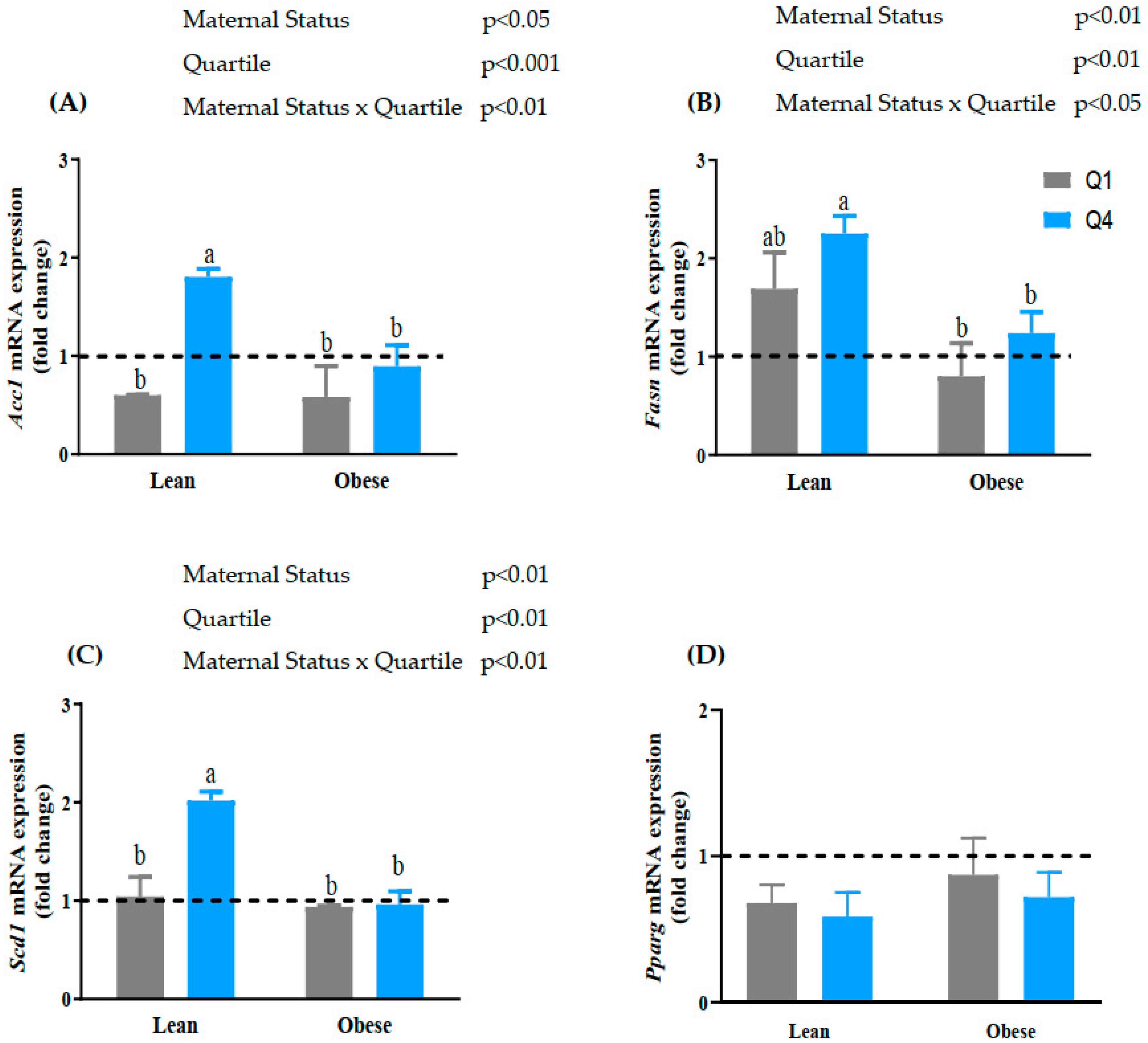
3.4. Maternal BMI and BM n-6:n-3 PUFA Ratio Alters the mRNA Expression of Lipogenic Genes
3.5. Maternal BMI and BM n-6:n-3 PUFA Ratio Had No Significant Effect on the mRNA Expression of Lipolytic Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Blüher, M. Obesity: Global epidemiology and pathogenesis. Nat. Rev. Endocrinol. 2019, 15, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.K.; DiBari, J.N. Marked disparities in pre-pregnancy obesity and overweight prevalence among US women by race/ethnicity, nativity/immigrant status, and sociodemographic characteristics, 2012–2014. J. Obes. 2019, 2019, 2419263. [Google Scholar] [CrossRef] [PubMed]
- Stubert, J.; Reister, F.; Hartmann, S.; Janni, W. The risks associated with obesity in pregnancy. Dtsch. Ärzteblatt Int. 2018, 115, 276. [Google Scholar] [CrossRef] [PubMed]
- Gaillard, R.; Felix, J.F.; Duijts, L.; Jaddoe, V.W. Childhood consequences of maternal obesity and excessive weight gain during pregnancy. Acta Obs. Gynecol. Scand. 2014, 93, 1085–1089. [Google Scholar] [CrossRef] [PubMed]
- Bernt, K.; Walker, W. Human milk as a carrier of biochemical messages. Acta Paediatr. 1999, 88, 27–41. [Google Scholar] [CrossRef]
- Pérez-Escamilla, R.; Buccini, G.S.; Segura-Pérez, S.; Piwoz, E. Perspective: Should exclusive breastfeeding still be recommended for 6 months? Adv. Nutr. 2019, 10, 931–943. [Google Scholar] [CrossRef]
- Koletzko, B. Human milk lipids. Ann. Nutr. Metab. 2016, 69, 27–40. [Google Scholar] [CrossRef]
- Innes, J.K.; Calder, P.C. Omega-6 fatty acids and inflammation. Prostaglandins Leukot. Essent. Fat. Acids 2018, 132, 41–48. [Google Scholar] [CrossRef]
- Wang, Y.; Huang, F. N-3 polyunsaturated fatty acids and inflammation in obesity: Local effect and systemic benefit. Biomed. Res. Int. 2015, 2015, 581469. [Google Scholar] [CrossRef]
- Yang, L.G.; Song, Z.X.; Yin, H.; Wang, Y.Y.; Shu, G.F.; Lu, H.X.; Wang, S.K.; Sun, G.J. Low n-6/n-3 PUFA ratio improves lipid metabolism, inflammation, oxidative stress and endothelial function in rats using plant oils as n-3 fatty acid source. Lipids 2016, 51, 49–59. [Google Scholar] [CrossRef]
- Vaidya, H.; Cheema, S. Breastmilk with a high omega-6 to omega-3 fatty acid ratio induced cellular events similar to insulin resistance and obesity in 3T3-L1 adipocytes. Pediatrics 2018, 13, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Enstad, S.; Cheema, S.; Thomas, R.; Fichorova, R.N.; Martin, C.R.; O’Tierney-Ginn, P.; Wagner, C.L.; Sen, S. The impact of maternal obesity and