
COVID-19 Infection Induce miR-371a-3p Upregulation Resulting in Influence on Male Fertility

 , , , , , , ,
, , , , , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Testicular Histological Changes
2.2. FFPE Tissue Samples
2.3. Serum Samples
2.4. Light Microscopy and Immunohistochemistry of FFPE Tissues
2.5. Double Labelling of AR Immunohistochemistry of FFPE Tissues and by miR-371a-3p In Situ PCR
2.6. Immunofluorescence of ACE2 Protein
2.7. Protein Extraction from FFPE
2.8. ELISA
2.9. miR Extraction from FFPE Tissue and RT-PCR
2.10. Quantitative Real-Time PCR (qRT-PCR)
2.11. Statistical Analysis
3. Results
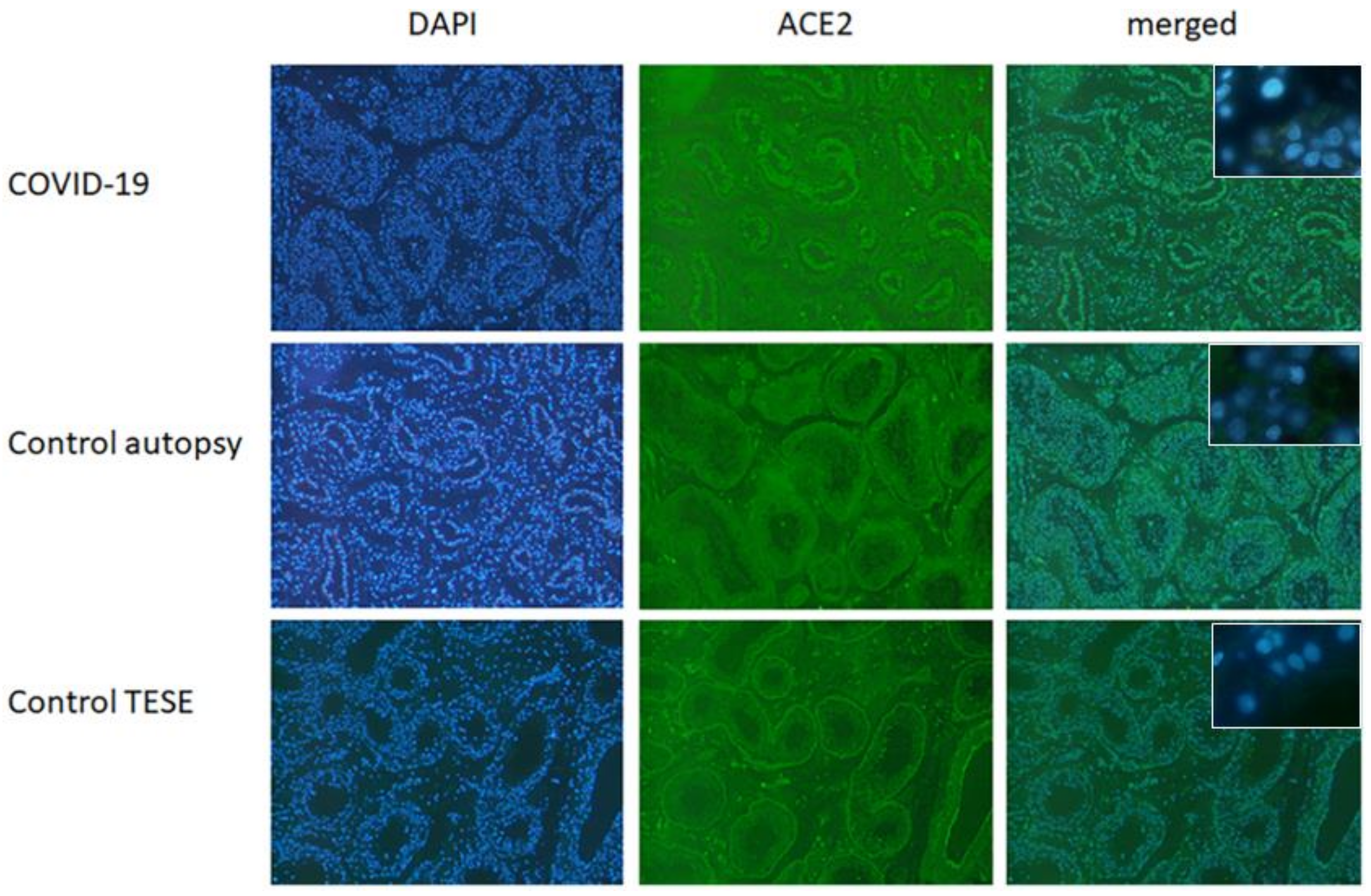
3.1. Immunofluorescence ACE2 in Testicular Tissue from Healthy Donors and COVID-19 Patients
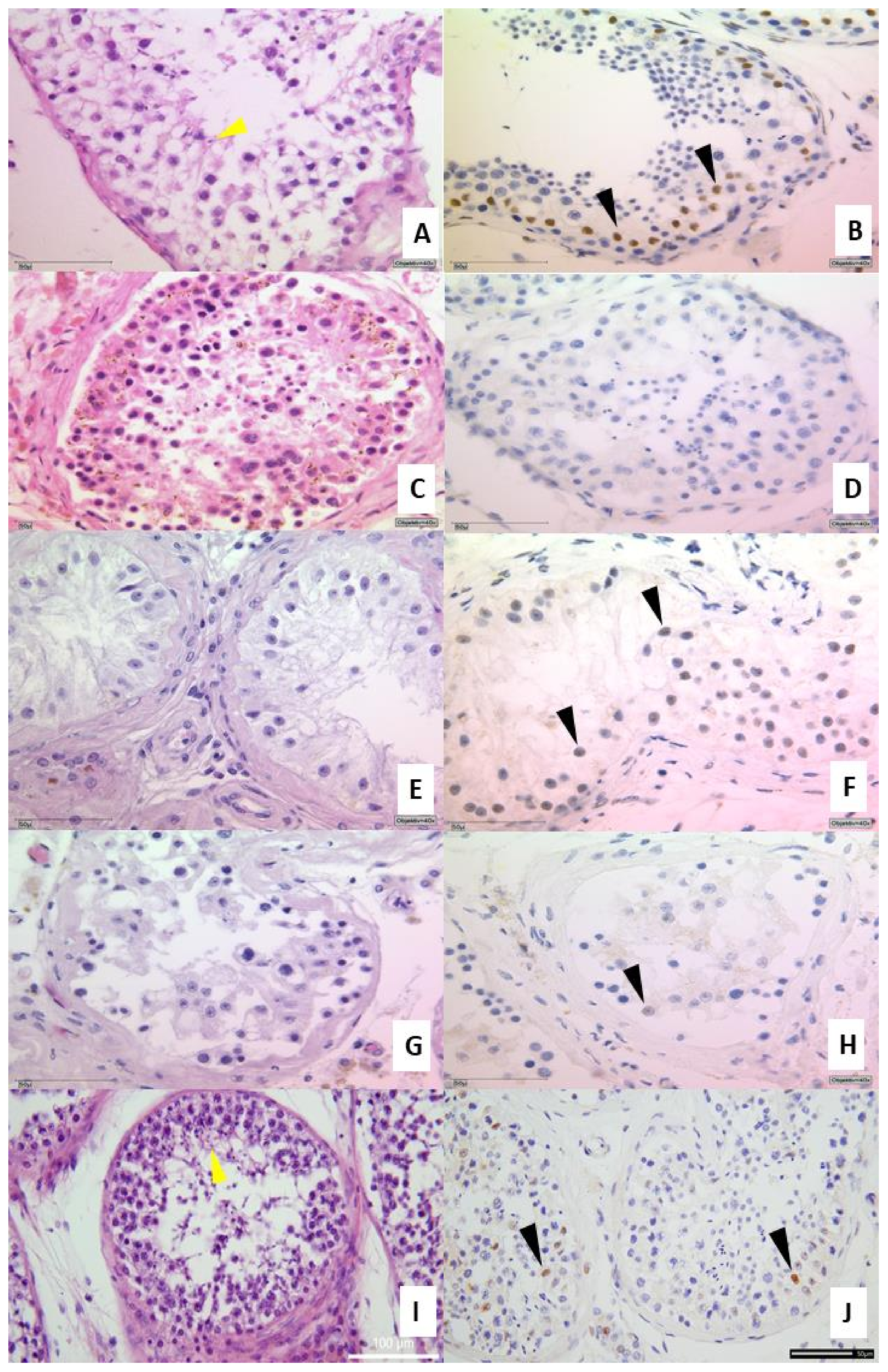
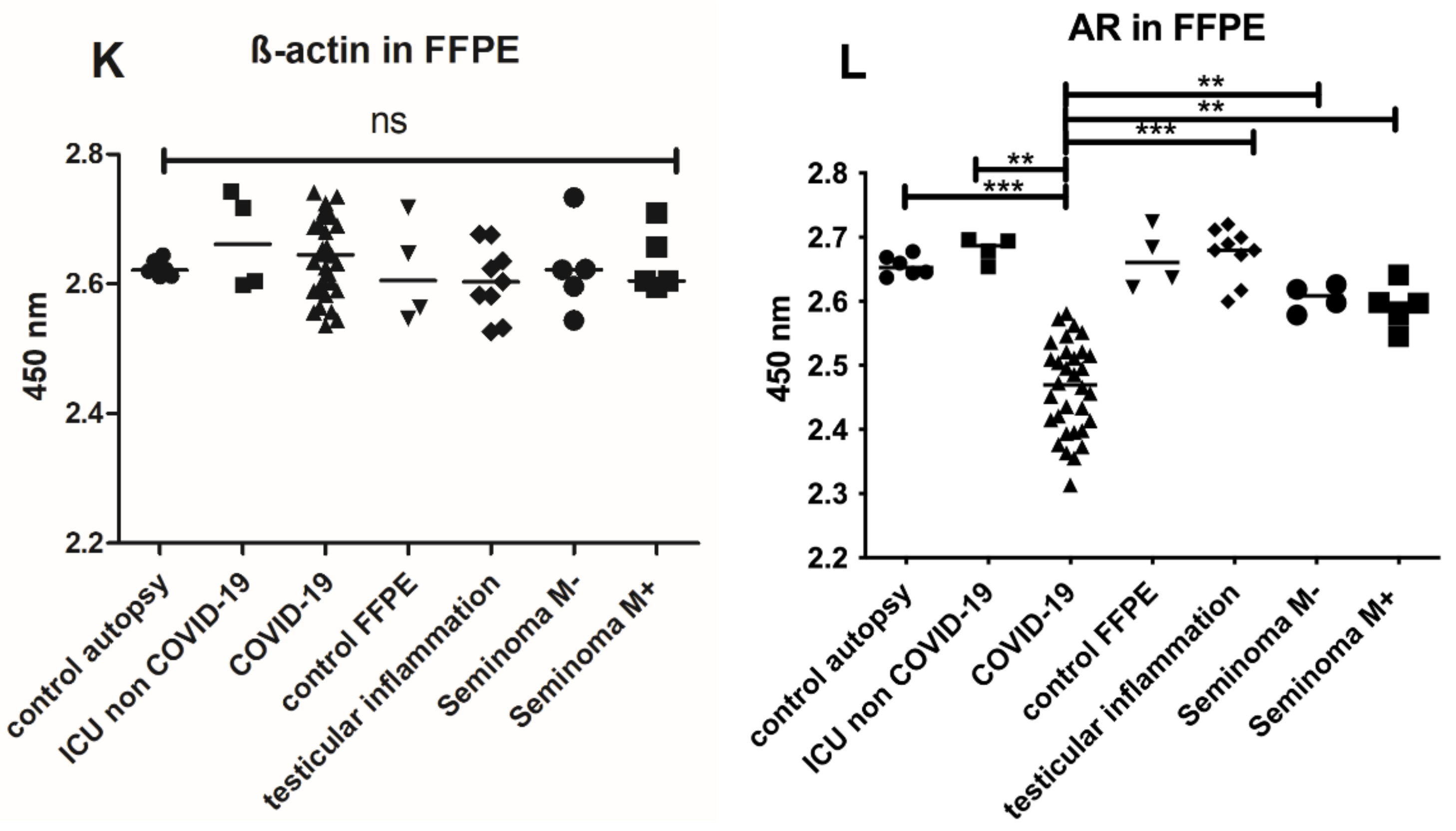
3.2. Histology and Expression of the Androgen Receptor in Testicular Tissue
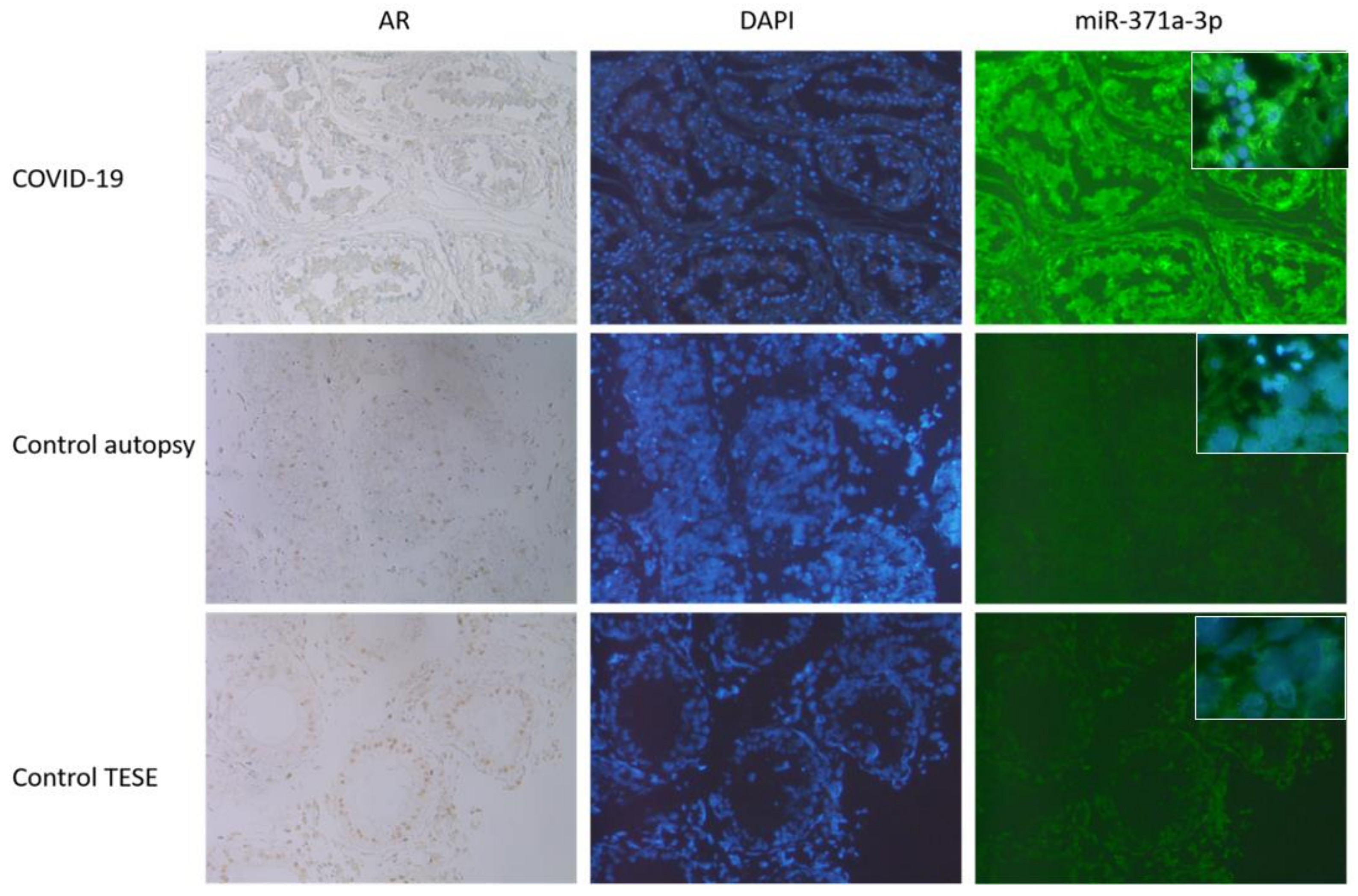
3.3. AR IHC Followed by miR-371a-3p In Situ PCR
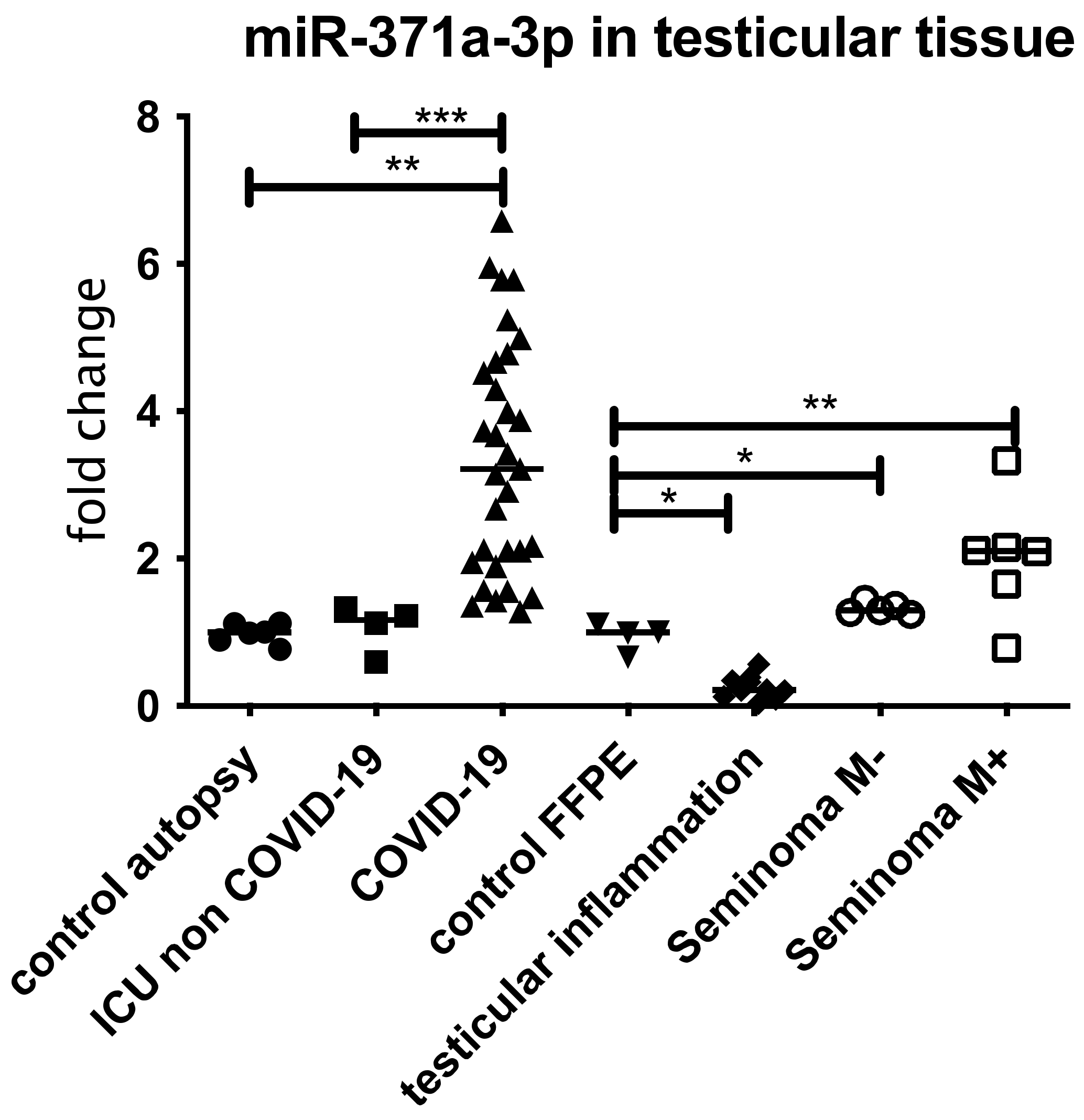
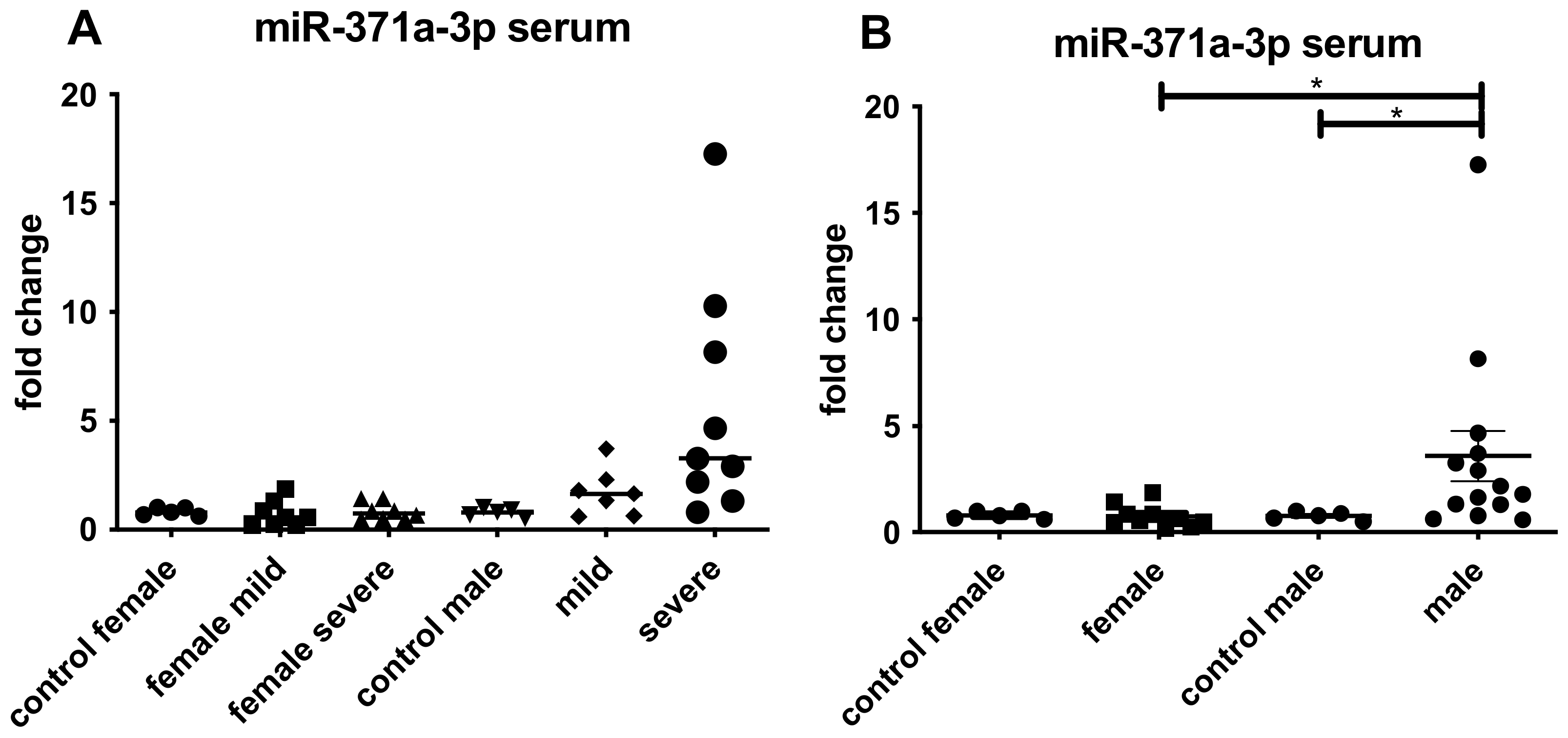
3.4. Expression of miR 371a-3p in Testicular Tissue Samples and Serum from COVID-19 Patients
4. Discussion
4.1. COVID-19 and Infiltration of Testes
4.2. COVID-19 and Androgen Receptor (AR)
4.3. COVID-19 and miR-371a-3p Overexpression
4.4. AR Expression in COVID-19 Patients
4.5. Detection of miR-371a-3p in Blood Samples
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, Y.; Geng, X.; Tan, Y.; Li, Q.; Xu, C.; Xu, J.; Hao, L.; Zeng, Z.; Luo, X.; Liu, F.; et al. New understanding of the damage of SARS-CoV-2 infection outside the respiratory system. Biomed. Pharmacother. 2020, 127, 110195. [Google Scholar] [CrossRef] [PubMed]
- Kirtipal, N.; Kumar, S.; Dubey, S.K.; Dwivedi, V.D.; Gireesh Babu, K.; Maly, P.; Bharadwaj, S. Understanding on the possible routes for SARS CoV-2 invasion via ACE2 in the host linked with multiple organs damage. Infect. Genet. Evol. 2022, 99, 105254. [Google Scholar] [CrossRef] [PubMed]
- Salamanna, F.; Maglio, M.; Landini, M.P.; Fini, M. Body Localization of ACE-2: On the Trail of the Keyhole of SARS-CoV-2. Front. Med. 2020, 7, 594495. [Google Scholar] [CrossRef] [PubMed]
- Ejaz, H.; Alsrhani, A.; Zafar, A.; Javed, H.; Junaid, K.; Abdalla, A.E.; Abosalif, K.O.A.; Ahmed, Z.; Younas, S. COVID-19 and comorbidities: Deleterious impact on infected patients. J. Infect. Public Health 2020, 13, 1833–1839. [Google Scholar] [CrossRef] [PubMed]
- Migliore, L.; Nicoli, V.; Stoccoro, A. Gender Specific Differences in Disease Susceptibility: The Role of Epigenetics. Biomedicines 2021, 9, 652. [Google Scholar] [CrossRef] [PubMed]
- Cosar, B.; Karagulleoglu, Z.Y.; Unal, S.; Ince, A.T.; Uncuoglu, D.B.; Tuncer, G.; Kilinc, B.R.; Ozkan, Y.E.; Ozkoc, H.C.; Demir, I.N.; et al. SARS-CoV-2 Mutations and their Viral Variants. Cytokine Growth Factor Rev. 2021, 63, 10–22. [Google Scholar] [CrossRef]
