
Expression of the Costimulatory Molecule B7-H4 in the Decidua and Placental Tissues in Patients with Placental Abruption

 ,
,  ,
,
Abstract
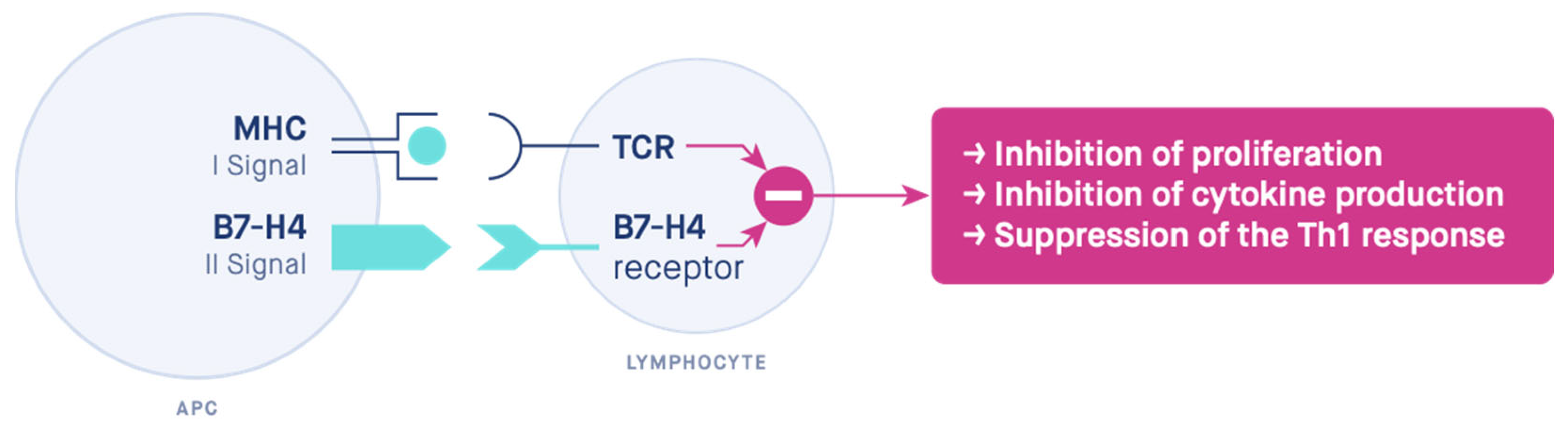
1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Tissue Samples and Immunohistochemistry
2.3. Statistical Analysis
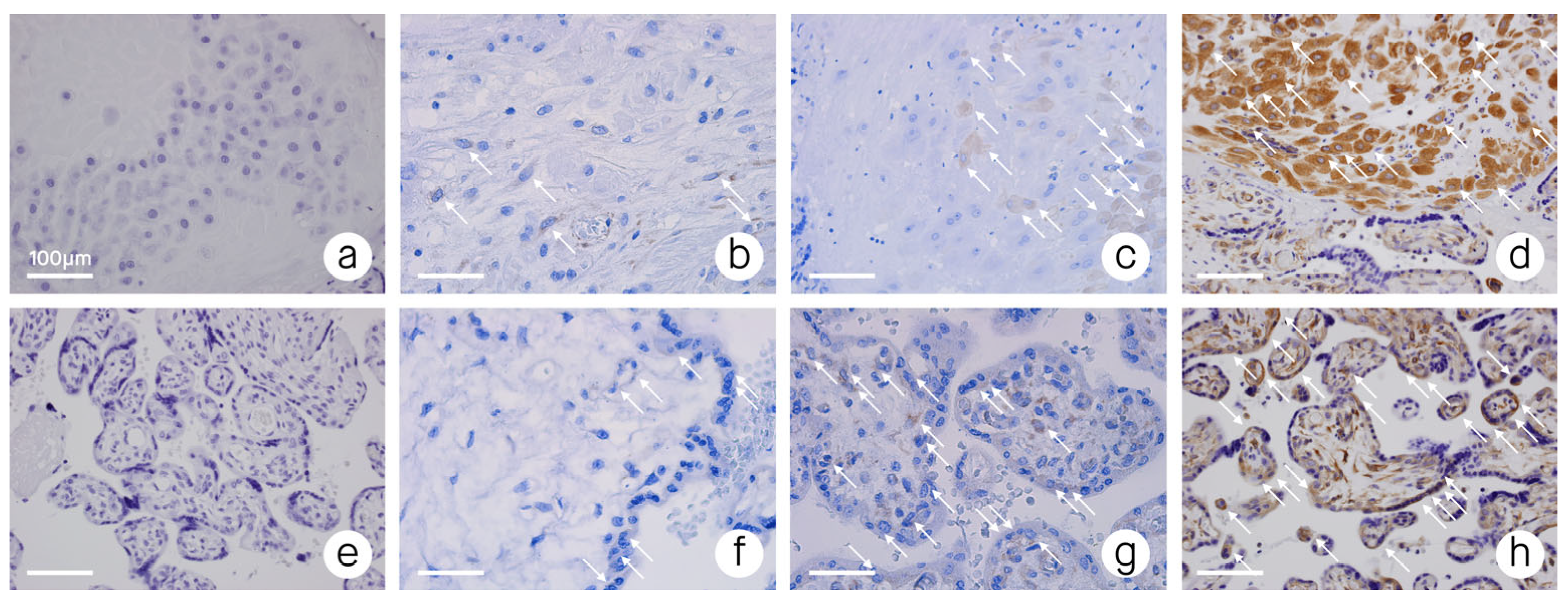
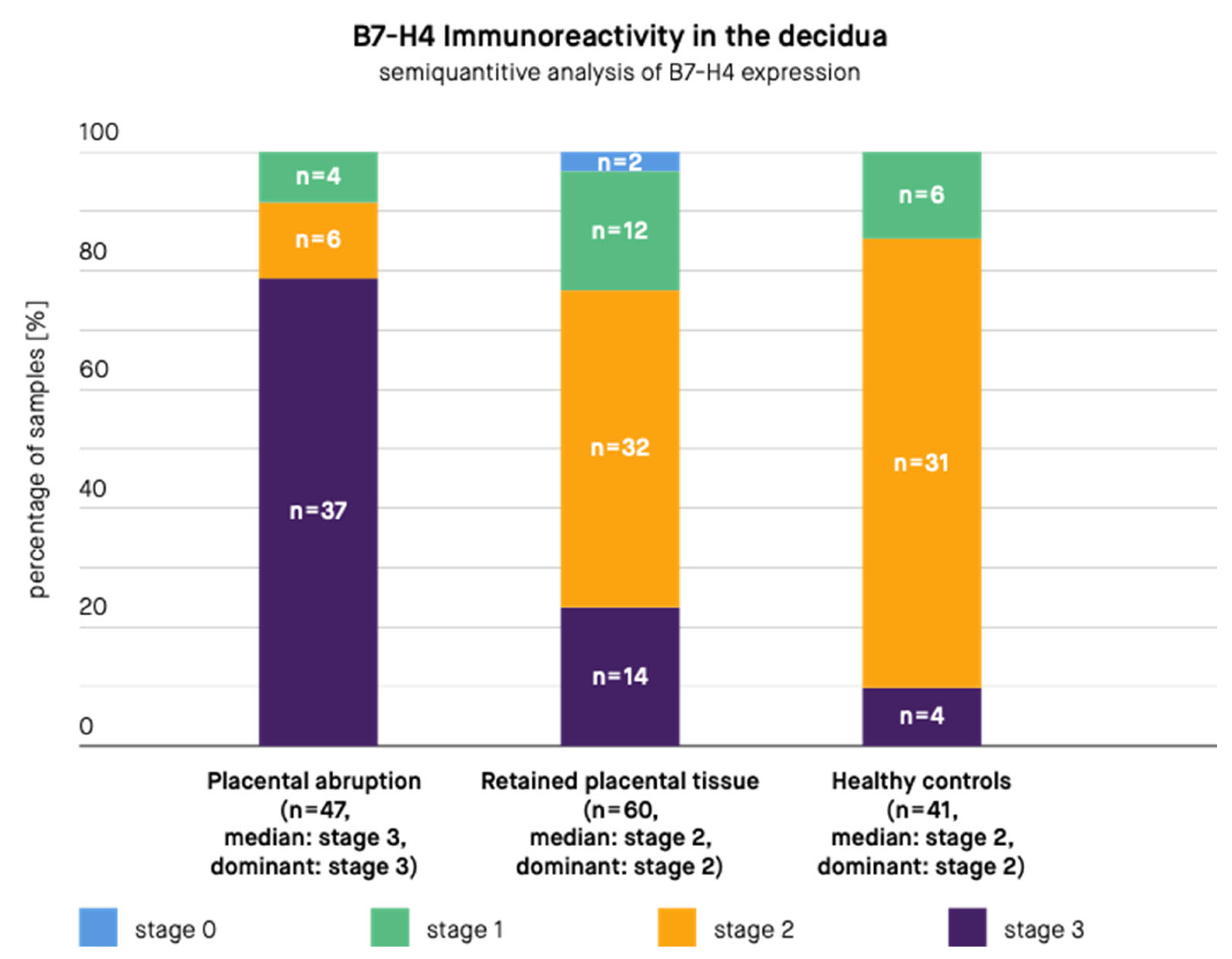
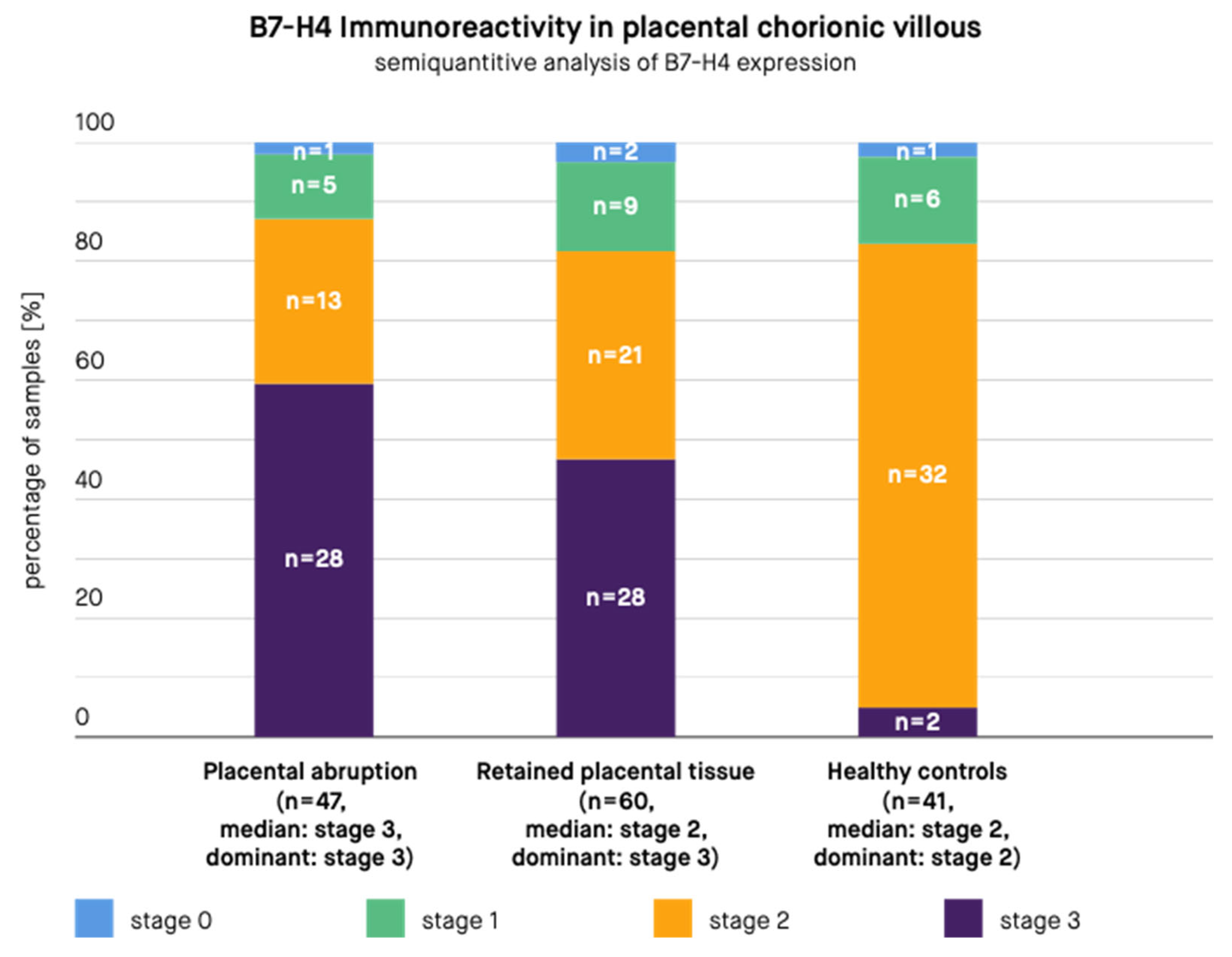
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wicherek, L.; Klimek, M.; Dutsch-Wicherek, M.; Kolodziejski, L.; Skotniczny, K. The molecular changes during placental detachment. Eur. J. Obs. Gynecol. Reprod. Biol. 2006, 125, 171–175. [Google Scholar] [CrossRef] [PubMed]
- Monin, L.; Whettlock, E.M.; Male, V. Immune responses in the human female reproductive tract. Immunology 2020, 160, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Wicherek, L. The role of the endometrium in the regulation of immune cell activity. Front. Biosci. 2008, 13, 1018–1035. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Morris, H.; Edwards, J.; Tiltman, A.; Emms, M. Endometrial lymphoid tissue: An immunohistological study. J. Clin. Pathol. 1985, 38, 644–652. [Google Scholar] [CrossRef]
- Wilczynski, J.R.; Tchórzewski, H.; Banasik, M.; Głowacka, E.; Wieczorek, A.; Lewkowicz, P.; Malinowski, A.; Szpakowski, M.; Wilczyński, J. Lymphocyte subset distribution and cytokine secretion in third trimester decidua in normal pregnancy and preeclampsia. Eur. J. Obstet. Gynecol. Reprod. Biol. 2003, 109, 8–15. [Google Scholar] [CrossRef]
- Galazka, K.; Pitynski, K.; Skret-Magierlo, J.; Mach, P.; Knafel, A.; Sikora, J.; Niemiec, T.; Dobrogowski, J.; Basta, A.; Wicherek, L. The Increase in Metallothionein and Ectopic Decidual Immunoreactivity with Respect to the Progression of Labor at Term and the Lack of Analogical Changes in Placental Abruption. Am. J. Reprod. Immunol. 2008, 60, 204–213. [Google Scholar] [CrossRef]
- King, A.; Burrows, T.; Loke, Y.W. Human uterine natural killer cells. Nat. Immunol. 1996, 15, 41–52. [Google Scholar]
- Watanabe, H.; Miyaji, C.; Kawachi, Y.; Iiai, T.; Ohtsuka, K.; Iwanage, T.; Takahashi-Iwanaga, H.; Abo, T. Relationships between intermediate TCR cells and NK1.1+ T cells in various immune organs. NK1.1+ T cells are present within a population of intermediate TCR cells. J. Immunol. 1995, 155, 2972–2983. [Google Scholar]
- Aluvihare, V.R.; Kallikourdis, M.; Betz, A.G. Regulatory T cells mediate maternal tolerance to the fetus. Nat. Immunol. 2004, 5, 266–271. [Google Scholar] [CrossRef]
- Saito, S.; Sasaki, Y.; Sakai, M. CD4(+)CD25high regulatory T cells in human pregnancy. J. Reprod. Immunol. 2005, 65, 111–120. [Google Scholar] [CrossRef]
- Jacek, R.W.; Wilczynski, J.R.; Kalinka, J.; Radwan, M. The role of T-regulatory cells in pregnancy and cancer. Front. Biosci. 2008, 13, 2275–2289. [Google Scholar] [CrossRef]
- Wicherek, L.; Galazka, K.; Lazar, A. RCAS1 Decidual Immunoreactivity During Placental Abruption: Immune Cell Presence and Activity. Am. J. Reprod. Immunol. 2007, 58, 46–55. [Google Scholar] [CrossRef]
- Ledee-Bataille, N.; Bonnet-Chea, K.; Hosny, G.; Dubanchet, S.; Frydman, R.; Chaouat, G. Role of the endometrial tripod interleukin-18, -15, and -12 in inadequate uterine receptivity in pa-tients with a history of repeated in vitro fertilization-embryo transfer failure. Fertil. Steril. 2005, 83, 598–605. [Google Scholar] [CrossRef]
- Dimitriadis, E.; Stoikos, C.; Stafford-Bell, M.; Clark, L.; Paiva, P.; Kovacs, G.; Salamonsen, L.A. Interleukin-11, IL-11 receptoralpha and leukemia inhibitory factor are dysregulated in endometrium of infertile women with endometriosis during the implantation window. J. Reprod. Immunol. 2006, 69, 53–64. [Google Scholar] [CrossRef]
- Dmowski, W.P.; Ding, J.; Shen, J.; Rana, N.; Fernandez, B.; Braun, D. Apoptosis in endometrial glandular and stromal cells in women with and without endometriosis. Hum. Reprod. 2001, 16, 1802–1808. [Google Scholar] [CrossRef]
