
Dental-Pulp Stem Cells as a Therapeutic Strategy for Ischemic Stroke

 , and
, and
Abstract
:1. Introduction
2. Dental Pulp Stem Cells
3. Therapeutic Effects of DPSCs in Cerebral Ischemia
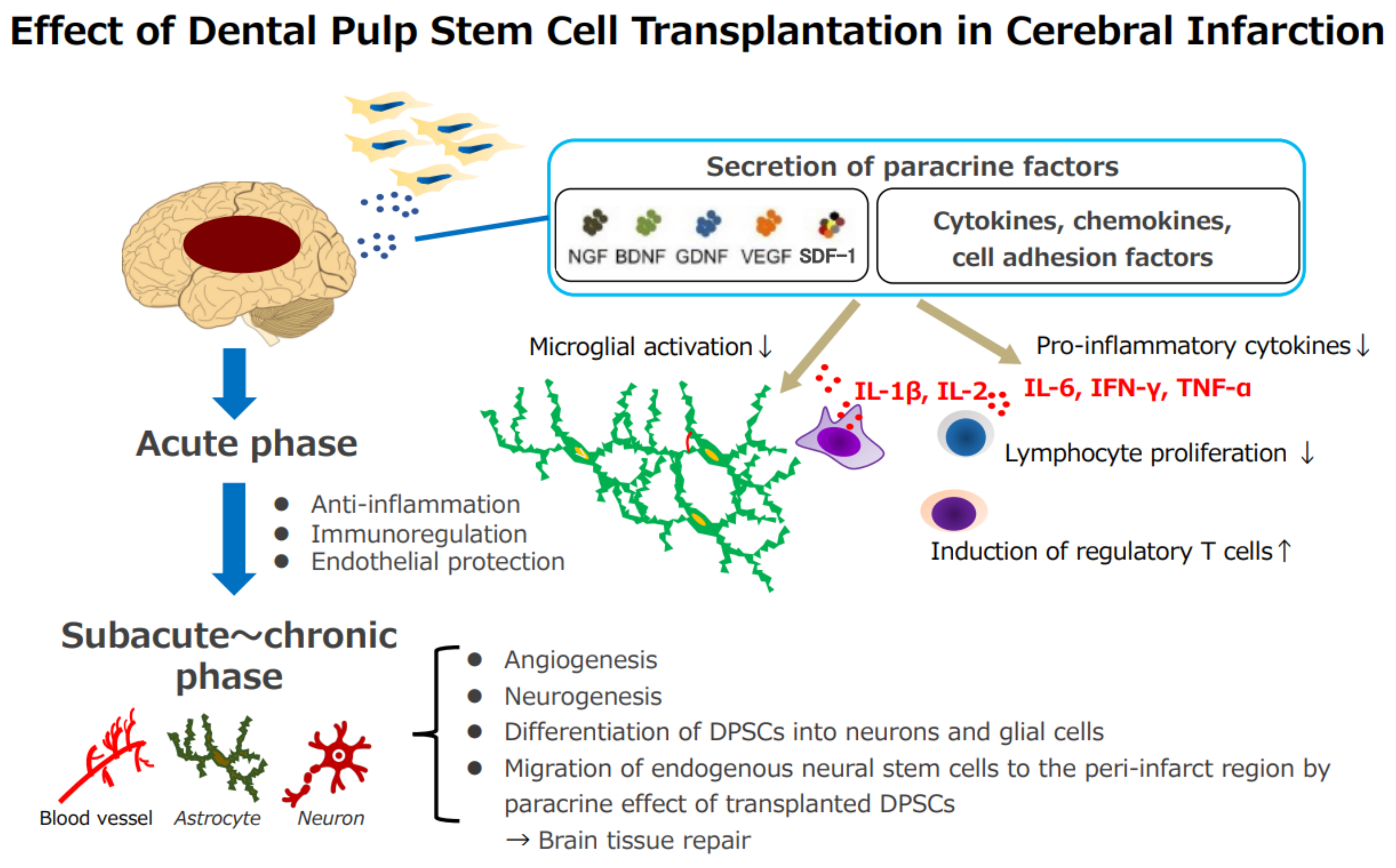
3.1. Neuroprotective Effects of DPSCs in Experimental Cerebral Ischemia
3.2. Therapeutic Mechanism
4. Enhancement of the Therapeutic Benefits of DPSCs by Genetic Modification
4.1. Genetic Modification of DPSCs
4.2. Enhanced Therapeutic Benefits of Gene-Modified DPSCs on Post-Ischemia/Reperfusion Injury
5. Clinical Studies of DPSCs for the Treatment of Stroke
5.1. Overview: Clinical Trials of Stem Cell Therapy for Stroke
5.2. Recent Clinical Trials of Stem Cells Other Than DPSCs against Stroke
5.3. Clinical Trials of DPSCs for the Treatment of Stroke
6. Perspective
Author Contributions
Funding
Conflicts of Interest
References
- Virani, S.S.; Alonso, A.; Benjamin, E.J.; Bittencourt, M.S.; Callaway, C.W.; Carson, A.P.; Chamberlain, A.M.; Chang, A.R.; Cheng, S.; Delling, F.N.; et al. Heart Disease and Stroke Statistics-2020 Update: A Report From the American Heart Association. Circulation 2020, 141, e139–e596. [Google Scholar] [CrossRef] [PubMed]
- Kuriakose, D.; Xiao, Z. Pathophysiology and Treatment of Stroke: Present Status and Future Perspectives. Int. J. Mol. Sci. 2020, 21, 7609. [Google Scholar] [CrossRef] [PubMed]
- Hacke, W.; Kaste, M.; Bluhmki, E.; Brozman, M.; Davalos, A.; Guidetti, D.; Larrue, V.; Lees, K.R.; Medeghri, Z.; Machnig, T.; et al. Thrombolysis with alteplase 3 to 4.5 hours after acute ischemic stroke. N. Engl. J. Med. 2008, 359, 1317–1329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, W.S.; Sung, G.; Saver, J.; Budzik, R.; Duckwiler, G.; Liebeskind, D.S.; Lutsep, H.L.; Rymer, M.M.; Higashida, R.T.; Starkman, S.; et al. Mechanical thrombectomy for acute ischemic stroke: Final results of the Multi MERCI trial. Stroke 2008, 39, 1205–1212. [Google Scholar] [CrossRef] [Green Version]
- Hao, L.; Zou, Z.; Tian, H.; Zhang, Y.; Zhou, H.; Liu, L. Stem cell-based therapies for ischemic stroke. Biomed Res. Int. 2014, 2014, 468748. [Google Scholar] [CrossRef] [Green Version]
- Friedenstein, A.J.; Petrakova, K.V.; Kurolesova, A.I.; Frolova, G.P. Heterotopic of bone marrow. Analysis of precursor cells for osteogenic and hematopoietic tissues. Transplantation 1968, 6, 230–247. [Google Scholar] [CrossRef]
- Zhang, W.; Walboomers, X.F.; Van Kuppevelt, T.H.; Daamen, W.F.; Van Damme, P.A.; Bian, Z.; Jansen, J.A. In vivo evaluation of human dental pulp stem cells differentiated towards multiple lineages. J. Tissue Eng. Regen. Med. 2008, 2, 117–125. [Google Scholar] [CrossRef]