- Barbagallo, F.; Calogero, A.E.; Cannarella, R.; Condorelli, R.A.; Mongioi, L.M.; Aversa, A.; La Vignera, S. The testis in patients with COVID-19: Virus reservoir or immunization resource? Transl. Androl. Urol. 2020, 9, 1897–1900. [Google Scholar] [CrossRef] [PubMed]
- Markiewicz-Gospodarek, A.; Wdowiak, P.; Czeczelewski, M.; Forma, A.; Flieger, J.; Januszewski, J.; Radzikowska-Buchner, E.; Baj, J. The Impact of SARS-CoV-2 Infection on Fertility and Female and Male Reproductive Systems. J. Clin. Med. 2021, 10, 4520. [Google Scholar] [CrossRef] [PubMed]
- Heidenreich, A.; Bokemeyer, C.; Souchon, R. Stage-specific treatment for testicular germ cell tumours. Urol. A 2009, 48, 377–385. [Google Scholar] [CrossRef]
- Clark, A.T. The stem cell identity of testicular cancer. Stem Cell Rev. 2007, 3, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Giannandrea, F.; Fargnoli, S. Environmental Factors Affecting Growth and Occurrence of Testicular Cancer in Childhood: An Overview of the Current Epidemiological Evidence. Children 2017, 4, 1. [Google Scholar] [CrossRef] [PubMed]
- Richiardi, L.; Pettersson, A.; Akre, O. Genetic and environmental risk factors for testicular cancer. Int. J. Androl. 2007, 30, 230–240. [Google Scholar] [CrossRef]
- Dieckmann, K.P.; Radtke, A.; Geczi, L.; Matthies, C.; Anheuser, P.; Eckardt, U.; Sommer, J.; Zengerling, F.; Trenti, E.; Pichler, R.; et al. Serum Levels of MicroRNA-371a-3p (M371 Test) as a New Biomarker of Testicular Germ Cell Tumors: Results of a Prospective Multicentric Study. J. Clin. Oncol. 2019, 37, 1412–1423. [Google Scholar] [CrossRef]
- Pan, B.; He, B.; Xu, X.; Liu, X.; Xu, T.; Xu, M.; Chen, X.; Zeng, K.; Lin, K.; Hu, X.; et al. MicroRNA-371-3 cluster as biomarkers for the diagnosis and prognosis of cancers. Cancer Manag. Res. 2019, 11, 5437–5457. [Google Scholar] [CrossRef] [PubMed]
- Skalsky, R.L.; Cullen, B.R. Viruses, microRNAs, and host interactions. Annu. Rev. Microbiol. 2010, 64, 123–141. [Google Scholar] [CrossRef]
- Von Brandenstein, M.; Richter, C.; Fries, J.W. MicroRNAs: Small but amazing, and their association with endothelin. Life Sci. 2012, 91, 475–489. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, C.E.; Sulpice, E.; Combe, S.; Shibakawa, A.; Leach, D.A.; Hamilton, M.P.; Chrysostomou, S.L.; Sharp, A.; Welti, J.; Yuan, W.; et al. Androgen receptor-modulatory microRNAs provide insight into therapy resistance and therapeutic targets in advanced prostate cancer. Oncogene 2019, 38, 5700–5724. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, H.; Ueda, T.; Ito, S.; Shiraishi, T.; Taniguchi, H.; Kayukawa, N.; Nakanishi, H.; Ushijima, S.; Kanazawa, M.; Nakamura, T.; et al. Androgen suppresses testicular cancer cell growth in vitro and in vivo. Oncotarget 2016, 7, 35224–35232. [Google Scholar] [CrossRef] [PubMed]
- Sigg, C. Classification of tubular testicular atrophies in the diagnosis of sterility. Significance of the so-called “bunte Atrophie”. Schweiz. Med. Wochenschr. 1979, 109, 1284–1293. [Google Scholar] [PubMed]
