
The Corpus Adiposum Infrapatellare (Hoffa’s Fat Pad)—The Role of the Infrapatellar Fat Pad in Osteoarthritis Pathogenesis

 , , and
, , and
Abstract
:1. Introduction
2. Methodology
3. Anatomy and Function of IFP
3.1. Anatomy of the IFP
3.2. Mechanical Aspects
3.3. Contribution to the Pathogenesis of Knee OA
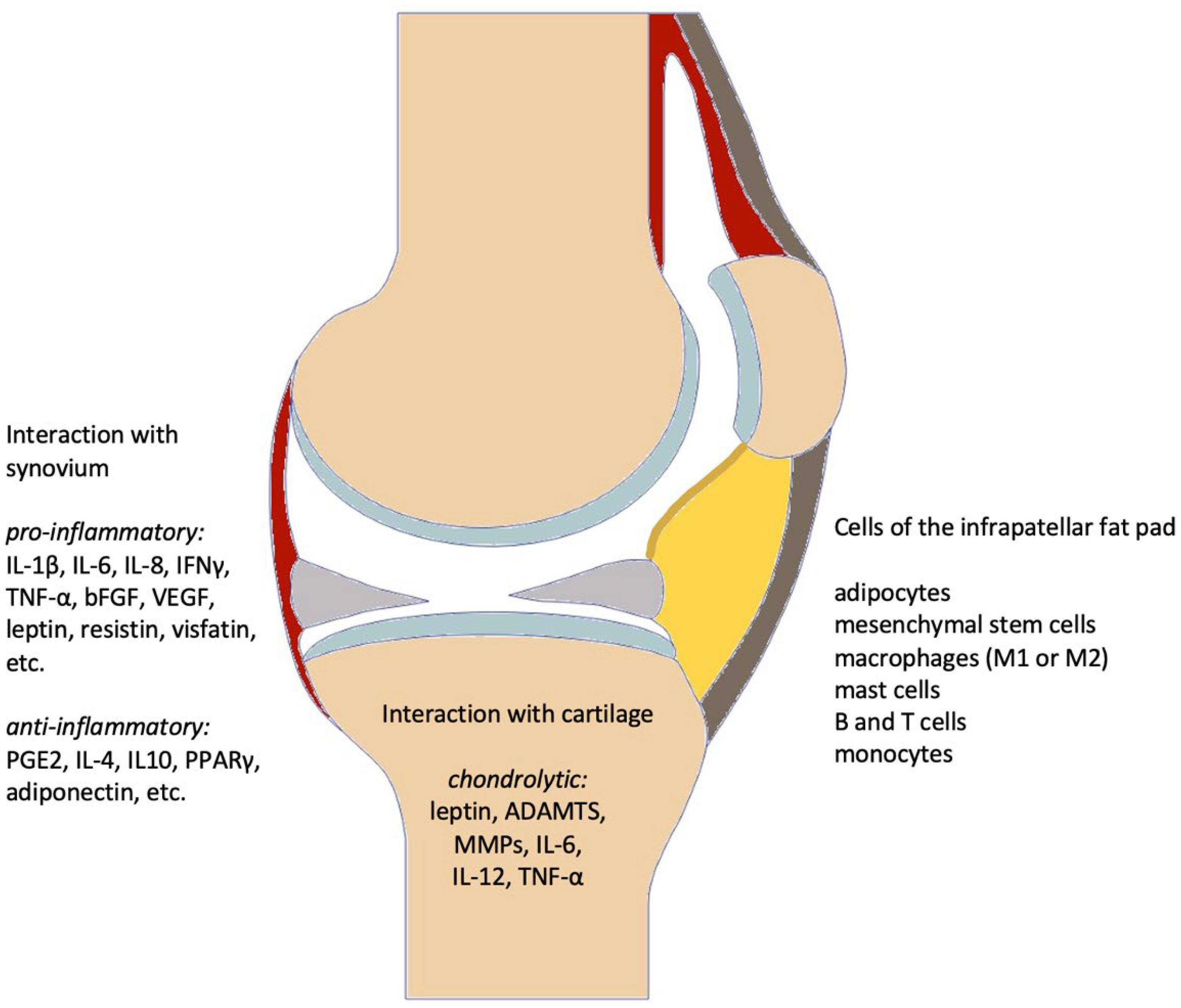
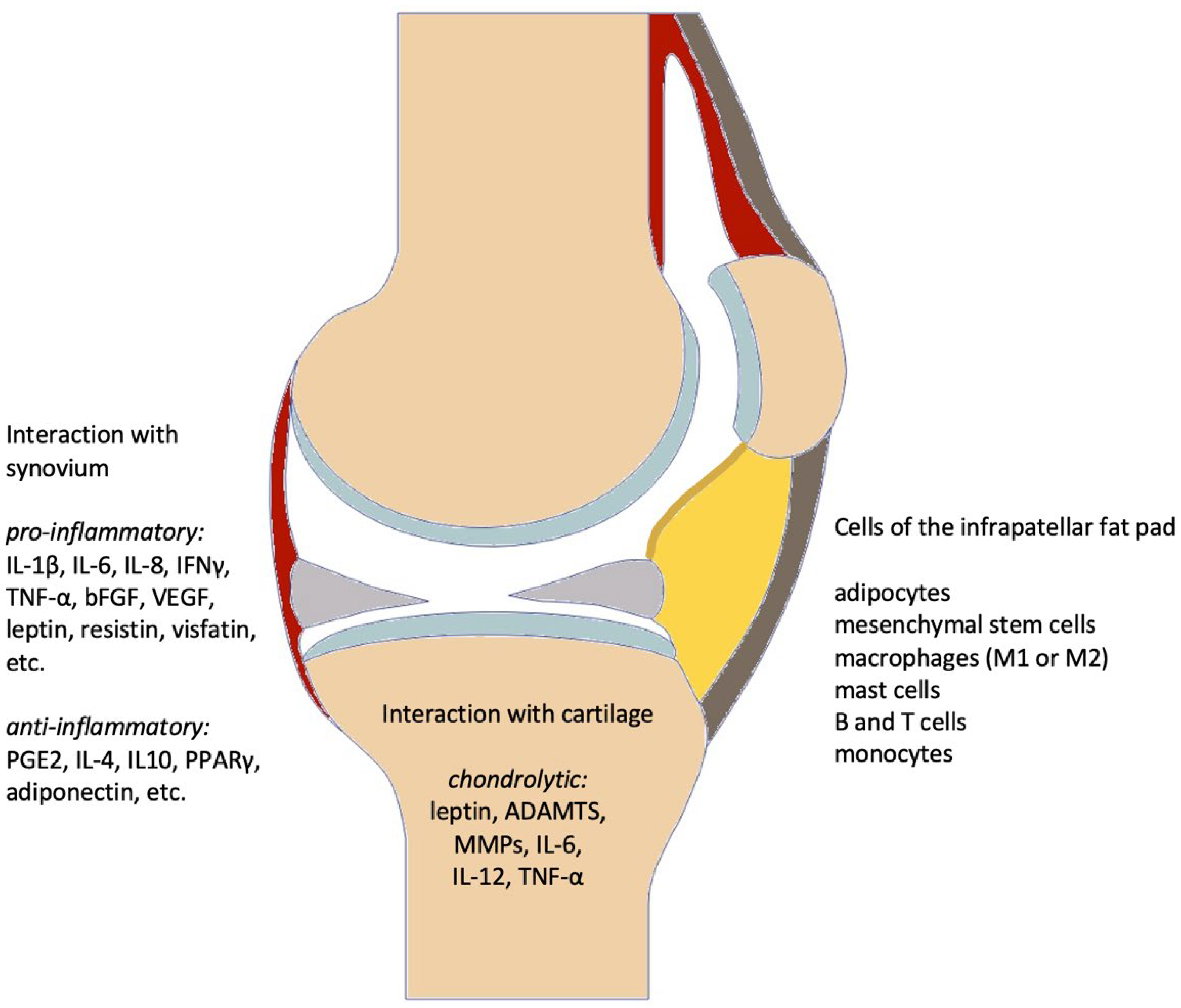
3.3.1. Secretory Profile
3.3.2. Immunological Role
3.3.3. IFP-Derived MSCs
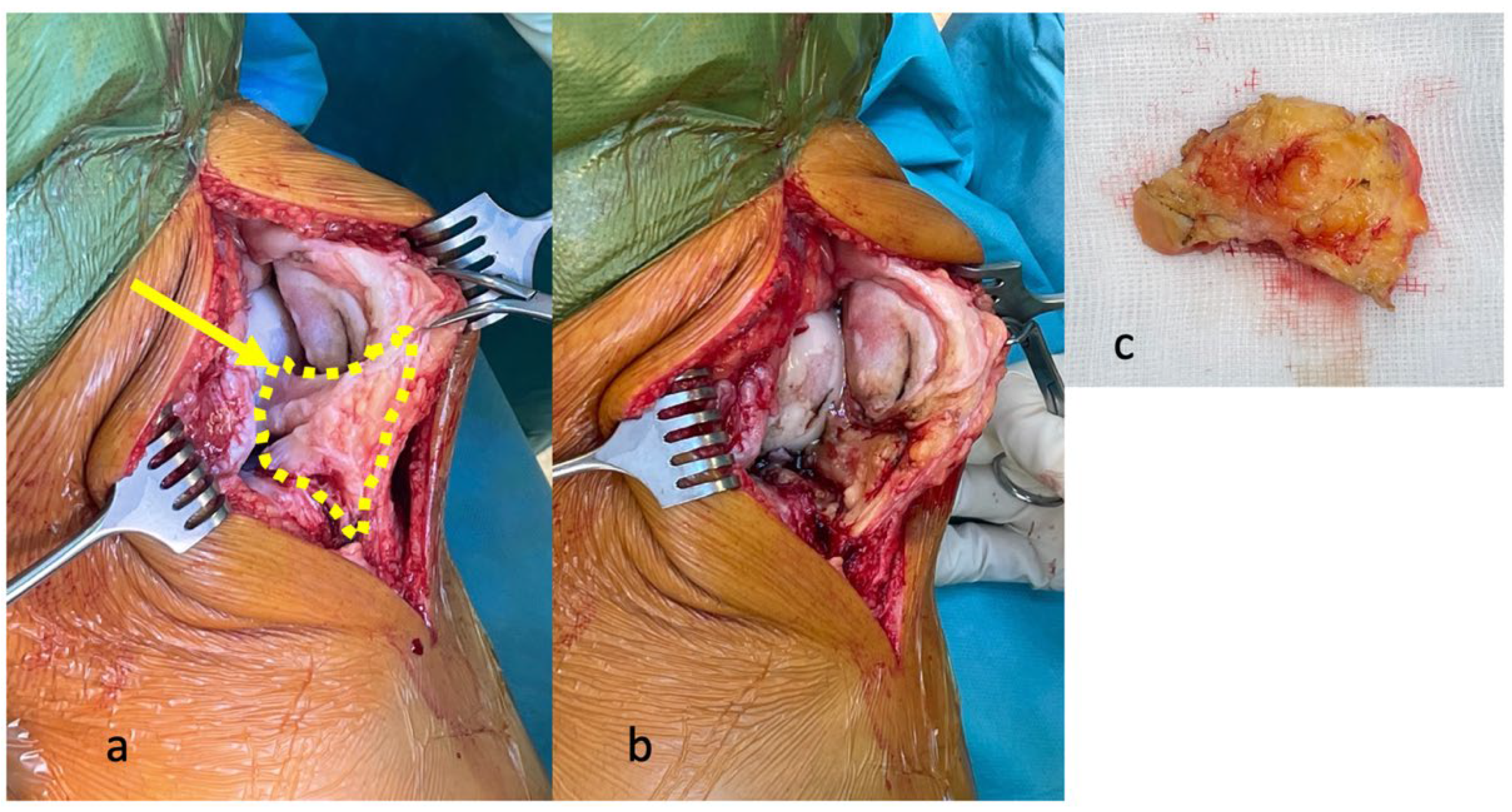
4. Effects of Surgical Resection of the IFP in TKA
4.1. Anterior Knee Pain
4.2. Vascularization of the Patella
4.3. Range of Motion and Functional Outcome
4.4. Length of the Patellar Tendon
4.5. Complications
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hoffa, A. Influence of adipose tissue with regard to the pathology of the knee joint. J. Am. Med. Assoc. 1904, 43, 795–796. [Google Scholar] [CrossRef] [Green Version]
- Van Duren, B.H.; Lamb, J.N.; Nisar, S.; Ashraf, Y.; Somashekar, N.; Pandit, H. Preservation vs. resection of the infrapatellar fat pad during total knee arthroplasty Part I: A survey of current practice in the UK. Knee 2019, 26, 416–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bastiaansen-Jenniskens, Y.M.; Clockaerts, S.; Feijt, C.; Zuurmond, A.M.; Stojanovic-Susulic, V.; Bridts, C.; de Clerck, L.; DeGroot, J.; Verhaar, J.A.; Kloppenburg, M.; et al. Infrapatellar fat pad of patients with end-stage osteoarthritis inhibits catabolic mediators in cartilage. Ann. Rheum. Dis. 2012, 71, 288–294. [Google Scholar] [CrossRef] [PubMed]
- Macchi, V.; Stocco, E.; Stecco, C.; Belluzzi, E.; Favero, M.; Porzionato, A.; De Caro, R. The infrapatellar fat pad and the synovial membrane: An anatomo-functional unit. J. Anat. 2018, 233, 146–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dragoo, J.L.; Johnson, C.; McConnell, J. Evaluation and treatment of disorders of the infrapatellar fat pad. Sports Med. 2012, 42, 51–67. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, J.; Tierney, P.; Murray, P.; O’Brien, M. The infrapatellar fat pad: Anatomy and clinical correlations. Knee Surg. Sports Traumatol. Arthrosc. 2005, 13, 268–272. [Google Scholar] [CrossRef] [PubMed]
- Diepold, J.; Ruhdorfer, A.; Dannhauer, T.; Wirth, W.; Steidle, E.; Eckstein, F. Sex-differences of the healthy infra-patellar (Hoffa) fat pad in relation to intermuscular and subcutaneous fat content-data from the Osteoarthritis Initiative. Ann. Anat. 2015, 200, 30–36. [Google Scholar] [CrossRef] [Green Version]
- Burda, B.; Steidle-Kloc, E.; Dannhauer, T.; Wirth, W.; Ruhdorfer, A.; Eckstein, F. Variance in infra-patellar fat pad volume: Does the body mass index matter?-Data from osteoarthritis initiative participants without symptoms or signs of knee disease. Ann. Anat. 2017, 213, 19–24. [Google Scholar] [CrossRef]
- Ioan-Facsinay, A.; Kloppenburg, M. An emerging player in knee osteoarthritis: The infrapatellar fat pad. Arthritis Res. Ther. 2013, 15, 225. [Google Scholar] [CrossRef] [Green Version]
- Eymard, F.; Pigenet, A.; Citadelle, D.; Tordjman, J.; Foucher, L.; Rose, C.; Flouzat Lachaniette, C.H.; Rouault, C.; Clément, K.; Berenbaum, F.; et al. Knee and hip intra-articular adipose tissues (IAATs) compared with autologous subcutaneous adipose tissue: A specific phenotype for a central player in osteoarthritis. Ann. Rheum. Dis. 2017, 76, 1142–1148. [Google Scholar] [CrossRef] [Green Version]