- Fiedler, T.; Rabe, M.; Mundkowski, R.G.; Oehmcke-Hecht, S.; Peters, K. Adipose-derived mesenchymal stem cells release microvesicles with procoagulant activity. Int. J. Biochem. Cell Biol. 2018, 100, 49–53. [Google Scholar] [CrossRef]
- Li, J.Y.; Ren, K.K.; Zhang, W.J.; Xiao, L.; Wu, H.Y.; Liu, Q.Y.; Ding, T.; Zhang, X.C.; Nie, W.J.; Ke, Y.; et al. Human amniotic mesenchymal stem cells and their paracrine factors promote wound healing by inhibiting heat stress-induced skin cell apoptosis and enhancing their proliferation through activating PI3K/AKT signaling pathway. Stem Cell Res. 2019, 10, 247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, S.; Sevak, J.K.; Das, A.; Arimbasseri, G.A.; Bhatnagar, S.; Gopinath, S.D. Umbilical cord tissue is a robust source for mesenchymal stem cells with enhanced myogenic differentiation potential compared to cord blood. Sci. Rep. 2020, 10, 18978. [Google Scholar] [CrossRef]
- DelaRosa, O.; Lombardo, E. Modulation of adult mesenchymal stem cells activity by toll-like receptors: Implications on therapeutic potential. Mediat. Inflamm. 2010, 2010, 865601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Huang, J.; Gong, L.; Yu, D.; An, C.; Bunpetch, V.; Dai, J.; Huang, H.; Zou, X.; Ouyang, H.; et al. The Plasticity of Mesenchymal Stem Cells in Regulating Surface HLA-I. iScience 2019, 15, 66–78. [Google Scholar] [CrossRef] [Green Version]
- Muroi, K.; Miyamura, K.; Okada, M.; Yamashita, T.; Murata, M.; Ishikawa, T.; Uike, N.; Hidaka, M.; Kobayashi, R.; Imamura, M.; et al. Bone marrow-derived mesenchymal stem cells (JR-031) for steroid-refractory grade III or IV acute graft-versus-host disease: A phase II/III study. Int. J. Hematol. 2016, 103, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Savitz, S.I.; Yavagal, D.; Rappard, G.; Likosky, W.; Rutledge, N.; Graffagnino, C.; Alderazi, Y.; Elder, J.A.; Chen, P.R.; Budzik, R.F., Jr.; et al. A Phase 2 Randomized, Sham-Controlled Trial of Internal Carotid Artery Infusion of Autologous Bone Marrow-Derived ALD-401 Cells in Patients With Recent Stable Ischemic Stroke (RECOVER-Stroke). Circulation 2019, 139, 192–205. [Google Scholar] [CrossRef] [PubMed]
- Osanai, T.; Houkin, K.; Uchiyama, S.; Minematsu, K.; Taguchi, A.; Terasaka, S. Treatment evaluation of acute stroke for using in regenerative cell elements (TREASURE) trial: Rationale and design. Int. J. Stroke 2018, 13, 444–448. [Google Scholar] [CrossRef] [PubMed]