- Beliard, A.; Noel, A.; Foidart, J.-M. Reduction of apoptosis and proliferation in endometriosis. Fertil. Steril. 2004, 82, 80–85. [Google Scholar] [CrossRef]
- Harlev, A.; Levy, A.; Zaulan, Y.; Koifman, A.; Mazor, M.; Wiznitzer, A.; Faizayev, E.; Sheiner, E. Idiopathic bleeding during the second half of pregnancy as a risk factor for adverse perinatal outcome. J. Matern. Fetal. Neonatal Med. 2008, 21, 331–335. [Google Scholar] [CrossRef]
- Koifman, A.; Levy, A.; Zaulan, Y.; Harlev, A.; Mazor, M.; Wiznitzer, A.; Sheiner, E. The clinical significance of bleeding during the second trimester of pregnancy. Arch. Gynecol. Obstet. 2008, 278, 47–51. [Google Scholar] [CrossRef]
- Cunningham, J.W. Prompt evaluation and treatment of third-trimester bleeding. J. Am. Acad. Physician Assist. 2021, 34, 26–31. [Google Scholar] [CrossRef]
- Ananth, C.V.; Wilcox, A.J. Placental Abruption and Perinatal Mortality in the United States. Am. J. Epidemiol. 2001, 153, 332–337. [Google Scholar] [CrossRef]
- Tikkanen, M. Placental abruption: Epidemiology, risk factors and consequences. Acta Obstet. Gynecol. Scand. 2011, 90, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Ananth, C.V.; Smulian, J.C.; Vintzileos, A.M. Ischemic placental disease: Maternal versus fetal clinical presentations by gestational age. J. Matern. Fetal. Neonatal Med. 2010, 23, 887–893. [Google Scholar] [CrossRef] [PubMed]
- Ananth, C.V. Ischemic placental disease: A unifying concept for preeclampsia, intrauterine growth restriction, and placental abruption. Semin. Perinatol. 2014, 38, 131–132. [Google Scholar] [CrossRef] [PubMed]
- Ananth, C.V.; Peltier, M.R.; Chavez, M.R.; Kirby, R.S.; Getahun, D.; Vintzileos, A.M. Recurrence of Ischemic Placental Disease. Obstet. Gynecol. 2007, 110, 128–133. [Google Scholar] [CrossRef]
- Dommisse, J.; Tiltman, A.J. Placental bed biopsies in placental abruption. BJOG Int. J. Obstet. Gynaecol. 1992, 99, 651–654. [Google Scholar] [CrossRef]
- Signore, C.; Mills, J.L.; Qian, C.; Yu, K.; Lam, C.; Epstein, F.H.; Karumanchi, S.A.; Levine, R.J. Circulating Angiogenic Factors and Placental Abruption. Obstet. Gynecol. 2006, 108, 338–344. [Google Scholar] [CrossRef]
- Geldenhuys, J.; Rossouw, T.M.; Lombaard, H.A.; Ehlers, M.M.; Kock, M.M. Disruption in the Regulation of Immune Responses in the Placental Subtype of Preeclampsia. Front. Immunol. 2018, 9, 1659. [Google Scholar] [CrossRef]
- Salafia, C.M.; López-Zeno, J.A.; Sherer, D.M.; Whittington, S.S.; Minior, V.K.; Vintzileos, A.M. Histologic evidence of old intrauterine bleeding is more frequent in prematurity. Am. J. Obstet. Gynecol. 1995, 173, 1065–1070. [Google Scholar] [CrossRef]
- Nakatsuka, M.; Asagiri, K.; Kimura, Y.; Kamada, Y.; Tada, K.; Kudo, T. Generation of peroxynitrite and apoptosis in placenta of patients with chorioamnionitis: Possible impli-cations in placental abruption. Hum. Reprod. 1999, 14, 1101–1106. [Google Scholar] [CrossRef]
- Ananth, C.V.; Oyelese, Y.; Srinivas, N.; Yeo, L.; Vintzileos, A.M. Preterm premature rupture of membranes, intrauterine infection, and oligohydramnios: Risk factors for placental abruption. Obstet. Gynecol. 2004, 104, 71–77. [Google Scholar] [CrossRef]
- Avagliano, L.; Falleni, M.; Marconi, A.M.; Bulfoni, C.; Prada, A.; Barbera, A.F.; Doi, P.; Bulfamante, G.P. An imbalance of COX level is not related to placental abruption. J. Clin. Pathol. 2011, 64, 605–609. [Google Scholar] [CrossRef]
- Darby, M.J.; Caritis, S.; Shen-Schwarz, S. Placental abruption in the preterm gestation: An association with chorioamnionitis. Obstet. Gynecol. 1989, 74, 88–92. [Google Scholar]
- Rana, A.; Sawhney, H.; Gopalan, S.; Panigrahi, D.; Nijhawan, R. Abruptio placentae and chorioamnionitis-microbiological and histologic correlation. Acta. Obstet. Gynecol. Scand. 1999, 78, 363–366. [Google Scholar]
- Balkundi, D.R.; Hanna, N.; Hileb, M.; Dougherty, J.; Sharma, S. Labor-Associated Changes in Fas Ligand Expression and Function in Human Placenta. Pediatr. Res. 2000, 47, 301–308. [Google Scholar] [CrossRef][Green Version]