- Favero, M.; El-Hadi, H.; Belluzzi, E.; Granzotto, M.; Porzionato, A.; Sarasin, G.; Rambaldo, A.; Iacobellis, C.; Cigolotti, A.; Fontanella, C.G.; et al. Infrapatellar fat pad features in osteoarthritis: A histopathological and molecular study. Rheumatology 2017, 56, 1784–1793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bohnsack, M.; Meier, F.; Walter, G.F.; Hurschler, C.; Schmolke, S.; Wirth, C.J.; Rühmann, O. Distribution of substance-P nerves inside the infrapatellar fat pad and the adjacent synovial tissue: A neurohistological approach to anterior knee pain syndrome. Arch. Orthop. Trauma Surg. 2005, 125, 592–597. [Google Scholar] [CrossRef] [PubMed]
- Eymard, F.; Chevalier, X. Inflammation of the infrapatellar fat pad. Jt. Bone Spine 2016, 83, 389–393. [Google Scholar] [CrossRef] [PubMed]
- Fontanella, C.G.; Belluzzi, E.; Rossato, M.; Olivotto, E.; Trisolino, G.; Ruggieri, P.; Rubini, A.; Porzionato, A.; Natali, A.; De Caro, R.; et al. Quantitative MRI analysis of infrapatellar and suprapatellar fat pads in normal controls, moderate and end-stage osteoarthritis. Ann. Anat. 2019, 221, 108–114. [Google Scholar] [CrossRef]
- Walsh, D.A.; Mapp, P.I.; Kelly, S. Calcitonin gene-related peptide in the joint: Contributions to pain and inflammation. Br. J. Clin. Pharmacol. 2015, 80, 965–978. [Google Scholar] [CrossRef] [Green Version]
- Lehner, B.; Koeck, F.X.; Capellino, S.; Schubert, T.E.; Hofbauer, R.; Straub, R.H. Preponderance of sensory versus sympathetic nerve fibers and increased cellularity in the infrapatellar fat pad in anterior knee pain patients after primary arthroplasty. J. Orthop. Res. 2008, 26, 342–350. [Google Scholar] [CrossRef]
- Koeck, F.X.; Schmitt, M.; Baier, C.; Stangl, H.; Beckmann, J.; Grifka, J.; Straub, R.H. Predominance of synovial sensory nerve fibers in arthrofibrosis following total knee arthroplasty compared to osteoarthritis of the knee. J. Orthop. Surg. Res. 2016, 11, 25. [Google Scholar] [CrossRef] [Green Version]
- Belluzzi, E.; El Hadi, H.; Granzotto, M.; Rossato, M.; Ramonda, R.; Macchi, V.; De Caro, R.; Vettor, R.; Favero, M. Systemic and Local Adipose Tissue in Knee Osteoarthritis. J. Cell. Physiol. 2017, 232, 1971–1978. [Google Scholar] [CrossRef]
- Zeng, N.; Yan, Z.P.; Chen, X.Y.; Ni, G.X. Infrapatellar Fat Pad and Knee Osteoarthritis. Aging Dis. 2020, 11, 1317–1328. [Google Scholar] [CrossRef]
- Belluzzi, E.; Stocco, E.; Pozzuoli, A.; Granzotto, M.; Porzionato, A.; Vettor, R.; De Caro, R.; Ruggieri, P.; Ramonda, R.; Rossato, M.; et al. Contribution of Infrapatellar Fat Pad and Synovial Membrane to Knee Osteoarthritis Pain. Biomed. Res. Int. 2019, 2019, 6390182. [Google Scholar] [CrossRef]
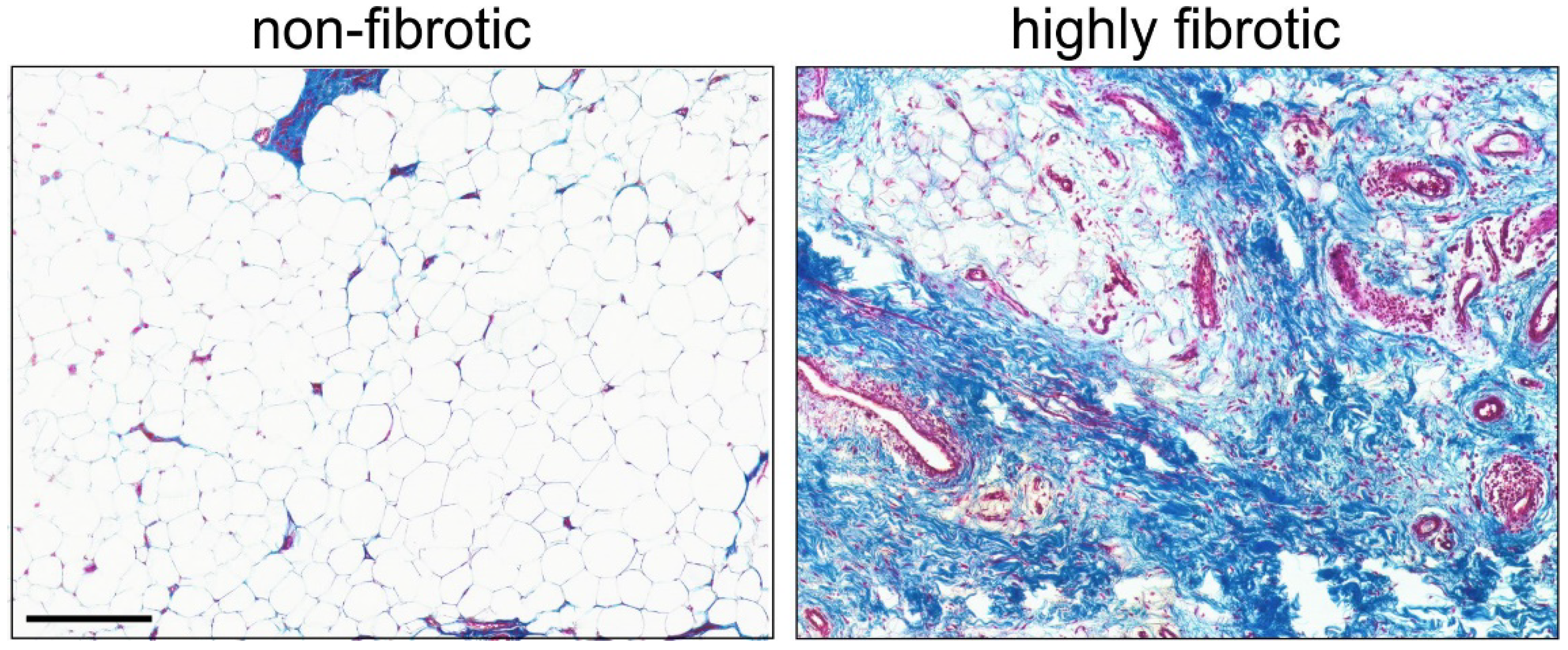
- Stocco, E.; Belluzzi, E.; Contran, M.; Boscolo-Berto, R.; Picardi, E.; Guidolin, D.; Fontanella, C.G.; Olivotto, E.; Filardo, G.; Borile, G.; et al. Age-Dependent Remodeling in Infrapatellar Fat Pad Adipocytes and Extracellular Matrix: A Comparative Study. Front. Med. 2021, 8, 661403. [Google Scholar] [CrossRef] [PubMed]
- Distel, E.; Cadoudal, T.; Durant, S.; Poignard, A.; Chevalier, X.; Benelli, C. The infrapatellar fat pad in knee osteoarthritis: An important source of interleukin-6 and its soluble receptor. Arthritis Rheum. 2009, 60, 3374–3377. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Maleitzke, T.; Geissler, S.; Hildebrandt, A.; Fleckenstein, F.; Niemann, M.; Fischer, H.; Perka, C.; Duda, G.N.; Winkler, T. Source and Hub of Inflammation—The Infrapatellar Fat Pad and its Interactions with Articular Tissues during Knee Osteoarthritis. J. Orthop. Res. 2022. [Google Scholar] [CrossRef] [PubMed]