- Muir, K.W.; Bulters, D.; Willmot, M.; Sprigg, N.; Dixit, A.; Ward, N.; Tyrrell, P.; Majid, A.; Dunn, L.; Bath, P.; et al. Intracerebral implantation of human neural stem cells and motor recovery after stroke: Multicentre prospective single-arm study (PISCES-2). J. Neurol. Neurosurg. Psychiatry 2020, 91, 396–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nosrat, I.V.; Smith, C.A.; Mullally, P.; Olson, L.; Nosrat, C.A. Dental pulp cells provide neurotrophic support for dopaminergic neurons and differentiate into neurons in vitro; implications for tissue engineering and repair in the nervous system. Eur. J. Neurosci. 2004, 19, 2388–2398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gronthos, S.; Mankani, M.; Brahim, J.; Robey, P.G.; Shi, S. Postnatal human dental pulp stem cells (DPSCs) in vitro and in vivo. Proc. Natl. Acad. Sci. USA 2000, 97, 13625–13630. [Google Scholar] [CrossRef] [Green Version]
- Pierdomenico, L.; Bonsi, L.; Calvitti, M.; Rondelli, D.; Arpinati, M.; Chirumbolo, G.; Becchetti, E.; Marchionni, C.; Alviano, F.; Fossati, V.; et al. Multipotent mesenchymal stem cells with immunosuppressive activity can be easily isolated from dental pulp. Transplantation 2005, 80, 836–842. [Google Scholar] [CrossRef] [Green Version]
- Rajendran, R.; Gopal, S.; Masood, H.; Vivek, P.; Deb, K. Regenerative potential of dental pulp mesenchymal stem cells harvested from high caries patient’s teeth. J. Stem Cells 2013, 8, 25–41. [Google Scholar]
- Ponnaiyan, D.; Jegadeesan, V. Comparison of phenotype and differentiation marker gene expression profiles in human dental pulp and bone marrow mesenchymal stem cells. Eur. J. Dent. 2014, 8, 307–313. [Google Scholar] [CrossRef]
- Özdemir, A.T.; Özgül Özdemir, R.B.; Kırmaz, C.; Sarıboyacı, A.E.; Ünal Halbutoğlları, Z.S.; Özel, C.; Karaöz, E. The paracrine immunomodulatory interactions between the human dental pulp derived mesenchymal stem cells and CD4 T cell subsets. Cell. Immunol. 2016, 310, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Kurozumi, K.; Nakamura, K.; Tamiya, T.; Kawano, Y.; Ishii, K.; Kobune, M.; Hirai, S.; Uchida, H.; Sasaki, K.; Ito, Y.; et al. Mesenchymal stem cells that produce neurotrophic factors reduce ischemic damage in the rat middle cerebral artery occlusion model. Mol. Ther. 2005, 11, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhang, Z.G.; Li, Y.; Wang, L.; Xu, Y.X.; Gautam, S.C.; Lu, M.; Zhu, Z.; Chopp, M. Intravenous administration of human bone marrow stromal cells induces angiogenesis in the ischemic boundary zone after stroke in rats. Circ. Res. 2003, 92, 692–699. [Google Scholar] [CrossRef] [Green Version]
- Vu, Q.; Xie, K.; Eckert, M.; Zhao, W.; Cramer, S.C. Meta-analysis of preclinical studies of mesenchymal stromal cells for ischemic stroke. Neurology 2014, 82, 1277–1286. [Google Scholar] [CrossRef]
- Leong, W.K.; Lewis, M.D.; Koblar, S.A. Concise review: Preclinical studies on human cell-based therapy in rodent ischemic stroke models: Where are we now after a decade? Stem Cells 2013, 31, 1040–1043. [Google Scholar] [CrossRef] [PubMed]