- Sigg, C.; Hedinger, C. Testicular germ cell tumors and atypical germ cells. Schweiz. Med. Wochenschr. 1980, 110, 801–806. [Google Scholar] [PubMed]
- Nuovo, G.J.; Elton, T.S.; Nana-Sinkam, P.; Volinia, S.; Croce, C.M.; Schmittgen, T.D. A methodology for the combined in situ analyses of the precursor and mature forms of microRNAs and correlation with their putative targets. Nat. Protoc. 2009, 4, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, K.; Monden, T.; Kanoh, T.; Tsujie, M.; Izawa, H.; Haba, A.; Ohnishi, T.; Sekimoto, M.; Tomita, N.; Shiozaki, H.; et al. Extraction and analysis of diagnostically useful proteins from formalin-fixed, paraffin-embedded tissue sections. J. Histochem. Cytochem. 1998, 46, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Gerstung, M.; Roth, T.; Dienes, H.P.; Licht, C.; Fries, J.W. Endothelin-1 induces NF-kappaB via two independent pathways in human renal tubular epithelial cells. Am. J. Nephrol. 2007, 27, 294–300. [Google Scholar] [CrossRef] [PubMed]
- Von Brandenstein, M.; Puetz, K.; Schlosser, M.; Loser, H.; Kallinowski, J.P.; Godde, D.; Buettner, R.; Storkel, S.; Fries, J.W. Vimentin 3, the new hope, differentiating RCC versus oncocytoma. Dis. Markers 2015, 2015, 368534. [Google Scholar] [CrossRef]
- Von Brandenstein, M.; Depping, R.; Schafer, E.; Dienes, H.P.; Fries, J.W. Protein kinase C alpha regulates nuclear pri-microRNA 15a release as part of endothelin signaling. Biochim. Biophys. Acta 2011, 1813, 1793–1802. [Google Scholar] [CrossRef] [PubMed]
- Samuel, R.M.; Majd, H.; Richter, M.N.; Ghazizadeh, Z.; Zekavat, S.M.; Navickas, A.; Ramirez, J.T.; Asgharian, H.; Simoneau, C.R.; Bonser, L.R.; et al. Androgen Signaling Regulates SARS-CoV-2 Receptor Levels and Is Associated with Severe COVID-19 Symptoms in Men. Cell Stem Cell 2020, 27, 876–889.e12. [Google Scholar] [CrossRef] [PubMed]
- Dutta, S.; Sengupta, P. SARS-CoV-2 and Male Infertility: Possible Multifaceted Pathology. Reprod. Sci. 2021, 28, 23–26. [Google Scholar] [CrossRef]
- Wang, N.; Qin, L.; Ma, L.; Yan, H. Effect of severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2) on reproductive system. Stem Cell Res. 2021, 52, 102189. [Google Scholar] [CrossRef]
- Medina, F.A.; Torres, G.; Acevedo, J.; Fonseca, S.; Casiano, L.; De Leon-Rodriguez, C.M.; Santiago, G.A.; Doyle, K.; Sharp, T.M.; Alvarado, L.I.; et al. Duration of the Presence of Infectious Zika Virus in Semen and Serum. J. Infect. Dis. 2019, 219, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Illiano, E.; Trama, F.; Costantini, E. Could COVID-19 have an impact on male fertility? Andrologia 2020, 52, e13654. [Google Scholar] [CrossRef]
- Almeida, S.; Rato, L.; Sousa, M.; Alves, M.G.; Oliveira, P.F. Fertility and Sperm Quality in the Aging Male. Curr. Pharm. Des. 2017, 23, 4429–4437. [Google Scholar] [CrossRef] [PubMed]