- Clockaerts, S.; Bastiaansen-Jenniskens, Y.M.; Runhaar, J.; Van Osch, G.J.; Van Offel, J.F.; Verhaar, J.A.; De Clerck, L.S.; Somville, J. The infrapatellar fat pad should be considered as an active osteoarthritic joint tissue: A narrative review. Osteoarthr. Cartil. 2010, 18, 876–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greif, D.N.; Kouroupis, D.; Murdock, C.J.; Griswold, A.J.; Kaplan, L.D.; Best, T.M.; Correa, D. Infrapatellar Fat Pad/Synovium Complex in Early-Stage Knee Osteoarthritis: Potential New Target and Source of Therapeutic Mesenchymal Stem/Stromal Cells. Front. Bioeng. Biotechnol. 2020, 8, 860. [Google Scholar] [CrossRef] [PubMed]
- Fontanella, C.G.; Macchi, V.; Carniel, E.L.; Frigo, A.; Porzionato, A.; Picardi, E.E.E.; Favero, M.; Ruggieri, P.; de Caro, R.; Natali, A.N. Biomechanical behavior of Hoffa’s fat pad in healthy and osteoarthritic conditions: Histological and mechanical investigations. Australas Phys. Eng. Sci. Med. 2018, 41, 657–667. [Google Scholar] [CrossRef]
- Fontanella, C.G.; Belluzzi, E.; Pozzuoli, A.; Favero, M.; Ruggieri, P.; Macchi, V.; Carniel, E.L. Mechanical behavior of infrapatellar fat pad of patients affected by osteoarthritis. J. Biomech. 2022, 131, 110931. [Google Scholar] [CrossRef]
- Klein-Wieringa, I.R.; de Lange-Brokaar, B.J.; Yusuf, E.; Andersen, S.N.; Kwekkeboom, J.C.; Kroon, H.M.; van Osch, G.J.; Zuurmond, A.M.; Stojanovic-Susulic, V.; Nelissen, R.G.; et al. Inflammatory Cells in Patients with Endstage Knee Osteoarthritis: A Comparison between the Synovium and the Infrapatellar Fat Pad. J. Rheumatol. 2016, 43, 771–778. [Google Scholar] [CrossRef]
- Jiang, L.F.; Fang, J.H.; Wu, L.D. Role of infrapatellar fat pad in pathological process of knee osteoarthritis: Future applications in treatment. World J. Clin. Cases 2019, 7, 2134–2142. [Google Scholar] [CrossRef]
- Wiegertjes, R.; van de Loo, F.A.J.; Blaney Davidson, E.N. A roadmap to target interleukin-6 in osteoarthritis. Rheumatology 2020, 59, 2681–2694. [Google Scholar] [CrossRef]
- Jenei-Lanzl, Z.; Meurer, A.; Zaucke, F. Interleukin-1β signaling in osteoarthritis—Chondrocytes in focus. Cell. Signal. 2019, 53, 212–223. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, T.; Iwabu, M.; Okada-Iwabu, M.; Kadowaki, T. Adiponectin receptors: A review of their structure, function and how they work. Best Pract. Res. Clin. Endocrinol. Metab. 2014, 28, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, K.; Parker, J.L.; Ouchi, N.; Higuchi, A.; Vita, J.A.; Gokce, N.; Pedersen, A.A.; Kalthoff, C.; Tullin, S.; Sams, A.; et al. Adiponectin promotes macrophage polarization toward an anti-inflammatory phenotype. J. Biol. Chem. 2010, 285, 6153–6160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vuolteenaho, K.; Koskinen, A.; Kukkonen, M.; Nieminen, R.; Päivärinta, U.; Moilanen, T.; Moilanen, E. Leptin enhances synthesis of proinflammatory mediators in human osteoarthritic cartilage-mediator role of NO in leptin-induced PGE2, IL-6, and IL-8 production. Mediat. Inflamm. 2009, 2009, 345838. [Google Scholar] [CrossRef] [Green Version]