- Nito, C.; Sowa, K.; Nakajima, M.; Sakamoto, Y.; Suda, S.; Nishiyama, Y.; Nakamura-Takahashi, A.; Nitahara-Kasahara, Y.; Ueda, M.; Okada, T.; et al. Transplantation of human dental pulp stem cells ameliorates brain damage following acute cerebral ischemia. Biomed. Pharmacother. 2018, 108, 1005–1014. [Google Scholar] [CrossRef]
- Sugiyama, M.; Iohara, K.; Wakita, H.; Hattori, H.; Ueda, M.; Matsushita, K.; Nakashima, M. Dental pulp-derived CD31(-)/CD146(-) side population stem/progenitor cells enhance recovery of focal cerebral ischemia in rats. Tissue Eng. Part A 2011, 17, 1303–1311. [Google Scholar] [CrossRef]
- Leong, W.K.; Henshall, T.L.; Arthur, A.; Kremer, K.L.; Lewis, M.D.; Helps, S.C.; Field, J.; Hamilton-Bruce, M.A.; Warming, S.; Manavis, J.; et al. Human adult dental pulp stem cells enhance poststroke functional recovery through non-neural replacement mechanisms. Stem Cells Transl. Med. 2012, 1, 177–187. [Google Scholar] [CrossRef]
- Song, M.; Lee, J.H.; Bae, J.; Bu, Y.; Kim, E.C. Human Dental Pulp Stem Cells Are More Effective Than Human Bone Marrow-Derived Mesenchymal Stem Cells in Cerebral Ischemic Injury. Cell Transplant. 2017, 26, 1001–1016. [Google Scholar] [CrossRef]
- Zhang, X.; Zhou, Y.; Li, H.; Wang, R.; Yang, D.; Li, B.; Cao, X.; Fu, J. Transplanted Dental Pulp Stem Cells Migrate to Injured Area and Express Neural Markers in a Rat Model of Cerebral Ischemia. Cell. Physiol. Biochem. 2018, 45, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhou, Y.; Li, H.; Wang, R.; Yang, D.; Li, B.; Fu, J. Intravenous administration of DPSCs and BDNF improves neurological performance in rats with focal cerebral ischemia. Int. J. Mol. Med. 2018, 41, 3185–3194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, T.; Xu, W.; Chen, H.; Li, S.; Dou, R.; Shen, H.; Liu, X.; Liu, X.; Hong, Y.; He, J. Comparison of the differentiation of dental pulp stem cells and periodontal ligament stem cells into neuron-like cells and their effects on focal cerebral ischemia. Acta Biochim. Biophys. Sin. 2020, 52, 1016–1029. [Google Scholar] [CrossRef] [PubMed]
- Kumasaka, A.; Kanazawa, K.; Ohke, H.; Miura, I.; Miura, Y. Post-ischemic Intravenous Administration of Allogeneic Dental Pulp-Derived Neurosphere Cells Ameliorated Outcomes of Severe Forebrain Ischemia in Rats. Neurocrit. Care 2017, 26, 133–142. [Google Scholar] [CrossRef]
- Sowa, K.; Nito, C.; Nakajima, M.; Suda, S.; Nishiyama, Y.; Sakamoto, Y.; Nitahara-Kasahara, Y.; Nakamura-Takahashi, A.; Ueda, M.; Kimura, K.; et al. Impact of Dental Pulp Stem Cells Overexpressing Hepatocyte Growth Factor after Cerebral Ischemia/Reperfusion in Rats. Mol. Ther. Methods Clin. Dev. 2018, 10, 281–290. [Google Scholar] [CrossRef] [Green Version]