breast milk inflammation on developmental programming of infant growth. Eur. J. Clin. Nutr. 2021, 75, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Whitaker, K.M.; Marino, R.C.; Haapala, J.L.; Foster, L.; Smith, K.D.; Teague, A.M.; Jacobs, D.R.; Fontaine, P.L.; McGovern, P.M.; Schoenfuss, T.C. Associations of maternal weight status before, during, and after pregnancy with inflammatory markers in breast milk. Obesity 2017, 25, 2092–2099. [Google Scholar] [CrossRef]
- Collado, M.C.; Laitinen, K.; Salminen, S.; Isolauri, E. Maternal weight and excessive weight gain during pregnancy modify the immunomodulatory potential of breast milk. Pediatr. Res. 2012, 72, 77–85. [Google Scholar] [CrossRef]
- Andreas, N.J.; Hyde, M.J.; Herbert, B.R.; Jeffries, S.; Santhakumaran, S.; Mandalia, S.; Holmes, E.; Modi, N. Impact of maternal BMI and sampling strategy on the concentration of leptin, insulin, ghrelin and resistin in breast milk across a single feed: A longitudinal cohort study. BMJ Open 2016, 6, e010778. [Google Scholar] [CrossRef] [PubMed]
- Riera-Heredia, N.; Lutfi, E.; Gutiérrez, J.; Navarro, I.; Capilla, E. Fatty acids from fish or vegetable oils promote the adipogenic fate of mesenchymal stem cells derived from gilthead sea bream bone potentially through different pathways. PLoS ONE 2019, 14, e0215926. [Google Scholar] [CrossRef] [PubMed]
- Hollis, B.W.; Wagner, C.L.; Howard, C.R.; Ebeling, M.; Shary, J.R.; Smith, P.G.; Taylor, S.N.; Morella, K.; Lawrence, R.A.; Hulsey, T.C. Maternal versus infant vitamin D supplementation during lactation: A randomized controlled trial. Pediatrics 2015, 136, 625–634. [Google Scholar] [CrossRef]
- Yuksel, S.; Yigit, A.A.; Cinar, M.; Atmaca, N.; Onaran, Y. Oxidant and antioxidant status of human breast milk during lactation period. Dairy Sci. Technol. 2015, 95, 295–302. [Google Scholar] [CrossRef]
- Prieto Vidal, N.; Adeseun Adigun, O.; Pham, T.H.; Mumtaz, A.; Manful, C.; Callahan, G.; Stewart, P.; Keough, D.; Thomas, R.H. The effects of cold saponification on the unsaponified fatty acid composition and sensory perception of commercial natural herbal soaps. Molecules 2018, 23, 2356. [Google Scholar] [CrossRef]
- Lyle, R.E.; Corley, J.D.; McGehe, R.E. Human milk and infant formula can induce in vitro adipocyte differentiation in murine 3T3-L1 preadipocytes. Pediatr. Res. 1998, 44, 798–803. [Google Scholar] [CrossRef][Green Version]
- Fujisawa, Y.; Yamaguchi, R.; Nagata, E.; Satake, E.; Sano, S.; Matsushita, R.; Kitsuta, K.; Nakashima, S.; Nakanishi, T.; Nakagawa, Y. The lipid fraction of human milk initiates adipocyte differentiation in 3T3-L1 cells. Early. Hum. Dev. 2013, 89, 713–719. [Google Scholar] [CrossRef] [PubMed]