- Hui, W.; Litherland, G.J.; Elias, M.S.; Kitson, G.I.; Cawston, T.E.; Rowan, A.D.; Young, D.A. Leptin produced by joint white adipose tissue induces cartilage degradation via upregulation and activation of matrix metalloproteinases. Ann. Rheum. Dis. 2012, 71, 455–462. [Google Scholar] [CrossRef]
- Conde, J.; Scotece, M.; Abella, V.; Gómez, R.; López, V.; Villar, R.; Hermida, M.; Pino, J.; Gómez-Reino, J.J.; Gualillo, O. Identification of novel adipokines in the joint. Differential expression in healthy and osteoarthritis tissues. PLoS ONE 2015, 10, e0123601. [Google Scholar] [CrossRef] [Green Version]
- Furuhashi, M.; Saitoh, S.; Shimamoto, K.; Miura, T. Fatty Acid-Binding Protein 4 (FABP4): Pathophysiological Insights and Potent Clinical Biomarker of Metabolic and Cardiovascular Diseases. Clin. Med. Insights Cardiol. 2014, 8, 23–33. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Li, T.; Chiu, K.Y.; Wen, C.; Xu, A.; Yan, C.H. FABP4 as a biomarker for knee osteoarthritis. Biomark. Med. 2018, 12, 107–118. [Google Scholar] [CrossRef]
- Ji, J.; Jia, S.; Ji, K.; Jiang, W.G. Wnt1 inducible signalling pathway protein-2 (WISP-2/CCN5): Roles and regulation in human cancers (review). Oncol. Rep. 2014, 31, 533–539. [Google Scholar] [CrossRef] [Green Version]
- Maiese, K. Picking a bone with WISP1 (CCN4): New strategies against degenerative joint disease. J. Transl. Sci. 2016, 1, 83–85. [Google Scholar] [CrossRef] [Green Version]
- Xie, C.; Chen, Q. Adipokines: New Therapeutic Target for Osteoarthritis? Curr. Rheumatol. Rep. 2019, 21, 71. [Google Scholar] [CrossRef] [PubMed]
- Takano, S.; Uchida, K.; Inoue, G.; Matsumoto, T.; Aikawa, J.; Iwase, D.; Mukai, M.; Miyagi, M.; Takaso, M. Vascular endothelial growth factor expression and their action in the synovial membranes of patients with painful knee osteoarthritis. BMC Musculoskelet. Disord. 2018, 19, 204. [Google Scholar] [CrossRef] [PubMed]
- Ellman, M.B.; Yan, D.; Ahmadinia, K.; Chen, D.; An, H.S.; Im, H.J. Fibroblast growth factor control of cartilage homeostasis. J. Cell. Biochem. 2013, 114, 735–742. [Google Scholar] [CrossRef] [Green Version]
- Gavrilovic, J. Fibroblast growth factor 2: A new key player in osteoarthritis. Arthritis Rheum. 2009, 60, 1869–1872. [Google Scholar] [CrossRef] [PubMed]
- Klein-Wieringa, I.R.; Kloppenburg, M.; Bastiaansen-Jenniskens, Y.M.; Yusuf, E.; Kwekkeboom, J.C.; El-Bannoudi, H.; Nelissen, R.G.; Zuurmond, A.; Stojanovic-Susulic, V.; Van Osch, G.J.; et al. The infrapatellar fat pad of patients with osteoarthritis has an inflammatory phenotype. Ann. Rheum. Dis. 2011, 70, 851–857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belluzzi, E.; Macchi, V.; Fontanella, C.G.; Carniel, E.L.; Olivotto, E.; Filardo, G.; Sarasin, G.; Porzionato, A.; Granzotto, M.; Pozzuoli, A.; et al. Infrapatellar Fat Pad Gene Expression and Protein Production in Patients with and without Osteoarthritis. Int. J. Mol. Sci. 2020, 21, 6016. [Google Scholar] [CrossRef]
- Apinun, J.; Sengprasert, P.; Yuktanandana, P.; Ngarmukos, S.; Tanavalee, A.; Reantragoon, R. Immune Mediators in Osteoarthritis: Infrapatellar Fat Pad-Infiltrating CD8+ T Cells Are Increased in Osteoarthritic Patients with Higher Clinical Radiographic Grading. Int. J. Rheumatol. 2016, 2016, 9525724. [Google Scholar] [CrossRef]
- Orlowsky, E.W.; Kraus, V.B. The role of innate immunity in osteoarthritis: When our first line of defense goes on the offensive. J. Rheumatol. 2015, 42, 363–371. [Google Scholar] [CrossRef] [Green Version]