- Yew, W.P.; Djukic, N.D.; Jayaseelan, J.S.P.; Kaidonis, X.; Kremer, K.L.; Choy, F.C.; Woodman, R.J.; Koblar, S.A.; Sims, N.R. Delayed Treatment with Human Dental Pulp Stem Cells Accelerates Functional Recovery and Modifies Responses of Peri-Infarct Astrocytes Following Photothrombotic Stroke in Rats. Cell Transplant. 2021, 30, 1–13. [Google Scholar] [CrossRef]
- Matsumura, H.; Marushima, A.; Ishikawa, H.; Toyomura, J.; Ohyama, A.; Watanabe, M.; Takaoka, S.; Bukawa, H.; Matsumura, A.; Matsumaru, Y.; et al. Induced Neural Cells from Human Dental Pulp Ameliorate Functional Recovery in a Murine Model of Cerebral Infarction. Stem Cell Rev. Rep. 2021, 18, 595–608. [Google Scholar] [CrossRef]
- Li, S.; Luo, L.; He, Y.; Li, R.; Xiang, Y.; Xing, Z.; Li, Y.; Albashari, A.A.; Liao, X.; Zhang, K.; et al. Dental pulp stem cell-derived exosomes alleviate cerebral ischaemia-reperfusion injury through suppressing inflammatory response. Cell Prolif. 2021, 54, e13093. [Google Scholar] [CrossRef]
- Fang, C.Z.; Yang, Y.J.; Wang, Q.H.; Yao, Y.; Zhang, X.Y.; He, X.H. Intraventricular injection of human dental pulp stem cells improves hypoxic-ischemic brain damage in neonatal rats. PLoS ONE 2013, 8, e66748. [Google Scholar] [CrossRef] [Green Version]
- Ge, J.; Guo, L.; Wang, S.; Zhang, Y.; Cai, T.; Zhao, R.C.; Wu, Y. The size of mesenchymal stem cells is a significant cause of vascular obstructions and stroke. Stem Cell Rev. 2014, 10, 295–303. [Google Scholar] [CrossRef]
- Cui, L.L.; Kerkelä, E.; Bakreen, A.; Nitzsche, F.; Andrzejewska, A.; Nowakowski, A.; Janowski, M.; Walczak, P.; Boltze, J.; Lukomska, B.; et al. The cerebral embolism evoked by intra-arterial delivery of allogeneic bone marrow mesenchymal stem cells in rats is related to cell dose and infusion velocity. Stem Cell Res. 2015, 6, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakajima, M.; Nito, C.; Sowa, K.; Suda, S.; Nishiyama, Y.; Nakamura-Takahashi, A.; Nitahara-Kasahara, Y.; Imagawa, K.; Hirato, T.; Ueda, M.; et al. Mesenchymal Stem Cells Overexpressing Interleukin-10 Promote Neuroprotection in Experimental Acute Ischemic Stroke. Mol. Ther. Methods Clin. Dev. 2017, 6, 102–111. [Google Scholar] [CrossRef] [Green Version]
- Inoue, T.; Sugiyama, M.; Hattori, H.; Wakita, H.; Wakabayashi, T.; Ueda, M. Stem cells from human exfoliated deciduous tooth-derived conditioned medium enhance recovery of focal cerebral ischemia in rats. Tissue Eng. Part A 2013, 19, 24–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalladka, D.; Muir, K.W. Stem cell therapy in stroke: Designing clinical trials. Neurochem. Int. 2011, 59, 367–370. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, T.B.; Duarte, A.S.; Longhini, A.L.; Pradella, F.; Farias, A.S.; Luzo, A.C.; Oliveira, A.L.; Olalla Saad, S.T. Neuroprotection and immunomodulation by xenografted human mesenchymal stem cells following spinal cord ventral root avulsion. Sci. Rep. 2015, 5, 16167. [Google Scholar] [CrossRef]