- Santillo, A.; Figliola, L.; Caroprese, M.; Marino, R.; Giardino, I.; Albenzio, M. Effect of lipid fraction of digested milk from different sources in mature 3T3-L1 adipocyte. J. Dairy Res. 2019, 86, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Chomczynski, P.; Sacchi, N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Sztalryd, C.; Brasaemle, D.L. The perilipin family of lipid droplet proteins: Gatekeepers of intracellular lipolysis. B Biophys. Acta Mol. Cell. Biol. Lipids 2017, 1862, 1221–1232. [Google Scholar] [CrossRef]
- Panagos, P.; Vishwanathan, R.; Penfield-Cyr, A.; Matthan, N.; Shivappa, N.; Wirth, M.; Hebert, J.; Sen, S. Breastmilk from obese mothers has pro-inflammatory properties and decreased neuroprotective factors. J. Perinatol. 2016, 36, 284–290. [Google Scholar] [CrossRef]
- Garcia-Ravelo, S.; Díaz-Gómez, N.M.; Martín, M.V.; Dorta-Guerra, R.; Murray, M.; Escuder, D.; Rodriguez, C. Fatty acid composition and eicosanoid levels (LTE4 and PGE2) of human milk from normal weight and overweight mothers. Breastfeed. Med. 2018, 13, 702–710. [Google Scholar] [CrossRef]
- Rudolph, M.C.; Young, B.E.; Lemas, D.J.; Palmer, C.E.; Hernandez, T.L.; Barbour, L.A.; Friedman, J.E.; Krebs, N.F.; MacLean, P.S. Early infant adipose deposition is positively associated with the n-6 to n-3 fatty acid ratio in human milk independent of maternal BMI. Int. J. Obes. 2017, 41, 510–517. [Google Scholar] [CrossRef]
- Saarela, T.; Kokkonen, J.; Koivisto, M. Macronutrient and energy contents of human milk fractions during the first six months of lactation. Acta Paediatr. 2005, 94, 1176–1181. [Google Scholar] [CrossRef]
- Bravi, F.; Wiens, F.; Decarli, A.; Dal Pont, A.; Agostoni, C.; Ferraroni, M. Impact of maternal nutrition on breast-milk composition: A systematic review. Am. J. Clin. Nutr. 2016, 104, 646–662. [Google Scholar] [CrossRef]
- Aumeistere, L.; Ciproviča, I.; Zavadska, D.; Andersons, J.; Volkovs, V.; Ceļmalniece, K. Impact of maternal diet on human milk composition among lactating women in Latvia. Medicina 2019, 55, 173. [Google Scholar] [CrossRef] [PubMed]
- Sprecher, H. Metabolism of highly unsaturated n-3 and n-6 fatty acids. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2000, 1486, 219–231. [Google Scholar] [CrossRef]