- Ananth, C.V.; Oyelese, Y.; Prasad, V.; Getahun, D.; Smulian, J.C. Evidence of placental abruption as a chronic process: Associations with vaginal bleeding early in pregnancy and placental lesions. Eur. J. Obstet. Gynecol. Reprod. Biol. 2006, 128, 15–21. [Google Scholar] [CrossRef]
- Nath, C.A.; Ananth, C.V.; Smulian, J.C.; Shen-Schwarz, S.; Kaminsky, L.; New Jersey–Placental Abruption Study Investigators. Histologic evidence of inflammation and risk of placental abruption. Am. J. Obstet. Gynecol. 2007, 197, 319.e1-6. [Google Scholar] [CrossRef]
- Harger, J.H.; Hsing, A.W.; Tuomala, R.E.; Gibbs, R.S.; Mead, P.B.; Eschenbach, D.A.; Knox, G.E.; Polk, B.F. Risk factors for preterm premature rupture of fetal membranes: A multicenter case-control study. Am. J. Obstet. Gynecol. 1990, 163, 130–137. [Google Scholar] [CrossRef]
- Williams, M.A.; Mittendorf, R.; Lieberman, E.; Monson, R.R. Adverse infant outcomes associated with first-trimester vaginal bleeding. Obstet. Gynecol. 1991, 78, 14–18. [Google Scholar]
- Lockwood, C.J.; Toti, P.; Arcuri, F.; Paidas, M.; Buchwalder, L.; Krikun, G.; Schatz, F. Mechanisms of Abruption-Induced Premature Rupture of the Fetal Membranes: Thrombin-Enhanced Interleukin-8 Expression in Term Decidua. Am. J. Pathol. 2005, 167, 1443–1449. [Google Scholar] [CrossRef]
- Oyelese, Y.; Ananth, C.V. Placental abruption. Obstet. Gynecol. 2006, 108, 1005–1016. [Google Scholar] [CrossRef]
- Huang, S.-T.J.; Schatz, F.; Salafia, C.; Stocco, C.; Lockwood, C.J.; Krikun, G. Thrombin activation of endometrial endothelial cells: A possible role in intrauterine growth restriction. Thromb. Haemost. 2007, 97, 245–253. [Google Scholar] [CrossRef]
- Brosens, I.; Pijnenborg, R.; Vercruysse, L.; Romero, R. The “Great Obstetrical Syndromes” are associated with disorders of deep placentation. Am. J. Obstet. Gynecol. 2011, 204, 193–201. [Google Scholar] [CrossRef]
- Zdoukopoulos, N.; Zintzaras, E. Genetic risk factors for placental abruption: A HuGE review and meta-analysis. Epidemiology 2008, 19, 309–323. [Google Scholar] [CrossRef]
- Workalemahu, T.; Enquobahrie, D.A.; Gelaye, B.; Sanchez, S.E.; Garcia, P.J.; Tekola-Ayele, F.; Hajat, A.; Thornton, T.A.; Ananth, C.V.; Williams, M.A. Genetic variations and risk of placental abruption: A genome-wide association study and meta-analysis of genome-wide association studies. Placenta 2018, 66, 8–16. [Google Scholar] [CrossRef]
- Workalemahu, T.; Enquobahrie, D.A.; Gelaye, B.; Thornton, T.A.; Tekola-Ayele, F.; Sanchez, S.E.; Garcia, P.J.; Palomino, H.G.; Hajat, A.; Romero, R.; et al. Abruptio placentae risk and genetic variations in mitochondrial biogenesis and oxidative phosphory-lation: Replication of a candidate gene association study. Am. J. Obstet. Gynecol. 2018, 219, 617.e1–617.e17. [Google Scholar] [CrossRef]
- Hemminki, E.; Merilainen, J. Long-term effects of cesarean sections: Ectopic pregnancies and placental problems. Am. J. Obstet. Gynecol. 1996, 174, 1569–1574. [Google Scholar] [CrossRef]
- Skręt-Magierło, J.E.; Wicherek, L.; Basta, P.; Galazka, K.; Sikora, J.; Wilk, M.; Fudali, L.; Skret, A. RCAS1 Decidual Immunoreactivity during Cesarean Section in Scar Deciduosis: Immune Cell Presence and Activity. Gynecol. Obstet. Investig. 2007, 65, 187–194. [Google Scholar] [CrossRef]
- Wicherek, L.; Basta, P.; Galazka, K.; Mak, P.; Dancewicz, L.; Kalinka, J. ORIGINAL ARTICLE: RCAS1 Decidual Immunoreactivity and RCAS1 Serum Level During Cesarean Section with Respect to the Progression of Labor. Am. J. Reprod. Immunol. 2008, 59, 152–158. [Google Scholar] [CrossRef]
- Choi, I.-H.; Zhu, G.; Sica, G.L.; Strome, S.E.; Cheville, J.C.; Lau, J.S.; Zhu, Y.; Flies, D.B.; Tamada, K.; Chen, L. Genomic Organization and Expression Analysis of B7-H4, an Immune Inhibitory Molecule of the B7 Family. J. Immunol. 2003, 171, 4650–4654. [Google Scholar] [CrossRef]
- Petroff, M.G.; Perchellet, A. B7 Family Molecules as Regulators of the Maternal Immune System in Pregnancy. Am. J. Reprod. Immunol. 2010, 63, 506–519. [Google Scholar] [CrossRef]