- Martire, A.; Bedada, F.B.; Uchida, S.; Poling, J.; Kruger, M.; Warnecke, H.; Richter, M.; Kubin, T.; Herold, S.; Braun, T. Mesenchymal stem cells attenuate inflammatory processes in the heart and lung via inhibition of TNF signaling. Basic Res. Cardiol. 2016, 111, 54. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.A.; Kim, B.I.; Jo, C.H.; Choi, C.W.; Kim, E.K.; Kim, H.S.; Yoon, K.S.; Choi, J.H. Mesenchymal stem-cell transplantation for hypoxic-ischemic brain injury in neonatal rat model. Pediatr. Res. 2010, 67, 42–46. [Google Scholar] [CrossRef] [Green Version]
- Ajmo, C.T., Jr.; Vernon, D.O.; Collier, L.; Hall, A.A.; Garbuzova-Davis, S.; Willing, A.; Pennypacker, K.R. The spleen contributes to stroke-induced neurodegeneration. J. Neurosci. Res. 2008, 86, 2227–2234. [Google Scholar] [CrossRef] [Green Version]
- Shichita, T.; Ago, T.; Kamouchi, M.; Kitazono, T.; Yoshimura, A.; Ooboshi, H. Novel therapeutic strategies targeting innate immune responses and early inflammation after stroke. J. Neurochem. 2012, 123 (Suppl. S2), 29–38. [Google Scholar] [CrossRef]
- Matsushita, K.; Motani, R.; Sakuta, T.; Yamaguchi, N.; Koga, T.; Matsuo, K.; Nagaoka, S.; Abeyama, K.; Maruyama, I.; Torii, M. The role of vascular endothelial growth factor in human dental pulp cells: Induction of chemotaxis, proliferation, and differentiation and activation of the AP-1-dependent signaling pathway. J. Dent. Res. 2000, 79, 1596–1603. [Google Scholar] [CrossRef]
- Nosrat, I.V.; Widenfalk, J.; Olson, L.; Nosrat, C.A. Dental pulp cells produce neurotrophic factors, interact with trigeminal neurons in vitro, and rescue motoneurons after spinal cord injury. Dev. Biol. 2001, 238, 120–132. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Ye, R.; Yan, T.; Yu, S.P.; Wei, L.; Xu, G.; Fan, X.; Jiang, Y.; Stetler, R.A.; Liu, G.; et al. Cell based therapies for ischemic stroke: From basic science to bedside. Prog. Neurobiol. 2014, 115, 92–115. [Google Scholar] [CrossRef] [Green Version]
- Liesz, A.; Bauer, A.; Hoheisel, J.D.; Veltkamp, R. Intracerebral interleukin-10 injection modulates post-ischemic neuroinflammation: An experimental microarray study. Neurosci. Lett. 2014, 579, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.G.; Zhang, L.; Jiang, Q.; Zhang, R.; Davies, K.; Powers, C.; Bruggen, N.; Chopp, M. VEGF enhances angiogenesis and promotes blood-brain barrier leakage in the ischemic brain. J. Clin. Investig. 2000, 106, 829–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Király, M.; Kádár, K.; Horváthy, D.B.; Nardai, P.; Rácz, G.Z.; Lacza, Z.; Varga, G.; Gerber, G. Integration of neuronally predifferentiated human dental pulp stem cells into rat brain in vivo. Neurochem. Int. 2011, 59, 371–381. [Google Scholar] [CrossRef]