- Saito, E.; Okada, T.; Abe, Y.; Odaka, M.; Kuromori, Y.; Iwata, F.; Hara, M.; Mugishima, H.; Kitamura, Y. Abdominal adiposity is associated with fatty acid desaturase activity in boys: Implications for C-reactive protein and insulin resistance. Prostaglandins Leukot Essent. Fat. Acids 2013, 88, 307–311. [Google Scholar] [CrossRef] [PubMed]
- Wolters, M.; Schlenz, H.; Börnhorst, C.; Risé, P.; Galli, C.; Moreno, L.A.; Pala, V.; Siani, A.; Veidebaum, T.; Tornaritis, M. Desaturase activity is associated with weight status and metabolic risk markers in young children. J. Clin. Endocrinol. Metab. 2015, 100, 3760–3769. [Google Scholar] [CrossRef]
- Brenna, J.T.; Varamini, B.; Jensen, R.G.; Diersen-Schade, D.A.; Boettcher, J.A.; Arterburn, L.M. Docosahexaenoic and arachidonic acid concentrations in human breast milk worldwide. Am. J. Clin. Nutr. 2007, 85, 1457–1464. [Google Scholar] [CrossRef]
- Waidyatillake, N.; Dharmage, S.; Allen, K.; Lodge, C.; Simpson, J.; Bowatte, G.; Abramson, M.; Lowe, A. Association of breast milk fatty acids with allergic disease outcomes—A systematic review. Allergy 2018, 73, 295–312. [Google Scholar] [CrossRef]
- Much, D.; Brunner, S.; Vollhardt, C.; Schmid, D.; Sedlmeier, E.-M.; Brüderl, M.; Heimberg, E.; Bartke, N.; Boehm, G.; Bader, B.L. Breast milk fatty acid profile in relation to infant growth and body composition: Results from the INFAT study. Pediatr. Res. 2013, 74, 230–237. [Google Scholar] [CrossRef]
- Nagai, S.; Matsumoto, C.; Shibano, M.; Fujimori, K. Suppression of fatty acid and triglyceride synthesis by the flavonoid orientin through decrease of C/EBPδ expression and inhibition of PI3K/Akt-FOXO1 signaling in adipocytes. Nutrients 2018, 10, 130. [Google Scholar] [CrossRef]
- Hellerstein, M.K. De novo lipogenesis in humans: Metabolic and regulatory aspects. Eur. J. Clin. Nutr. 1999, 53 (Suppl. S1), S53–S65. [Google Scholar] [CrossRef]
- Liu, X.; Strable, M.S.; Ntambi, J.M. Stearoyl CoA desaturase 1: Role in cellular inflammation and stress. Adv. Nutr. 2011, 2, 15–22. [Google Scholar] [CrossRef]
- Popeijus, H.; Saris, W.; Mensink, R. Role of stearoyl-CoA desaturases in obesity and the metabolic syndrome. Int. J. Obes. 2008, 32, 1076–1082. [Google Scholar] [CrossRef] [PubMed]
- ALJohani, A.M.; Syed, D.N.; Ntambi, J.M. Insights into stearoyl-CoA desaturase-1 regulation of systemic metabolism. Trends Endocrinol. Metab. 2017, 28, 831–842. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Michal, J.J.; Tobey, D.J.; Daniels, T.F.; Rule, D.C.; MacNeil, M.D. Significant associations of stearoyl-CoA desaturase (SCD1) gene with fat deposition and composition in skeletal muscle. Int. J. Biol. Sci. 2008, 4, 345. [Google Scholar] [CrossRef] [PubMed]