- Mach, P.; Gellhaus, A.; Wicherek, L.; Schmidt, B.; Kimmig, R.; Kasimir-Bauer, S.; Köninger, A. Changes in the Blood Serum Levels of the Costimulatory Soluble B7-H4 Molecule in Pregnant Women During the Peripartal Phase. Am. J. Reprod. Immunol. 2015, 74, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Mach, P.; Köninger, A.; Wicherek, L.; Kimmig, R.; Kasimir-Bauer, S.; Birdir, C.; Schmidt, B.; Gellhaus, A. Serum concentrations of soluble B7-H4 in early pregnancy are elevated in women with preterm premature rupture of fetal membranes. Am. J. Reprod. Immunol. 2016, 76, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Mach, P.; Nolte-Boenigk, L.; Droste, L.; Fox, L.; Frank, M.; Schmidt, B.; Herse, F.; Verlohren, S.; Wicherek, L.; Iannaccone, A.; et al. Soluble B7-H4 blood serum levels are elevated in women at high risk for preeclampsia in the first trimester, as well as in patients with confirmed preeclampsia. Am. J. Reprod. Immunol. 2018, 80, e12988. [Google Scholar] [CrossRef] [PubMed]
- Greenwald, R.J.; Freeman, G.J.; Sharpe, A.H. The B7 family revisited. Annu. Rev. Immunol. 2005, 23, 515–548. [Google Scholar] [CrossRef]
- Zhao, Y.; Zheng, Q.; Jin, L. The Role of B7 Family Molecules in Maternal–Fetal Immunity. Front. Immunol. 2020, 11, 458. [Google Scholar] [CrossRef]
- Prasad, D.V.; Richards, S.; Mai, X.M.; Dong, C. B7S1, a Novel B7 Family Member that Negatively Regulates T Cell Activation. Immunity 2003, 18, 863–873. [Google Scholar] [CrossRef]
- Sica, G.L.; Choi, I.-H.; Zhu, G.; Tamada, K.; Wang, S.-D.; Tamura, H.; Chapoval, A.I.; Flies, D.B.; Bajorath, J.; Chen, L. B7-H4, a Molecule of the B7 Family, Negatively Regulates T Cell Immunity. Immunity 2003, 18, 849–861. [Google Scholar] [CrossRef]
- Zang, X.; Loke, P.; Kim, J.; Murphy, K.; Waitz, R.; Allison, J.P. B7x: A widely expressed B7 family member that inhibits T cell activation. Proc. Natl. Acad. Sci. USA 2003, 100, 10388–10392. [Google Scholar] [CrossRef]
- Miyatake, T.; Tringler, B.; Liu, W.; Liu, S.H.; Papkoff, J.; Enomoto, T.; Torkkoa, K.C.; Dehna, D.L.; Swishera, A.; Shroyer, K.R.; et al. B7-H4 (DD-O110) is overexpressed in high risk uterine endometrioid adenocarcinomas and inversely cor-related with tumor T-cell infiltration. Gynecol. Oncol. 2007, 106, 119–127. [Google Scholar] [CrossRef]
- Park, G.B.; Song, H.; Kim, Y.-S.; Sung, M.; Ryu, J.W.; Lee, H.-K.; Cho, D.H.; Kim, D.; Lee, W.J.; Hur, D.Y.; et al. Cell cycle arrest induced by engagement of B7-H4 on Epstein-Barr virus-positive B-cell lymphoma cell lines. Immunology 2009, 128, 360–368. [Google Scholar] [CrossRef]
- MacGregor, H.L.; Ohashi, P.S. Molecular Pathways: Evaluating the Potential for B7-H4 as an Immunoregulatory Target. Clin. Cancer Res. 2017, 23, 2934–2941. [Google Scholar] [CrossRef]
- Kaur, G.; Janakiram, M. B7x-from bench to bedside. ESMO Open 2019, 4, e000554. [Google Scholar] [CrossRef]
- Suh, W.-K.; Wang, S.; Duncan, G.S.; Miyazaki, Y.; Cates, T.; Walker, T.; Gajewska, B.U.; Deenick, E.; Dawicki, W.; Okada, H.; et al. Generation and characterization of B7-H4/B7S1/B7x-deficient mice. Mol. Cell. Biol. 2006, 26, 6403–6411. [Google Scholar] [CrossRef]
- Zou, W.; Chen, L. Inhibitory B7-family molecules in the tumour microenvironment. Nat. Rev. Immunol. 2008, 8, 467–477. [Google Scholar] [CrossRef]
- Ahangar, N.K.; Hemmat, N.; Khalaj-Kondori, M.; Shadbad, M.A.; Sabaie, H.; Mokhtarzadeh, A.; Alizadeh, N.; Derakhshani, A.; Baghbanzadeh, A.; Dolatkhah, K.; et al. The Regulatory Cross-Talk between microRNAs and Novel Members of the B7 Family in Human Diseases: A Scoping Review. Int. J. Mol. Sci. 2021, 22, 2652. [Google Scholar] [CrossRef]
- Petroff, M.G.; Chen, L.; Phillips, T.A.; Azzola, D.; Sedlmayr, P.; Hunt, J.S. B7 Family Molecules Are Favorably Positioned at the Human Maternal-Fetal Interface1. Biol. Reprod. 2003, 68, 1496–1504. [Google Scholar] [CrossRef]
- Tringler, B.; Zhuo, S.; Pilkington, G.; Torkko, K.C.; Singh, M.; Lucia, M.S.; Heinz, D.E.; Papkoff, J.; Shroyer, K.R. B7-H4 Is Highly Expressed in Ductal and Lobular Breast Cancer. Clin. Cancer Res. 2005, 11, 1842–1848. [Google Scholar] [CrossRef]