- Bhowmick, N.A.; Oft, J.; Dorff, T.; Pal, S.; Agarwal, N.; Figlin, R.A.; Posadas, E.M.; Freedland, S.J.; Gong, J. COVID-19 and androgen-targeted therapy for prostate cancer patients. Endocr. Relat. Cancer 2020, 27, R281–R292. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, M.S.; Moulin, T.C.; Schioth, H.B. Sex differences in COVID-19: The role of androgens in disease severity and progression. Endocrine 2021, 71, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Kuba, K.; Imai, Y.; Rao, S.; Gao, H.; Guo, F.; Guan, B.; Huan, Y.; Yang, P.; Zhang, Y.; Deng, W.; et al. A crucial role of angiotensin converting enzyme 2 (ACE2) in SARS coronavirus-induced lung injury. Nat. Med. 2005, 11, 875–879. [Google Scholar] [CrossRef] [PubMed]
- Stilhano, R.S.; Costa, A.J.; Nishino, M.S.; Shams, S.; Bartolomeo, C.S.; Breithaupt-Faloppa, A.C.; Silva, E.A.; Ramirez, A.L.; Prado, C.M.; Ureshino, R.P. SARS-CoV-2 and the possible connection to ERs, ACE2, and RAGE: Focus on susceptibility factors. FASEB J. 2020, 34, 14103–14119. [Google Scholar] [CrossRef] [PubMed]
- Montopoli, M.; Zumerle, S.; Vettor, R.; Rugge, M.; Zorzi, M.; Catapano, C.V.; Carbone, G.M.; Cavalli, A.; Pagano, F.; Ragazzi, E.; et al. Androgen-deprivation therapies for prostate cancer and risk of infection by SARS-CoV-2: A population-based study (N = 4532). Ann. Oncol. 2020, 31, 1040–1045. [Google Scholar] [CrossRef] [PubMed]
- Garolla, A.; Vitagliano, A.; Muscianisi, F.; Valente, U.; Ghezzi, M.; Andrisani, A.; Ambrosini, G.; Foresta, C. Role of Viral Infections in Testicular Cancer Etiology: Evidence from a Systematic Review and Meta-Analysis. Front. Endocrinol. 2019, 10, 355. [Google Scholar] [CrossRef]
- Li, H.; Xiao, X.; Zhang, J.; Zafar, M.I.; Wu, C.; Long, Y.; Lu, W.; Pan, F.; Meng, T.; Zhao, K.; et al. Impaired spermatogenesis in COVID-19 patients. EClinicalMedicine 2020, 28, 100604. [Google Scholar] [CrossRef]
- Mannur, S.; Jabeen, T.; Khader, M.A.; Rao, L.S.S. Post-Covid-19 Associated Decline in Long-Term Male Fertility and Embryo Quality during Assisted Reproductive Technology. QJM 2021, 114, 328–330. [Google Scholar] [CrossRef]
- Kaufman, J.M.; Vermeulen, A. The decline of androgen levels in elderly men and its clinical and therapeutic implications. Endocr. Rev. 2005, 26, 833–876. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.S.; Yeh, S.; Tzeng, C.R.; Chang, C. Androgen receptor roles in spermatogenesis and fertility: Lessons from testicular cell-specific androgen receptor knockout mice. Endocr. Rev. 2009, 30, 119–132. [Google Scholar] [CrossRef] [PubMed]
- Sigg, C.; Hedinger, C. Atypical germ cells in testicular biopsy in male sterility. Int. J. Androl. 1981, 4 (Suppl. S4), 163–170. [Google Scholar] [CrossRef] [PubMed]
- McCoy, J.; Wambier, C.G.; Vano-Galvan, S.; Shapiro, J.; Sinclair, R.; Ramos, P.M.; Washenik, K.; Andrade, M.; Herrera, S.; Goren, A. Racial variations in COVID-19 deaths may be due to androgen receptor genetic variants associated with prostate cancer and androgenetic alopecia. Are anti-androgens a potential treatment for COVID-19? J. Cosmet. Dermatol. 2020, 19, 1542–1543. [Google Scholar] [CrossRef] [PubMed]