- Konige, M.; Wang, H.; Sztalryd, C. Role of adipose specific lipid droplet proteins in maintaining whole body energy homeostasis. Biochim. Biophys Acta Mol. Basis Dis. 2014, 1842, 393–401. [Google Scholar] [CrossRef]
- Kim, J.I.; Huh, J.Y.; Sohn, J.H.; Choe, S.S.; Lee, Y.S.; Lim, C.Y.; Jo, A.; Park, S.B.; Han, W.; Kim, J.B. Lipid-overloaded enlarged adipocytes provoke insulin resistance independent of inflammation. Mol. Cell. Biol. 2015, 35, 1686–1699. [Google Scholar] [CrossRef]
- Roberts, R.; Hodson, L.; Dennis, A.; Neville, M.; Humphreys, S.; Harnden, K.; Micklem, K.; Frayn, K. Markers of de novo lipogenesis in adipose tissue: Associations with small adipocytes and insulin sensitivity in humans. Diabetologia 2009, 52, 882–890. [Google Scholar] [CrossRef]
- Koska, J.; Stefan, N.; Dubois, S.; Trinidad, C.; Considine, R.V.; Funahashi, T.; Bunt, J.C.; Ravussin, E.; Permana, P.A. mRNA concentrations of MIF in subcutaneous abdominal adipose cells are associated with adipocyte size and insulin action. Int. J. Obes. 2009, 33, 842–850. [Google Scholar] [CrossRef]
- Gallardo, J.M.; Gómez-López, J.; Medina-Bravo, P.; Juárez-Sánchez, F.; Contreras-Ramos, A.; Galicia-Esquivel, M.; Sánchez-Urbina, R.; Klünder-Klünder, M. Maternal obesity increases oxidative stress in the newborn. Obesity 2015, 23, 1650–1654. [Google Scholar] [CrossRef]
- Negro, S.; Boutsikou, T.; Briana, D.; Tataranno, M.; Longini, M.; Proietti, F.; Bazzini, F.; Dani, C.; Malamitsi-Puchner, A.; Buonocore, G. Maternal obesity and perinatal oxidative stress: The strength of the association. J. Biol. Regul. Homeost. 2017, 31, 221–227. [Google Scholar]
- Okuno, Y.; Fukuhara, A.; Hashimoto, E.; Kobayashi, H.; Kobayashi, S.; Otsuki, M.; Shimomura, I. Oxidative stress inhibits healthy adipose expansion through suppression of SREBF1-mediated lipogenic pathway. Diabetes 2018, 67, 1113–1127. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Maher, F.; Naji, N. Effect of leptin and oxidative stress in the blood of obese individuals. Biochem. Anal. Biochem. 2016, 5, 288. [Google Scholar]
- Berger, S.; Polotsky, V.Y. Leptin and leptin resistance in the pathogenesis of obstructive sleep apnea: A possible link to oxidative stress and cardiovascular complications. Oxid. Med. Cell. Longev. 2018, 2018, 5137947. [Google Scholar] [CrossRef] [PubMed]
- Fortuño, A.; Bidegain, J.; Baltanas, A.; Moreno, M.U.; Montero, L.; Landecho, M.F.; Beloqui, O.; Diez, J.; Zalba, G. Is leptin involved in phagocytic NADPH oxidase overactivity in obesity? Potential clinical implications. J. Hypertens. 2010, 28, 1944–1950. [Google Scholar] [CrossRef] [PubMed]