- Christiaens, I.; Zaragoza, D.B.; Guilbert, L.; Robertson, S.; Mitchell, B.F.; Olson, D.M. Inflammatory processes in preterm and term parturition. J. Reprod. Immunol. 2008, 79, 50–57. [Google Scholar] [CrossRef]
- Repnik, U.; Tilburgs, T.; Roelen, D.; van der Mast, B.; Kanhai, H.; Scherjon, S.; Claas, F. Comparison of Macrophage Phenotype Between Decidua Basalis and Decidua Parietalis by Flow Cytometry. Placenta 2008, 29, 405–412. [Google Scholar] [CrossRef]
- Azuma, T.; Zhu, G.; Xu, H.; Rietz, A.C.; Drake, C.G.; Matteson, E.L.; Chen, L. Potential Role of Decoy B7-H4 in the Pathogenesis of Rheumatoid Arthritis: A Mouse Model Informed by Clinical Data. PLoS Med. 2009, 6, e1000166. [Google Scholar] [CrossRef]
- Kamimura, Y.; Kobori, H.; Piao, J.; Hashiguchi, M.; Matsumoto, K.; Hirose, S.; Azuma, M. Possible involvement of soluble B7-H4 in T cell-mediated inflammatory immune responses. Biochem. Biophys. Res. Commun. 2009, 389, 349–353. [Google Scholar] [CrossRef]
- Lappas, M. Visfatin regulates the terminal processes of human labour and delivery via activation of the nuclear factor-kappaB pathway. Mol. Cell. Endocrinol. 2012, 348, 128–134. [Google Scholar] [CrossRef]
- Zhang, L.; Wu, H.; Lu, D.; Li, G.; Sun, C.; Song, H.; Li, J.; Zhai, T.; Huang, L.; Hou, C.; et al. The costimulatory molecule B7-H4 promote tumor progression and cell proliferation through translocating into nucleus. Oncogene 2013, 32, 5347–5358. [Google Scholar] [CrossRef]
- Simon, I.; Zhuo, S.; Corral, L.; Diamandis, E.P.; Sarno, M.J.; Wolfert, R.L.; Kim, N.W. B7-H4 Is a Novel Membrane-Bound Protein and a Candidate Serum and Tissue Biomarker for Ovarian Cancer. Cancer Res. 2006, 66, 1570–1575. [Google Scholar] [CrossRef]
- Thompson, R.H.; Zang, X.; Lohse, C.M.; Leibovich, B.C.; Slovin, S.F.; Reuter, V.E.; Cheville, J.C.; Blute, M.L.; Russo, P.; Kwon, E.D.; et al. Serum-Soluble B7x Is Elevated in Renal Cell Carcinoma Patients and Is Associated with Advanced Stage. Cancer Res. 2008, 68, 6054–6058. [Google Scholar] [CrossRef]
- Leandersson, P.; Kalapotharakos, G.; Henic, E.; Borgfeldt, H.; Petzold, M.; Høyer-Hansen, G.; Borgfeldt, C. A Biomarker Panel Increases the Diagnostic Performance for Epithelial Ovarian Cancer Type I and II in Young Women. Anticancer Res. 2016, 36, 957–965. [Google Scholar]
- Jiang, X.; Liu, G.; Li, Y.; Pan, Y. Immune checkpoint: The novel target for antitumor therapy. Genes Dis. 2019, 8, 25–37. [Google Scholar] [CrossRef]
- Podojil, J.R.; Chiang, M.Y.; Ifergan, I.; Copeland, R.; Liu, L.N.; Maloveste, S.; Miller, S.D. B7-H4 Modulates Regulatory CD4(+) T Cell Induction and Function via Ligation of a Semaphorin 3a/Plexin A4/Neuropilin-1 Complex. J. Immunol. 2018, 201, 897–907. [Google Scholar] [CrossRef] [PubMed]
- Yi, K.H.; Chen, L. Fine tuning the immune response through B7-H3 and B7-H4. Immunol. Rev. 2009, 229, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Galazka, K.; Wicherek, L.; Pitynski, K.; Kijowski, J.; Zajac, K.; Bednarek, W.; Dutsch-Wicherek, M.; Rytlewski, K.; Kalinka, J.; Basta, A.; et al. Changes in the subpopulation of CD25+ CD4+ and FOXP3+ regulatory T cells in decidua with respect to the progression of labor at term and the lack of analogical changes in the subpopulation of suppressive B7-H4 macrophages—A preliminary report. Am. J. Reprod. Immunol. 2009, 61, 136–146. [Google Scholar] [CrossRef]
- Darmochwal-Kolarz, D.; Kludka-Sternik, M.; Kolarz, B.; Chmielewski, T.; Tabarkiewicz, J.; Rolinski, J.; Leszczynska-Gorzelak, B.; Oleszczuk, J. The expression of B7-H1 and B7-H4 co-stimulatory molecules on myeloid and plasmacytoid dendritic cells in pre-eclampsia and normal pregnancy. J. Reprod. Immunol. 2013, 99, 33–38. [Google Scholar] [CrossRef]
- Duan, L.; Reisch, B.; Iannaccone, A.; Hadrovic, E.; Wu, Y.; Vogtmann, R.; Winterhager, E.; Kimmig, R.; Köninger, A.; Mach, P.; et al. Abnormal expression of the costimulatory molecule B7-H4 in placental chorionic villous and decidual basalis tissues of patients with preeclampsia and HELLP syndrome. Am. J. Reprod. Immunol. 2021, 86, e13430. [Google Scholar] [CrossRef]