- Liu-Bryan, R. Synovium and the innate inflammatory network in osteoarthritis progression. Curr. Rheumatol. Rep. 2013, 15, 323. [Google Scholar] [CrossRef] [Green Version]
- Woodell-May, J.E.; Sommerfeld, S.D. Role of Inflammation and the Immune System in the Progression of Osteoarthritis. J. Orthop. Res. 2020, 38, 253–257. [Google Scholar] [CrossRef]
- Gomez, R.; Lago, F.; Gomez-Reino, J.; Dieguez, C.; Gualillo, O. Adipokines in the skeleton: Influence on cartilage function and joint degenerative diseases. J. Mol. Endocrinol. 2009, 43, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Ouchi, N.; Parker, J.L.; Lugus, J.J.; Walsh, K. Adipokines in inflammation and metabolic disease. Nat. Rev. Immunol. 2011, 11, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, R.; Takahashi, M.; Virtanen, C.; Syed, K.; Davey, J.R.; Mahomed, N.N. Microarray analysis of the infrapatellar fat pad in knee osteoarthritis: Relationship with joint inflammation. J. Rheumatol. 2011, 38, 1966–1972. [Google Scholar] [CrossRef] [PubMed]
- Buckley, M.G.; Gallagher, P.J.; Walls, A.F. Mast cell subpopulations in the synovial tissue of patients with osteoarthritis: Selective increase in numbers of tryptase-positive, chymase-negative mast cells. J. Pathol. 1998, 186, 67–74. [Google Scholar] [CrossRef]
- Gotis-Graham, I.; McNeil, H.P. Mast cell responses in rheumatoid synovium. Association of the MCTC subset with matrix turnover and clinical progression. Arthritis Rheum. 1997, 40, 479–489. [Google Scholar] [CrossRef]
- De Lange-Brokaar, B.J.; Kloppenburg, M.; Andersen, S.N.; Dorjée, A.L.; Yusuf, E.; Herb-van Toorn, L.; Kroon, H.M.; Zuurmond, A.M.; Stojanovic-Susulic, V.; Bloem, J.L.; et al. Characterization of synovial mast cells in knee osteoarthritis: Association with clinical parameters. Osteoarthr. Cartil. 2016, 24, 664–671. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Lepus, C.M.; Raghu, H.; Reber, L.L.; Tsai, M.M.; Wong, H.H.; von Kaeppler, E.; Lingampalli, N.; Bloom, M.S.; Hu, N.; et al. IgE-mediated mast cell activation promotes inflammation and cartilage destruction in osteoarthritis. Elife 2019, 8, e39905. [Google Scholar] [CrossRef]
- Magarinos, N.J.; Bryant, K.J.; Fosang, A.J.; Adachi, R.; Stevens, R.L.; McNeil, H.P. Mast cell-restricted, tetramer-forming tryptases induce aggrecanolysis in articular cartilage by activating matrix metalloproteinase-3 and -13 zymogens. J. Immunol. 2013, 191, 1404–1412. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Chen, S.; Pei, M. Comparative advantages of infrapatellar fat pad: An emerging stem cell source for regenerative medicine. Rheumatology 2018, 57, 2072–2086. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Buckley, C.T.; Almeida, H.V.; Mulhall, K.J.; Kelly, D.J. Infrapatellar fat pad-derived stem cells maintain their chondrogenic capacity in disease and can be used to engineer cartilaginous grafts of clinically relevant dimensions. Tissue Eng. Part A 2014, 20, 3050–3062. [Google Scholar] [CrossRef] [Green Version]
- Stocco, E.; Barbon, S.; Piccione, M.; Belluzzi, E.; Petrelli, L.; Pozzuoli, A.; Ramonda, R.; Rossato, M.; Favero, M.; Ruggieri, P.; et al. Infrapatellar Fat Pad Stem Cells Responsiveness to Microenvironment in Osteoarthritis: From Morphology to Function. Front. Cell Dev. Biol. 2019, 7, 323. [Google Scholar] [CrossRef] [PubMed]