- Morawietz, H.; Bornstein, S.R. Leptin, endothelin, NADPH oxidase, and heart failure. Hypertension 2006, 47, e20–e21. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Henriksbo, B.D.; Tamrakar, A.K.; Xu, J.; Duggan, B.M.; Cavallari, J.F.; Phulka, J.; Stampfli, M.R.; Ashkar, A.A.; Schertzer, J.D. Statins Promote Interleukin-1β–Dependent Adipocyte Insulin Resistance Through Lower Prenylation, Not Cholesterol. Diabetes 2019, 68, 1441–1448. [Google Scholar] [CrossRef]
- Al-Sulaiti, H.; Dömling, A.S.; Elrayess, M.A. Mediators of Impaired Adipogenesis in Obesity-Associated Insulin Resistance and T2DM. In Adipose Tissue—An Update; IntechOpen: London, UK, 2019. [Google Scholar]
- Sari, A. The Relationship between Leptin and Fatty Acid. J. Mol. Biomark. Diagn. 2013, 4, 2. [Google Scholar] [CrossRef]
- Stern, J.H.; Rutkowski, J.M.; Scherer, P.E. Adiponectin, leptin, and fatty acids in the maintenance of metabolic homeostasis through adipose tissue crosstalk. Cell Metab. 2016, 23, 770–784. [Google Scholar] [CrossRef]
- Bai, Y.; Zhang, S.; Kim, K.-S.; Lee, J.-K.; Kim, K.-H. Obese gene expression alters the ability of 30A5 preadipocytes to respond to lipogenic hormones. J. Biol. Chem. 1996, 271, 13939–13942. [Google Scholar] [CrossRef]
- Soukas, A.; Cohen, P.; Socci, N.D.; Friedman, J.M. Leptin-specific patterns of gene expression in white adipose tissue. Genes Dev. 2000, 14, 963–980. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Non-Obese | Obese | |
|---|---|---|
| N | 20 | 20 |
| Age (years), mean (SD) | 31.5 (6.2) | 29 (7) |
| BMI (kg/m2), mean (SD) | 25 (2.5) | 33 (3.7) |
| Race/Ethnicity, n (%) | ||
| Caucasian, non-Hispanic | 15 (75%) | 9 (45%) |
| Hispanic | 3 (15%) | 7 (35%) |
| African American, non-Hispanic | 0 | 4 (20%) |
| Asian or Pacific Islander | 2 (10%) | 0 |
| Education Level n (%) | ||
| High school | ||
| Some college | 3 (15%) | 3 (15%) |
| Associate degree (2 years college) | 1 (5%) | 0 |
| Bachelor’s degree | 3 (15%) | 5 (25%) |
| Master’s degree | 7 (35%) | 3 (15%) |
| Ph.D./Sc.D./M.D. | 1 (5%) | 1 (5%) |
| Fatty Acid Name | FA (nmol%) | Non-Obese | Obese | p-Value |
|---|---|---|---|---|
| Palmitic acid | C16:0 | 22.57 ± 2.45 | 21.41 ± 2.96 | p = 0.104 |
| Stearic acid | C18:0 | 6.77 ± 1.08 a | 5.62 ±0.99 b | p < 0.0001 |
| Palmitoleic acid | C16:1 | 2.55 ± 0.74 | 2.49 ± 0.76 | p = 0.669 |
| Oleic acid | C18:1 | 33.02 ± 3.19 a | 30.46 ± 3.96 b | p < 0.001 |