- Prelabor Rupture of Membranes: ACOG Practice Bulletin, Number 217. Obstet. Gynecol. 2020, 135, e80–e97. [CrossRef]
- Darmochwal-Kolarz, D.; Kludka-Sternik, M.; Chmielewski, T.; Kolarz, B.; Rolinski, J.; Oleszczuk, J. PP069. The expressions of B7-H1 and B7-H4 co-stimulatory molecules on myeloid and lymphoid dendritic cells in pre-eclampsia and normal pregnancy. The expressions of B7-H1 and B7-H4 co-stimulatory moleculeson myeloid and lymphoid dendritic cells in pre-eclampsia and normal pregnancy. Pregnancy Hypertens. Int. J. Women’s Cardiovasc. Health 2012, 2, 278–279. [Google Scholar] [CrossRef]
- Gestational Hypertension and Preeclampsia: ACOG Practice Bulletin, Number 222. Obstet. Gynecol. 2020, 135, e237–e260. [CrossRef]
- Koga, K.; Aldo, P.B.; Mor, G. Toll-like receptors and pregnancy: Trophoblast as modulators of the immune response. J. Obstet. Gynaecol. Res. 2009, 35, 191–202. [Google Scholar] [CrossRef]
- Sasaki, Y.; Darmochwal-Kolarz, D.; Suzuki, D.; Sakai, M.; Ito, M.; Shima, T.; Shiozaki, A.; Rolinski, J.; Saito, S. Proportion of peripheral blood and decidual CD4+ CD25bright regulatory T cells in pre-eclampsia. Clin. Exp. Immunol. 2007, 149, 139–145. [Google Scholar] [CrossRef]
- Steinborn, A.; Haensch, G.M.; Mahnke, K.; Schmitt, E.; Toermer, A.; Meuer, S.; Sohn, C. Distinct subsets of regulatory T cells during pregnancy: Is the imbalance of these subsets involved in the pathogenesis of preeclampsia? Clin. Immunol. 2008, 129, 401–412. [Google Scholar] [CrossRef]
- Santner-Nanan, B.; Peek, M.J.; Khanam, R.; Richarts, L.; Zhu, E.; Groth, B.F.D.S.; Nanan, R. Systemic Increase in the Ratio between Foxp3+and IL-17-Producing CD4+T Cells in Healthy Pregnancy but Not in Preeclampsia. J. Immunol. 2009, 183, 7023–7030. [Google Scholar] [CrossRef]
- Redman, C.W.G.; Sargent, I.L. Immunology of Pre-Eclampsia. Am. J. Reprod. Immunol. 2010, 63, 534–543. [Google Scholar] [CrossRef]
- Saito, S.; Nakashima, A.; Shima, T.; Ito, M. Th1/Th2/Th17 and regulatory T-cell paradigm in pregnancy. Am. J. Reprod. Immunol. 2010, 63, 601–610. [Google Scholar] [CrossRef] [PubMed]
- Tikkanen, M.; Nuutila, M.; Hiilesmaa, V.; Paavonen, J.; Ylikorkala, O. Clinical presentation and risk factors of placental abruption. Acta Obstet. Gynecol. Scand. 2006, 85, 700–705. [Google Scholar] [CrossRef] [PubMed]
- Nyberg, D.; Cyr, D.R.; Mack, L.; Wilson, D.; Shuman, W. Sonographic spectrum of placental abruption. Am. J. Roentgenol. 1987, 148, 161–164. [Google Scholar] [CrossRef] [PubMed]
- Page, E.W.; King, E.B.; Merrill, J.A. Abruptio placentae; dangers of delay in delivery. Obstet. Gynecol. 1954, 3, 385–393. [Google Scholar] [PubMed]
- Hurd, W.W.; Miodovnik, M.; Hertzberg, V.; Lavin, J.P. Selective management of abruptio placentae: A prospective study. Obstet. Gynecol. 1983, 61, 467–473. [Google Scholar] [PubMed]
- Wicherek, L.; Basta, P.; Sikora, J.; Galazka, K.; Rytlewski, K.; Grabiec, M.; Lazar, A.; Kalinka, J. RCAS1 Decidual Immunoreactivity in Severe Pre-Eclampsia: Immune Cell Presence and Activity. Am. J. Reprod. Immunol. 2007, 58, 358–366. [Google Scholar] [CrossRef] [PubMed]
- Menon, R.; Bonney, E.A.; Condon, J.; Mesiano, S.; Taylor, R.N. Novel concepts on pregnancy clocks and alarms: Redundancy and synergy in human parturition. Hum. Reprod. Updat. 2016, 22, 535–560. [Google Scholar] [CrossRef]
- Bączkowska, M.; Zgliczyńska, M.; Faryna, J.; Przytuła, E.; Nowakowski, B.; Ciebiera, M. Molecular Changes on Maternal–Fetal Interface in Placental Abruption—A Systematic Review. Int. J. Mol. Sci. 2021, 22, 6612. [Google Scholar] [CrossRef]
- Lockwood, C.J.; Stocco, C.; Murk, W.; Kayisli, U.A.; Funai, E.F.; Schatz, F. Human Labor Is Associated with Reduced Decidual Cell Expression of Progesterone, But Not Glucocorticoid, Receptors. J. Clin. Endocrinol. Metab. 2010, 95, 2271–2275. [Google Scholar] [CrossRef]