- Jin, K.; Sun, Y.; Xie, L.; Mao, X.O.; Childs, J.; Peel, A.; Logvinova, A.; Banwait, S.; Greenberg, D.A. Comparison of ischemia-directed migration of neural precursor cells after intrastriatal, intraventricular, or intravenous transplantation in the rat. Neurobiol. Dis. 2005, 18, 366–374. [Google Scholar] [CrossRef] [PubMed]
- Gancheva, M.R.; Kremer, K.L.; Gronthos, S.; Koblar, S.A. Using Dental Pulp Stem Cells for Stroke Therapy. Front. Neurol. 2019, 10, 422. [Google Scholar] [CrossRef]
- Boltze, J.; Abe, K.; Clarkson, A.N.; Detante, O.; Pimentel-Coelho, P.M.; Rosado-de-Castro, P.H.; Janowski, M. Editorial: Cell-based Therapies for Stroke: Promising Solution or Dead End? Front. Neurol. 2020, 11, 171. [Google Scholar] [CrossRef]
- Suda, S.; Nito, C.; Yokobori, S.; Sakamoto, Y.; Nakajima, M.; Sowa, K.; Obinata, H.; Sasaki, K.; Savitz, S.I.; Kimura, K. Recent Advances in Cell-Based Therapies for Ischemic Stroke. Int. J. Mol. Sci. 2020, 21, 6718. [Google Scholar] [CrossRef]
- Nie, W.B.; Zhang, D.; Wang, L.S. Growth Factor Gene-Modified Mesenchymal Stem Cells in Tissue Regeneration. Drug Des. Dev. Ther. 2020, 14, 1241–1256. [Google Scholar] [CrossRef] [Green Version]
- Nonaka, K.; Han, X.; Kato, H.; Sato, H.; Yamaza, H.; Hirofuji, Y.; Masuda, K. Novel gain-of-function mutation of TRPV4 associated with accelerated chondrogenic differentiation of dental pulp stem cells derived from a patient with metatropic dysplasia. Biochem. Biophys. Rep. 2019, 19, 100648. [Google Scholar] [CrossRef] [PubMed]
- Egbuniwe, O.; Grant, A.D.; Renton, T.; Di Silvio, L. Phenotype-independent effects of retroviral transduction in human dental pulp stem cells. Macromol. Biosci. 2013, 13, 851–859. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhao, Y.; Liu, X.; Chen, Y.; Liu, P.; Zhao, L. Effect of SOX2 on odontoblast differentiation of dental pulp stem cells. Mol. Med. Rep. 2017, 16, 9659–9663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, H.; Li, W.; Yang, H.; Cao, Y.; Ge, L.; Shi, R.; Fan, Z.; Dong, R.; Zhang, C. FAM96B inhibits the senescence of dental pulp stem cells. Cell Biol. Int. 2020, 44, 1193–1203. [Google Scholar] [CrossRef]
- Li, N.; Zhang, Y.; Nepal, N.; Li, G.; Yang, N.; Chen, H.; Lin, Q.; Ji, X.; Zhang, S.; Jin, S. Dental pulp stem cells overexpressing hepatocyte growth factor facilitate the repair of DSS-induced ulcerative colitis. Stem Cell Res. 2021, 12, 30. [Google Scholar] [CrossRef]