| Eicosenoic acid | C20:1n9 | 0.35 ± 0.06 | 0.33 ± 0.06 | p = 0.141 |
| Linoleic acid (LA) | C18:2n6 | 22.98 ± 4.55 a | 27.10 ± 4.23 b | p < 0.0001 |
| γ-Linolenic acid (GLA) | C18:3n6 | 0.26 ± 0.04 | 0.28 ± 0.06 | p = 0.462 |
| Eicosatrienoic acid | C20:3n6 | 0.72 ± 0.09 | 0.83 ± 0.19 | p = 0.023 |
| Arachidonic acid (AA) | C20:4n6 | 0.75 ± 0.11 | 0.79 ± 0.12 | p = 0.573 |
| α-Linolenic acid (ALA) | C18:3n3 | 1.71 ± 0.53 | 1.81 ± 0.46 | p = 0.777 |
| Eicosatrienoic acid | C20:3n3 | 0.19 ± 0.04 | 0.20 ± 0.04 | p = 0.886 |
| Eicosatetraenoic acid | C20:4n3 | 0.16 ± 0.02 | 0.16 ± 0.02 | p = 0.784 |
| Eicosapentaenoic acid (EPA) | C20:5n3 | 0.23 ± 0.06 | 0.22 ± 0.05 | p = 0.420 |
| Docosapentaenoic acid | C22:5n3 | 0.34 ± 0.06 | 0.36 ± 0.08 | p = 0.709 |
| Docosahexaenoic acid (DHA) | C22:6n3 | 0.46 ± 0.11 | 0.41 ± 0.12 | p = 0.161 |
| ΣSFA | 36.27 ± 3.77 | 34.56 ± 4.66 | p = 0.072 | |
| ΣMUFA | 35.92 ± 3.12 | 33.28 ± 6.68 | p = 0.510 | |
| ΣPUFA | 27.81 ± 5.04 b | 32.16 ± 4.52 a | p < 0.0001 | |
| Σn6-PUFA | 24.70 ± 4.58 b | 29.00 ± 4.24 a | p = 0.037 | |
| Σn3-PUFA | 3.11 ± 0.59 | 3.17 ± 0.49 | p = 0.937 | |
| n-6:n-3 PUFA | 7.95 ± 1.38 a | 9.15 ± 1.56 b | p < 0.0001 | |
| D6D | 0.01 ± 0.01 b | 0.13 ± 0.01 a | p < 0.001 | |
| D5D | 1.04 ± 0.01 | 0.05 ± 0.63 | p = 0.159 |
| Non-Obese | Obese | |||||
| Fatty Acid Name | FA (nmol%) | Q1 | Q2 | Q3 | Q4 | p-Value |
| Myristic acid | C14:0 | 8.00 ± 1.74 | 5.96 ± 1.03 | 7.56 ± 0.96 | 7.50 ± 3.20 | p = 0.220 |
| Palmitic acid | C16:0 | 23.90 ± 2.44 | 20.78 ± 1.12 | 22.51 ± 1.08 | 20.49 ± 1.49 | p = 0.065 |
| Stearic acid | C18:0 | 7.80 ± 1.06 | 6.17 ± 0.98 | 6.03 ± 0.83 | 5.99 ± 1.25 | p = 0.086 |
| Palmitoleic acid | C16:1 | 2.45 ± 0.16 | 2.56 ± 0.64 | 2.63 ± 0.41 | 2.00 ± 0.79 | p = 0.549 |
| Oleic acid | C18:1 | 33.35 ± 2.71 | 33.65 ± 4.97 | 30.29 ± 2.53 | 32.05 ± 3.76 | p = 0.405 |
| Eicosenoic acid | C20:1n9 | 0.36 ± 0.06 | 0.37 ± 0.09 | 0.30 ± 0.02 | 0.36 ± 0.07 | p = 0.436 |
| Linoleic acid (LA) | C18:2n6 | 19.35 ± 2.88 a | 25.91 ±4.41 b | 25.26 ±5.03 b | 27.21 ± 5.02 b | p = 0.041 |
| γ-Linolenic acid (GLA) | C18:3n6 | 0.26 ± 0.06 | 0.25 ± 0.02 | 0.29 ± 0.06 | 0.29 ± 0.06 | p = 0.700 |
| Eicosatrienoic acid | C20:3n6 | 0.68 ± 0.13 | 0.78 ± 0.04 | 0.74 ± 0.07 | 0.78 ± 0.05 | p = 0.226 |
| Arachidonic acid (AA) | C20:4n6 | 0.74 ± 0.08 | 0.74 ± 0.13 | 0.84 ± 0.12 | 0.77 ± 0.09 | p = 0.554 |
| α-Linolenic acid (ALA) | C18:3n3 | 1.56 ± 0.21 | 1.62 ± 0.64 | 2.01 ± 0.51 | 1.30 ± 0.27 | p = 0.257 |
| Eicosatrienoic acid | C20:3n3 | 0.20 ± 0.04 | 0.17 ± 0.03 | 0.20 ± 0.03 | 0.20 ± 0.02 | p = 0.401 |