- Lockwood, C.J.; Kayisli, U.A.; Stocco, C.; Murk, W.; Vatandaslar, E.; Buchwalder, L.F.; Schatz, F. Abruption-Induced Preterm Delivery Is Associated with Thrombin-Mediated Functional Progesterone Withdrawal in Decidual Cells. Am. J. Pathol. 2012, 181, 2138–2148. [Google Scholar] [CrossRef]
- Guzeloglu-Kayisli, O.; Kayisli, U.A.; Semerci, N.; Basar, M.; Buchwalder, L.F.; Buhimschi, C.S.; Lockwood, C.J. Mechanisms of chorioamnionitis-associated preterm birth: Interleukin-1beta inhibits progesterone receptor expression in decidual cells. J. Pathol. 2015, 237, 423–434. [Google Scholar] [CrossRef]
- Ackerman, W.E., IV; Summerfield, T.L.; Mesiano, S.; Schatz, F.; Lockwood, C.J.; Kniss, D.A. Agonist-Dependent Downregulation of Progesterone Receptors in Human Cervical Stromal Fibro-blasts. Reprod. Sci. 2016, 23, 112–123. [Google Scholar] [CrossRef][Green Version]
- Nadeem, L.; Shynlova, O.; Matysiak-Zablocki, E.; Mesiano, S.; Dong, X.; Lye, S. Molecular evidence of functional progesterone withdrawal in human myometrium. Nat. Commun. 2016, 7, 11565. [Google Scholar] [CrossRef]
- Patel, B.; Peters, G.A.; Skomorovska-Prokvolit, Y.; Yi, L.; Tan, H.; Yousef, A.; Wang, J.; Mesiano, S. Control of Progesterone Receptor-A Transrepressive Activity in Myometrial Cells: Implications for the Control of Human Parturition. Reprod. Sci. 2018, 25, 214–221. [Google Scholar] [CrossRef]
- Norwitz, E.R.; Snegovskikh, V.; Schatz, F.; Foyouzi, N.; Rahman, M.; Buchwalder, L.; Lee, H.J.; Funai, E.F.; Buhimschi, C.S.; Buhimschi, I.A.; et al. Progestin inhibits and thrombin stimulates the plasminogen activator/inhibitor system in term decidual stromal cells: Implications for parturition. Am. J. Obstet. Gynecol. 2007, 196, 382.e1–382.e8. [Google Scholar] [CrossRef]
- Steinborn, A.; Rebmann, V.; Scharf, A.; Sohn, C.; Grosse-Wilde, H. Placental Abruption Is Associated with Decreased Maternal Plasma Levels of Soluble HLA-G. J. Clin. Immunol. 2003, 23, 307–314. [Google Scholar] [CrossRef]
- Steinborn, A.; Seidl, C.; Sayehli, C.; Sohn, C.; Seifried, E.; Kaufmann, M.; Schmitt, E. Anti-fetal immune response mechanisms may be involved in the pathogenesis of placental abruption. Clin. Immunol. 2004, 110, 45–54. [Google Scholar] [CrossRef]
- Szekeres-Bartho, J.; Varga, P.; Pacsa, A. Immunologic factors contributing to the initiation of labor—Lymphocyte reactivity in term labor and threatened preterm delivery. Am. J. Obstet. Gynecol. 1986, 155, 108–112. [Google Scholar] [CrossRef]
- Abadia-Molina, A.C.; Ruiz, C.; Montes, M.; King, A.; Loke, Y.; Olivares, E.G. Immune phenotype and cytotoxic activity of lymphocytes from human term decidua against trophoblast. J. Reprod. Immunol. 1996, 31, 109–123. [Google Scholar] [CrossRef]
- Osman, I.; Young, A.; Ledingham, M.A.; Thomson, A.J.; Jordan, F.; Greer, I.A.; Norman, J.E. Leukocyte density and pro-inflammatory cytokine expression in human fetal membranes, decidua, cervix and myometrium before and during labour at term. Mol. Hum. Reprod. 2003, 9, 41–45. [Google Scholar] [CrossRef]
- Steinborn, A.; Sohn, C.; Sayehli, C.; Baudendistel, A.; Hüwelmeier, D.; Solbach, C.; Schmitt, E.; Kaufmann, M. Spontaneous labour at term is associated with fetal monocyte activation. Clin. Exp. Immunol. 1999, 117, 147–152. [Google Scholar] [CrossRef]
- Sindram-Trujillo, A.P.; Scherjon, S.A.; van Hulst-van Miert, P.P.; Kanhai, H.H.; Roelen, D.L.; Claas, F.H. Comparison of decidual leukocytes following spontaneous vaginal delivery and elective cesarean section in uncomplicated human term pregnancy. J. Reprod. Immunol. 2004, 62, 125–137. [Google Scholar] [CrossRef]
- Lin, H.; Mosmann, T.R.; Guilbert, L.; Tuntipopipat, S.; Wegmann, T.G. Synthesis of T helper 2-type cytokines at the maternal-fetal interface. J. Immunol. 1993, 151, 4562–4573. [Google Scholar]
- Steinborn, A.; Rebmann, V.; Scharf, A.; Sohn, C.; Grosse-Wilde, H. Soluble HLA-DR levels in the maternal circulation of normal and pathologic pregnancy. Am. J. Obstet. Gynecol. 2003, 188, 473–479. [Google Scholar] [CrossRef]
- Kryczek, I.; Wei, S.; Zou, L.; Zhu, G.; Mottram, P.; Xu, H.; Chen, L.; Zou, W. Cutting Edge: Induction of B7-H4 on APCs through IL-10: Novel Suppressive Mode for Regulatory T Cells. J. Immunol. 2006, 177, 40–44. [Google Scholar] [CrossRef]