- Liao, H.J.; Chang, C.H.; Huang, C.F.; Chen, H.T. Potential of Using Infrapatellar-Fat-Pad-Derived Mesenchymal Stem Cells for Therapy in Degenerative Arthritis: Chondrogenesis, Exosomes, and Transcription Regulation. Biomolecules 2022, 12, 386. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, M.; Diederichs, S.; Melnik, S.; Riegger, J.; Trivanović, D.; Li, S.; Jenei-Lanzl, Z.; Brenner, R.E.; Huber-Lang, M.; Zaucke, F.; et al. Extracellular Vesicles in Musculoskeletal Pathologies and Regeneration. Front. Bioeng. Biotechnol. 2020, 8, 624096. [Google Scholar] [CrossRef] [PubMed]
- Duda, G.N.; Heller, M.O.; Pfitzner, T. AE-Manual der Endoprothetik—Knie; Springer: Berlin/Heidelberg, Germany, 2011; Volume 1. [Google Scholar] [CrossRef]
- Yao, B.; Samuel, L.T.; Acuña, A.J.; Faour, M.; Roth, A.; Kamath, A.F.; Mont, M.A. Infrapatellar Fat Pad Resection or Preservation during Total Knee Arthroplasty: A Systematic Review. J. Knee Surg. 2021, 34, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Afzali, M.F.; Radakovich, L.B.; Campbell, M.A.; Sanford, J.L.; Sykes, M.M.; Marolf, A.J.; Donahue, T.H.; Santangelo, K.S. Early removal of the infrapatellar fat pad beneficially alters the pathogenesis of moderate stage idiopathic knee osteoarthritis in the Dunkin Hartley guinea pig. bioRxiv 2022. [Google Scholar] [CrossRef]
- Bohnsack, M.; Wilharm, A.; Hurschler, C.; Rühmann, O.; Stukenborg-Colsman, C.; Wirth, C.J. Biomechanical and kinematic influences of a total infrapatellar fat pad resection on the knee. Am. J. Sports Med. 2004, 32, 1873–1880. [Google Scholar] [CrossRef] [PubMed]
- Lemon, M.; Packham, I.; Narang, K.; Craig, D.M. Patellar tendon length after knee arthroplasty with and without preservation of the infrapatellar fat pad. J. Arthroplast. 2007, 22, 574–580. [Google Scholar] [CrossRef]
- Chougule, S.S.; Stefanakis, G.; Stefan, S.C.; Rudra, S.; Tselentakis, G. Effects of fat pad excision on length of the patellar tendon after total knee replacement. J. Orthop. 2015, 12, 197–204. [Google Scholar] [CrossRef] [Green Version]
- Gandhi, R.; de Beer, J.; Leone, J.; Petruccelli, D.; Winemaker, M.; Adili, A. Predictive risk factors for stiff knees in total knee arthroplasty. J. Arthroplast. 2006, 21, 46–52. [Google Scholar] [CrossRef]
- Seo, J.G.; Lee, S.A.; Moon, Y.W.; Lee, B.H.; Ko, Y.H.; Chang, M.J. Infrapatellar fat pad preservation reduces wound complications after minimally invasive total knee arthroplasty. Arch. Orthop. Trauma Surg. 2015, 135, 1157–1162. [Google Scholar] [CrossRef]
- Maculé, F.; Sastre, S.; Lasurt, S.; Sala, P.; Segur, J.M.; Mallofré, C. Hoffa’s fat pad resection in total knee arthroplasty. Acta Orthop. Belg. 2005, 71, 714–717. [Google Scholar] [PubMed]
- Pinsornsak, P.; Naratrikun, K.; Chumchuen, S. The effect of infrapatellar fat pad excision on complications after minimally invasive TKA: A randomized controlled trial. Clin. Orthop. Relat. Res. 2014, 472, 695–701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meneghini, R.M.; Pierson, J.L.; Bagsby, D.; Berend, M.E.; Ritter, M.A.; Meding, J.B. The effect of retropatellar fat pad excision on patellar tendon contracture and functional outcomes after total knee arthroplasty. J. Arthroplast. 2007, 22, 47–50. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, N.; Sakahashi, H.; Sato, E.; Hirose, K.; Isima, T. Influence of the infrapatellar fat pad resection in a synovectomy during total knee arthroplasty in patients with rheumatoid arthritis. J. Arthroplast. 2003, 18, 897–902. [Google Scholar] [CrossRef]