| Eicosatetraenoic acid | C20:4n3 | 0.18 ± 0.02 | 0.14 ± 0.02 | 0.16 ± 0.02 | 0.15 ± 0.01 | p = 0.303 |
| Eicosapentaenoic acid (EPA) | C20:5n3 | 0.27 ± 0.08 | 0.19 ± 0.03 | 0.25 ± 0.05 | 0.21 ± 0.03 | p = 0.211 |
| Docosapentaenoic acid | C22:5n3 | 0.37 ± 0.07 | 0.31 ± 0.05 | 0.42 ± 0.07 | 0.32 ± 0.04 | p = 0.127 |
| Docosahexaenoic acid (DHA) | C22:6n3 | 0.52 ± 0.09 | 0.42 ± 0.12 | 0.51 ± 0.16 | 0.38 ± 0.03 | p = 0.386 |
| ΣSFA | 39.71 ± 3.26 a | 32.91 ± 2.59 b | 36.09 ± 2.42 ab | 33.98 ± 5.23 ab | p = 0.045 | |
| ΣMUFA | 36.16 ± 2.76 | 36.58 ± 4.65 | 33.23 ± 2.28 | 34.41 ± 4.57 | p = 0.517 | |
| ΣPUFA | 24.13 ± 3.33 b | 30.51 ± 5.16 a | 30.68 ± 5.17 a | 31.61 ± 5.41 a | p = 0.041 | |
| Σn6-PUFA | 21.03 ± 2.88 b | 27.68 ± 4.50 a | 27.13 ± 4.90 a | 29.05 ± 5.02 a | p = 0.031 | |
| Σn3-PUFA | 3.10 ± 0.44 | 2.84 ± 0.69 | 3.55 ± 0.34 | 2.56 ± 0.39 | p = 0.098 | |
| n-6:n-3 PUFA | 6.77 ± 0.07 c | 9.76 ± 1.31 b | 7.63 ± 1.06 c | 11.35 ± 0.50 a | p < 0.001 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Isesele, P.; Enstad, S.; Huong, P.; Thomas, R.; Wagner, C.L.; Sen, S.; Cheema, S.K. Breast Milk from Non-Obese Women with a High Omega-6 to Omega-3 Fatty Acid Ratio, but Not from Women with Obesity, Increases Lipogenic Gene Expression in 3T3-L1 Preadipocytes, Suggesting Adipocyte Dysfunction. Biomedicines 2022, 10, 1129. https://doi.org/10.3390/biomedicines10051129
Isesele P, Enstad S, Huong P, Thomas R, Wagner CL, Sen S, Cheema SK. Breast Milk from Non-Obese Women with a High Omega-6 to Omega-3 Fatty Acid Ratio, but Not from Women with Obesity, Increases Lipogenic Gene Expression in 3T3-L1 Preadipocytes, Suggesting Adipocyte Dysfunction. Biomedicines. 2022; 10(5):1129. https://doi.org/10.3390/biomedicines10051129
Chicago/Turabian StyleIsesele, Peter, Samantha Enstad, Pham Huong, Raymond Thomas, Carol L. Wagner, Sarbattama Sen, and Sukhinder K. Cheema. 2022. "Breast Milk from Non-Obese Women with a High Omega-6 to Omega-3 Fatty Acid Ratio, but Not from Women with Obesity, Increases Lipogenic Gene Expression in 3T3-L1 Preadipocytes, Suggesting Adipocyte Dysfunction" Biomedicines 10, no. 5: 1129. https://doi.org/10.3390/biomedicines10051129
APA StyleIsesele, P., Enstad, S., Huong, P., Thomas, R., Wagner, C. L., Sen, S., & Cheema, S. K. (2022). Breast Milk from Non-Obese Women with a High Omega-6 to Omega-3 Fatty Acid Ratio, but Not from Women with Obesity, Increases Lipogenic Gene Expression in 3T3-L1 Preadipocytes, Suggesting Adipocyte Dysfunction. Biomedicines, 10(5), 1129. https://doi.org/10.3390/biomedicines10051129