- Kryczek, I.; Wei, S.; Zhu, G.; Myers, L.; Mottram, P.; Cheng, P.; Chen, L.; Coukos, G.; Zou, W. Relationship between B7-H4, Regulatory T Cells, and Patient Outcome in Human Ovarian Carcinoma. Cancer Res. 2007, 67, 8900–8905. [Google Scholar] [CrossRef] [PubMed]
- Kryczek, I.; Zou, L.; Rodriguez, P.; Zhu, G.; Wei, S.; Mottram, P.; Brumlik, M.; Cheng, P.; Curiel, T.; Myers, L.; et al. B7-H4 expression identifies a novel suppressive macrophage population in human ovarian carcinoma. J. Exp. Med. 2006, 203, 871–881. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Hao, J.; Metzger, D.; Ao, Z.; Chen, L.; Ou, D.; Verchere, B.; Mui, A.; Warnock, G.L. B7-H4 Treatment of T Cells Inhibits ERK, JNK, p38, and AKT Activation. PLoS ONE 2012, 7, e28232. [Google Scholar] [CrossRef] [PubMed]
- Collins, M.; Ling, V.; Carreno, B.M. The B7 family of immune-regulatory ligands. Genome Biol. 2005, 6, 223. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lafferty, K.J.; Cunningham, A.J. A NEW ANALYSIS OF ALLOGENEIC INTERACTIONS. Aust. J. Exp. Biol. Med Sci. 1975, 53, 27–42. [Google Scholar] [CrossRef]
- Jenkins, M.K.; Schwartz, R.H. Antigen presentation by chemically modified splenocytes induces antigen-specific T cell unre-sponsiveness in vitro and in vivo. J. Exp. Med. 1987, 165, 302–319. [Google Scholar] [CrossRef]
- Holt, M.P.; Punkosdy, G.A.; Glass, D.D.; Shevach, E.M. TCR Signaling and CD28/CTLA-4 Signaling Cooperatively Modulate T Regulatory Cell Homeostasis. J. Immunol. 2017, 198, 1503–1511. [Google Scholar] [CrossRef]
- Wicherek, L.; Galazka, K. The possible correlation between the patient’s immune tolerance level during cesaerean section and the incidence of subsequent emergency peripartum hysterectomy. Clin. Dev. Immunol. 2007, 2007, 63596. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patients (n = 148) | Maternal Age ± SD (Year) | BMI ± SD (BMI) | Gestational Age ± SD (Week) | Parity Nulliparous (%) | Newborn Weight ± SD (g) | Newborn Length ± SD (cm) | Live Births (%) |
|---|---|---|---|---|---|---|---|
| Placental abruption (n = 47) | 33 ± 5 | 26 ± 4 | 30 ± 4 | 44 | 1489 ± 865 | 40 ± 7 | 87 |
| Retained placental tissue (n = 60) | 31 ± 5 | 26 ± 4 | 32 ± 6 | 40 | 1959 ± 1128 | 44 ± 9 | 67 |
| Healthy controls (n = 41) | 32 ± 6 | 27 ± 6 | 37 ± 3 | 37 | 2851 ± 738 | 51 ± 4 | 95 |
| p-value | 0.368 | 0.770 | <0.01 | 0.739 | <0.01 | <0.01 | <0.01 |
| Percentage of Cells Showing Reactivity (%) | Staining Pattern | Stage |
|---|---|---|
| <1 | No reactivity | 0 |
| 1–20 | Any | 1 |
| 21–50 | Low | 2 |
| >50 | Low | 3 |
| >20 | High | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bączkowska, M.; Dutsch-Wicherek, M.M.; Przytuła, E.; Faryna, J.; Wojtyła, C.; Ali, M.; Knafel, A.; Ciebiera, M. Expression of the Costimulatory Molecule B7-H4 in the Decidua and Placental Tissues in Patients with Placental Abruption. Biomedicines 2022, 10, 918. https://doi.org/10.3390/biomedicines10040918
Bączkowska M, Dutsch-Wicherek MM, Przytuła E, Faryna J, Wojtyła C, Ali M, Knafel A, Ciebiera M. Expression of the Costimulatory Molecule B7-H4 in the Decidua and Placental Tissues in Patients with Placental Abruption. Biomedicines. 2022; 10(4):918. https://doi.org/10.3390/biomedicines10040918
Chicago/Turabian StyleBączkowska, Monika, Magdalena Maria Dutsch-Wicherek, Ewa Przytuła, Jan Faryna, Cezary Wojtyła, Mohamed Ali, Anna Knafel, and Michał Ciebiera. 2022. "Expression of the Costimulatory Molecule B7-H4 in the Decidua and Placental Tissues in Patients with Placental Abruption" Biomedicines 10, no. 4: 918. https://doi.org/10.3390/biomedicines10040918
APA StyleBączkowska, M., Dutsch-Wicherek, M. M., Przytuła, E., Faryna, J., Wojtyła, C., Ali, M., Knafel, A., & Ciebiera, M. (2022). Expression of the Costimulatory Molecule B7-H4 in the Decidua and Placental Tissues in Patients with Placental Abruption. Biomedicines, 10(4), 918. https://doi.org/10.3390/biomedicines10040918