- Nisar, S.; Lamb, J.N.; Somashekar, N.; Pandit, H.; van Duren, B.H. Preservation vs. resection of the infrapatellar fat pad during total knee arthroplasty part II: A systematic review of published evidence. Knee 2019, 26, 422–426. [Google Scholar] [CrossRef] [Green Version]
- White, L.; Holyoak, R.; Sant, J.; Hartnell, N.; Mullan, J. The effect of infrapatellar fat pad resection on outcomes post-total knee arthroplasty: A systematic review. Arch. Orthop. Trauma Surg. 2016, 136, 701–708. [Google Scholar] [CrossRef]
- Subramanyam, P.; Sundaram, P.S.; Rao, N. Scintigraphic assessment of patellar vascularity in total knee replacement surgeries following lateral release. Avicenna J. Med. 2012, 2, 54–59. [Google Scholar] [CrossRef]
- McMahon, M.S.; Scuderi, G.R.; Glashow, J.L.; Scharf, S.C.; Meltzer, L.P.; Scott, W.N. Scintigraphic determination of patellar viability after excision of infrapatellar fat pad and/or lateral retinacular release in total knee arthroplasty. Clin. Orthop. Relat. Res. 1990, 260, 10–16. [Google Scholar] [CrossRef]
- İmren, Y.; Dedeoğlu, S.S.; Çakar, M.; Çabuk, H.; Bayraktar, T.O.; Gürbüz, H. Infrapatellar Fat Pad Excision during Total Knee Arthroplasty Did Not Alter the Patellar Tendon Length: A 5-Year Follow-Up Study. J. Knee Surg. 2017, 30, 479–483. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Feature | Findings |
|---|---|
| Anatomy | macroscopic:
|
| Vascularisation | |
| Innervation | |
| Function | mechanic:
|
| Components | Pro-Inflammatory | Anti-Inflammatory |
|---|---|---|
| Cytokines/Mediators | IL-α [13,20] IL-1β [13,20,25] sIL-6R [13,19,20] TNF-α [13,19,20,25,45] IFNγ [13,19,25,45] IL-6 [13,19,20,25,45,46] IL-8 [13,19,20,25] IL-12 [19,25] IL-15 [20] IL-17 [45] IL-18 [20] IL-33 [20] IL-36α [20] IL-36β [20] IL-36γ [20] | IL-1Ra [19,20] IL-4 [19] IL-10 [19,45] IL-36Ra [20] IL-37 [20] IL-38 [20] Arg1 [19] PGE2 [19,20] PGF2α [25] |
| Adipokines | resistin [19,45] leptin [13,19,45,46] visfatin [13,19,45] FABP4 [19,46] WISP2 [19] chemerin [19] | adiponectin [13,19,45,46] |
| Growth factors | VEGF [13,19,20,46] FGF-2 [13] TGF-β [19,20,46] | FGF-2 [13] TGF-β [19,20,46] |
| Gene expression | PPARγ [19,46] | |
| Immune cells | M1-macrophages [13,19,25] CD4+ T-cells (Th1) [19,25] Mast cells [13,25,45] | M2-macrophages [13,19,25] CD8+ T-cells (Th2) [19,25] |
| Study | Primary Outcome | Results |
|---|---|---|
| Chougule et al. [69] | Length of patellar tendon | Shortening of patellar tendon after IFP-resection |
| İmren et al. [80] | Range of motion | No difference |
| Length of patellar tendon | No difference | |
| Lemon et al. [68] | Length of patellar tendon | Shortening of patellar tendon after IFP-resection |
| Maculé et al. [72] | Anterior knee pain | Decreased pain after IFP-resection |
| Range of motion | No difference | |
| Length of patellar tendon | No difference | |
| McMahon et al. [79] | Vascularization of the patella | No difference |
| Meneghini et al. [74] | Anterior knee pain | Increased pain after IFP-resection |
| Range of motion | No difference | |
| Length of patellar tendon | No difference | |
| Insall–Salvati ratio | No difference | |
| Knee society score | No difference | |
| Pinsornsak et al. [73] | Anterior knee pain | Increased pain after IFP-resection |
| Range of motion | No difference | |