- Leite, K.R.; Morais, D.R.; Florez, M.G.; Reis, S.T.; Iscaife, A.; Viana, N.; Moura, C.M.; Silva, I.A.; Katz, B.S.; Pontes, J., Jr.; et al. The role of microRNAs 371 and 34a in androgen receptor control influencing prostate cancer behavior. Urol. Oncol. 2015, 33, 267.e15–267.e22. [Google Scholar] [CrossRef]
- Radtke, A.; Dieckmann, K.P.; Grobelny, F.; Salzbrunn, A.; Oing, C.; Schulze, W.; Belge, G. Expression of miRNA-371a-3p in seminal plasma and ejaculate is associated with sperm concentration. Andrology 2019, 7, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Elzinga-Tinke, J.E.; Dohle, G.R.; Looijenga, L.H. Etiology and early pathogenesis of malignant testicular germ cell tumors: Towards possibilities for preinvasive diagnosis. Asian J. Androl. 2015, 17, 381–393. [Google Scholar] [CrossRef] [PubMed]
- Van der Zwan, Y.G.; Rijlaarsdam, M.A.; Rossello, F.J.; Notini, A.J.; de Boer, S.; Watkins, D.N.; Gillis, A.J.; Dorssers, L.C.; White, S.J.; Looijenga, L.H. Seminoma and embryonal carcinoma footprints identified by analysis of integrated genome-wide epigenetic and expression profiles of germ cell cancer cell lines. PLoS ONE 2014, 9, e98330. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Zhu, S.; Dai, Z.; Hao, L.; Luan, C.; Guo, Q.; Meng, C.; Zhang, Y. Effects of COVID-19 and mRNA vaccines on human fertility. Hum. Reprod. 2021, 37, 5–13. [Google Scholar] [CrossRef]
- Eringyte, I.; Zamarbide Losada, J.N.; Powell, S.M.; Bevan, C.L.; Fletcher, C.E. Coordinated AR and microRNA regulation in prostate cancer. Asian J. Urol. 2020, 7, 233–250. [Google Scholar] [CrossRef]
- Tanaka, M.; Ueno, T.; Nakahara, T.; Sasaki, K.; Ishimoto, A.; Sakai, H. Downregulation of CD4 is required for maintenance of viral infectivity of HIV-1. Virology 2003, 311, 316–325. [Google Scholar] [CrossRef]
- Davison, S.L.; Bell, R.; Donath, S.; Montalto, J.G.; Davis, S.R. Androgen levels in adult females: Changes with age, menopause, and oophorectomy. J. Clin. Endocrinol. Metab. 2005, 90, 3847–3853. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Consecutively Autopsied COVID-19 Positive Male Patients: Spermiogenesis and miR-371a-3p | Control: Age Adjusted Autopsied Male Patients |
|---|---|
| Control: testes biopsied (TESE) due to inflammation for exclusion of inflammatory process involvement | |
| Living COVID-19 male patients: serum samples for miR-371a-3p | Control: Intensive Care/ECMO adjusted autopsied male patients for exclusion of therapy process involvement |
| Control: seminoma for miR-371a-3p expression levels |
| Age | BMI | AR IHC | Spermiogenesis | Sertoli Cells | Interstitial Area | Inflammation | Tumor/ GCNIS | Atrophy Class Acc. SIGG | |
|---|---|---|---|---|---|---|---|---|---|
| COVID-19 | |||||||||
| 85 | n.d. | neg | one testis: some spermatogonia and primary spermatocytes | autolytic changes | partial atrophy in one testes, the other complete atrophy | neg | neg | IIB | |
| 70 | n.d. | neg | normal spermiogenesis in both testes | autolytic changes | discrete interstitial edema | neg | neg | nl | |
| 76 | n.d. | neg | some spermatogonia and primary spermatocytes in both testes | autolytic changes | focal fibrosis, focal slightly thickened basement membrane of tubules | neg | neg | IIB | |
| 80 | 37.1 | neg | typical spermiogenesis in one testis, the other spermatogonia and primary spermatocytes | autolytic changes | interstitial edema | neg | neg | nl/IIB | |
| 56 | 29 | neg | single spermatogonia and primary spermatocytes | autolytic changes | slightly thickened basement membrane of tubules, discrete edema, focal interstitial fibrosis | neg | neg | IIB | |
| 64 | 26.9 | neg | single spermatogonia | autolytic changes | interstitial edema, focal interstitial fibrosis | neg | neg | IIC | |
| 74 | 42.8 | neg | single spermatogonia and primary spermatocytes | autolytic changes | medium interstitial fibrosis | neg | neg | IIB | |
| 72 | 33.9 | neg, IC + | reduced spermiogenesis | autolytic changes | discrete interstitial edema | neg | neg | I | |
| 85 | n.d. | neg | one testis: some spermatogonia and primary spermatocytes | autolytic changes | partial atrophy in one testes, the other complete atrophy | neg | neg | IIB | |
| 70 | n.d. | neg | normal spermiogenesis in both testes | autolytic changes | discrete interstitial edema | neg | neg | nl | |
| 76 | n.d. | neg | some spermatogonia and primary spermatocytes in both testes | autolytic changes | focal fibrosis, focal slightly thickened basement membrane of tubules | neg | neg | IIB | |
| 80 | 37.1 | neg | typical spermiogenesis in one testis, the other spermatogonia and primary spermatocytes | autolytic changes | interstitial edema | neg | neg | nl/IIB | |
| 56 | 29 | neg | single spermatogonia and primary spermatocytes | autolytic changes | slightly thickened basement membrane of tubules, discrete edema, focal interstitial fibrosis | neg | neg | IIB | |
| 64 | 26.9 | neg | single spermatogonia | autolytic changes | interstitial edema, focal interstitial fibrosis | neg | neg | IIC | |
| 74 | 42.8 | neg | single spermatogonia and primary spermatocytes | autolytic changes | medium interstitial fibrosis | neg | neg | IIB | |
| 72 | 33.9 | neg, IC+ | reduced spermiogenesis | autolytic changes | discrete interstitial edema | neg | neg | I | |
| 65 | 25.86 | (+), IC + | normal spermiogenesis | autolytic changes | discrete interstitial edema | neg | neg | 0 | |
| 72 | 31.14 | neg | slight reduced spermiogenesis | autolytic changes | discrete interstitial edema | neg | neg | I | |
| 47 | 20.76 | pos, IC + | normal spermiogenesis | autolytic changes | discrete interstitial edema | neg | neg | 0 | |
| 49 | 27 | neg, IC (+) | sertoli only | autolytic changes | fibrosis | neg | neg | IV | |
| 69 | 28.9 | neg | reduced spermiogenesis | autolytic changes | discrete interstitial edema | neg | neg | IIa | |
| 72 | 22.09 | neg | sertoli only | autolytic changes | fibrosis | neg | neg | IV | |