| Length of patellar tendon | No difference | |
| Insall–Salvati ratio | No difference | |
| Knee society score | No difference | |
| Seo et al. [71] | Anterior knee pain | No difference |
| Subramanyam et al. [78] | Vascularization of the patella | Hypoperfusion of patella after IFP-resection |
| Tanaka et al. [75] | Anterior knee pain | Increased pain after IFP-resection |
| Range of motion | Decreased after IFP-resection | |
| Length of patellar tendon | Shortening of patellar tendon after IFP-resection | |
| Insall–Salvati ratio | Shortening of patellar tendon after IFP-resection | |
| Chougule et al. [69] | Length of patellar tendon | Shortening of patellar tendon after IFP-resection |
| İmren et al. [80] | Range of motion | No difference |
| Length of patellar tendon | No difference | |
| Lemon et al. [68] | Length of patellar tendon | Shortening of patellar tendon after IFP-resection |
| Maculé et al. [72] | Anterior knee pain | Decreased pain after IFP-resection |
| Range of motion | No difference | |
| Length of patellar tendon | No difference | |
| McMahon et al. [79] | Vascularization of the patella | No difference |
| Meneghini et al. [74] | Anterior knee pain | Increased pain after IFP-resection |
| Range of motion | No difference | |
| Length of patellar tendon | No difference | |
| Insall–Salvati ratio | No difference | |
| Knee society score | No difference | |
| Pinsornsak et al. [73] | Anterior knee pain | Increased pain after IFP-resection |
| Range of motion | No difference | |
| Length of patellar tendon | No difference | |
| Insall–Salvati ratio | No difference | |
| Knee society score | No difference | |
| Seo et al. [71] | Anterior knee pain | No difference |
| Subramanyam et al. [78] | Vascularization of the patella | Hypoperfusion of patella after IFP-resection |
| Tanaka et al. [75] | Anterior knee pain | Increased pain after IFP-resection |
| Range of motion | Decreased after IFP-resection | |
| Length of patellar tendon | Shortening of patellar tendon after IFP-resection | |
| Insall–Salvati ratio | Shortening of patellar tendon after IFP-resection |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Braun, S.; Zaucke, F.; Brenneis, M.; Rapp, A.E.; Pollinger, P.; Sohn, R.; Jenei-Lanzl, Z.; Meurer, A. The Corpus Adiposum Infrapatellare (Hoffa’s Fat Pad)—The Role of the Infrapatellar Fat Pad in Osteoarthritis Pathogenesis. Biomedicines 2022, 10, 1071. https://doi.org/10.3390/biomedicines10051071
Braun S, Zaucke F, Brenneis M, Rapp AE, Pollinger P, Sohn R, Jenei-Lanzl Z, Meurer A. The Corpus Adiposum Infrapatellare (Hoffa’s Fat Pad)—The Role of the Infrapatellar Fat Pad in Osteoarthritis Pathogenesis. Biomedicines. 2022; 10(5):1071. https://doi.org/10.3390/biomedicines10051071
Chicago/Turabian StyleBraun, Sebastian, Frank Zaucke, Marco Brenneis, Anna E. Rapp, Patrizia Pollinger, Rebecca Sohn, Zsuzsa Jenei-Lanzl, and Andrea Meurer. 2022. "The Corpus Adiposum Infrapatellare (Hoffa’s Fat Pad)—The Role of the Infrapatellar Fat Pad in Osteoarthritis Pathogenesis" Biomedicines 10, no. 5: 1071. https://doi.org/10.3390/biomedicines10051071
APA StyleBraun, S., Zaucke, F., Brenneis, M., Rapp, A. E., Pollinger, P., Sohn, R., Jenei-Lanzl, Z., & Meurer, A. (2022). The Corpus Adiposum Infrapatellare (Hoffa’s Fat Pad)—The Role of the Infrapatellar Fat Pad in Osteoarthritis Pathogenesis. Biomedicines, 10(5), 1071. https://doi.org/10.3390/biomedicines10051071