| 58 | 26.3 | (+), IC neg | slight reduced spermiogenesis | autolytic changes | discrete interstitial edema | neg | neg | I | |
| 76 | 31.6 | neg, IC (+) | reduced spermiogenesis | autolytic changes | discrete interstitial edema | neg | neg | I-IIa | |
| 83 | 27 | (+), IC (+) | slight reduced spermiogenesis | autolytic changes | discrete interstitial edema | neg | neg | I | |
| 58 | 24.7 | neg | sertoli only | autolytic changes | interstitial edema, interstitial fibrosis | neg | neg | IV | |
| 66 | 28.7 | neg | sertoli only | autolytic changes | interstitial edema | neg | neg | IV | |
| 55 | 23 | neg, IC + | reduced spermiogenesis | autolytic changes | interstitial edema | neg | neg | III | |
| 60 | 36.08 | neg | sertoli only | autolytic changes | fibrosis | neg | neg | IV | |
| 66 | 30.68 | neg, IC (+) | reduced spermiogenesis | autolytic changes | edema | neg | neg | IIB | |
| 73 | 23.67 | neg, IC (+) | slight reduced spermiogenesis | autolytic changes | discrete interstitial edema | neg | neg | I | |
| (+), IC (+) | reduced spermiogenesis | autolytic changes | discrete interstitial edema, fibrosis | neg | neg | IIA/IIB | |||
| Mean | 64 | 27 | |||||||
| CONTROLS INFLAMMATION | |||||||||
| 57 | n.d. | neg | extensive necrosis of tubular structures | neg | extensive necrosis of tubular structures | massive | neg | n.d. | |
| 24 | n.d. | + | spermatogonia and primary spermatocytes | pos | slightly thickened basement membrane, discrete edema | discrete | neg | IIB | |
| 78 | n.d. | focal (+) | partial tubules with Sertoli cells | pos | partial complete atrophy, focal slightly thickened basement membrane of tubules, focal interstitial fibrosis | massive | neg | IV | |
| 85 | n.d. | ++ | neg | autolytic changes | focal slightly thickened basement membrane of tubules, focal interstitial fibrosis | discrete | neg | IV–V | |
| 75 | n.d. | +, int con ++ | neg | autolytic changes | focal slightly thickened basement membrane, focal interstitial fibrosis | focal | neg | V | |
| 79 | n.d. | ++ | some spermatogonia and primary spermatocytes and spermatids | pos | focal interstitial fibrosis and edema | discrete | neg | IIA | |
| 79 | n.d. | + | some spermatogonia and primary spermatocytes and spermatids | autolytic changes | focal slightly thickened basement membrane of tubules, focal interstitial fibrosis | discrete | neg | IIA | |
| 81 | n.d. | + | single spermatogonia and primary spermatocytes, no further spermiogenesis | autolytic changes | medium interstitial fibrosis | neg | neg | IIA | |
| 44 | n.d. | neg | neg | autolytic changes | focal interstitial fibrosis | massive | neg | n.d | |
| Mean | 66.8 | ||||||||
| AGE/AUTOPSY CONTROLS | |||||||||
| 61 | n.d. | + | some spermatogonia and primary spermatocytes and spermatids | pos | focal slightly thickened basement membrane, focal interstitial fibrosis | neg | neg | IIA | |
| 53 | n.d. | + | reduced spermiogenesis | discrete interstitial edema | neg | neg | I | ||
| 63 | n.d. | + | normal spermiogenesis | discrete interstitial edema | neg | neg | Nl | ||
| 76 | 23,2 | neg | reduced spermiogenesis | discrete interstitial edema | neg | neg | I | ||
| 65 | n.d. | neg | some spermatogonia and primary spermatocytes and spermatids | autolytic changes | focal slightly thickened basement membrane, focal interstitial fibrosi | neg | neg | IIA | |
| 59 | ++ | normal spermiogenesis | discrete interstitial edema | neg | neg | Nl | |||
| Mean | 66.7 | ||||||||
| INTENSIVE CARE CONTROLS | |||||||||
| 67 | 39.1 | neg | reduced spermiogenesis | pos | discrete interstitial edema | neg | neg | I | |
| 55 | n.d. | neg, int cont + | neg | autolytic changes | focal slightly thickened basement membrane of tubules, focal interstitial fibrosis | neg | neg | V | |
| 57 | 44.1 | neg | reduced spermiogenesis | pos | discrete interstitial edema | neg | neg | I | |
| 76 | norm | neg, int cont + | some spermatogonia and primary spermatocytes and spermatids | autolytic changes | focal slightly thickened basement membrane of tubules, focal interstitial fibrosis | neg | neg | IIA | |
| Mean | 63.75 | 41.6 | |||||||
| SEMINOMA | |||||||||
| n.d. | neg | pos | |||||||
| 58 | n.d. | neg | pos | ||||||
| 36 | n.d. | neg | ass | pos | |||||
| n.d. | neg | pos | |||||||
| 59 | n.d. | n.d | pos | ||||||
| n.d. | neg | pos | |||||||
| 51 | n.d. | n.d. | pos | ||||||
| 36 | n.d. | neg | pos | ||||||
| 27 | n.d. | neg | pos | ||||||
| 48 | n.d. | neg | pos | ||||||
| 51 | n.d. | neg | pos | ||||||
| 35 | n.d. | n.d. | pos | ||||||
| Mean | 39.4 | ||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goebel, H.; Koeditz, B.; Huerta, M.; Kameri, E.; Nestler, T.; Kamphausen, T.; Friemann, J.; Hamdorf, M.; Ohrmann, T.; Koehler, P.; et al. COVID-19 Infection Induce miR-371a-3p Upregulation Resulting in Influence on Male Fertility. Biomedicines 2022, 10, 858. https://doi.org/10.3390/biomedicines10040858
Goebel H, Koeditz B, Huerta M, Kameri E, Nestler T, Kamphausen T, Friemann J, Hamdorf M, Ohrmann T, Koehler P, et al. COVID-19 Infection Induce miR-371a-3p Upregulation Resulting in Influence on Male Fertility. Biomedicines. 2022; 10(4):858. https://doi.org/10.3390/biomedicines10040858
Chicago/Turabian StyleGoebel, Heike, Barbara Koeditz, Manuel Huerta, Ersen Kameri, Tim Nestler, Thomas Kamphausen, Johannes Friemann, Matthias Hamdorf, Timo Ohrmann, Philipp Koehler, and et al. 2022. "COVID-19 Infection Induce miR-371a-3p Upregulation Resulting in Influence on Male Fertility" Biomedicines 10, no. 4: 858. https://doi.org/10.3390/biomedicines10040858
APA StyleGoebel, H., Koeditz, B., Huerta, M., Kameri, E., Nestler, T., Kamphausen, T., Friemann, J., Hamdorf, M., Ohrmann, T., Koehler, P., Cornely, O. A., Montesinos-Rongen, M., Nicol, D., Schorle, H., Boor, P., Quaas, A., Pallasch, C., Heidenreich, A., & von Brandenstein, M. (2022). COVID-19 Infection Induce miR-371a-3p Upregulation Resulting in Influence on Male Fertility. Biomedicines, 10(4), 858. https://doi.org/10.3390/biomedicines10040858