- Maciejewska, I.; Sakowicz-Burkiewicz, M.; Pawelczyk, T. Id1 Expression Level Determines the Differentiation of Human Dental Pulp Stem Cells. J. Dent. Res. 2014, 93, 576–581. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.; Kong, F.; Liu, C.; Dai, S.; Zhang, Y.; Xiao, F.; Zhang, H.; Wu, C.T.; Wang, H. Pulp stem cells with hepatocyte growth factor overexpression exhibit dual effects in rheumatoid arthritis. Stem Cell Res. 2020, 11, 229. [Google Scholar] [CrossRef]
- Nitahara-Kasahara, Y.; Kuraoka, M.; Oda, Y.; Hayashita-Kinoh, H.; Takeda, S.; Okada, T. Enhanced cell survival and therapeutic benefits of IL-10-expressing multipotent mesenchymal stromal cells for muscular dystrophy. Stem Cell Res. 2021, 12, 105. [Google Scholar] [CrossRef]
- Chan, J.; O’Donoghue, K.; de la Fuente, J.; Roberts, I.A.; Kumar, S.; Morgan, J.E.; Fisk, N.M. Human fetal mesenchymal stem cells as vehicles for gene delivery. Stem Cells 2005, 23, 93–102. [Google Scholar] [CrossRef]
- Santiago-Torres, J.E.; Lovasz, R.; Bertone, A.L. Fetal vs adult mesenchymal stem cells achieve greater gene expression, but less osteoinduction. World J. Stem Cells 2015, 7, 223–234. [Google Scholar] [CrossRef]
- Jooss, K.; Yang, Y.; Fisher, K.J.; Wilson, J.M. Transduction of dendritic cells by DNA viral vectors directs the immune response to transgene products in muscle fibers. J. Virol. 1998, 72, 4212–4223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jooss, K.; Ertl, H.C.; Wilson, J.M. Cytotoxic T-lymphocyte target proteins and their major histocompatibility complex class I restriction in response to adenovirus vectors delivered to mouse liver. J. Virol. 1998, 72, 2945–2954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozawa, K.; Sato, K.; Oh, I.; Ozaki, K.; Uchibori, R.; Obara, Y.; Kikuchi, Y.; Ito, T.; Okada, T.; Urabe, M.; et al. Cell and gene therapy using mesenchymal stem cells (MSCs). J. Autoimmun. 2008, 30, 121–127. [Google Scholar] [CrossRef]
- Okada, T.; Takeda, S. Current Challenges and Future Directions in Recombinant AAV-Mediated Gene Therapy of Duchenne Muscular Dystrophy. Pharmaceuticals 2013, 6, 813–836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okada, H.; Ishibashi, H.; Hayashita-Kinoh, H.; Chiyo, T.; Nitahara-Kasahara, Y.; Baba, Y.; Watanabe, S.; Takeda, S.; Okada, T. Robust Long-term Transduction of Common Marmoset Neuromuscular Tissue With rAAV1 and rAAV9. Mol. Nucleic Acids 2013, 2, e95. [Google Scholar] [CrossRef] [PubMed]
- Zubkova, E.S.; Beloglazova, I.B.; Ratner, E.I.; Dyikanov, D.T.; Dergilev, K.V.; Menshikov, M.Y.; Parfyonova, Y.V. Transduction of rat and human adipose-tissue derived mesenchymal stromal cells by adeno-associated viral vector serotype DJ. Biol. Open 2021, 10, bio058461. [Google Scholar] [CrossRef] [PubMed]
- Sehara, Y.; Fujimoto, K.I.; Ikeguchi, K.; Katakai, Y.; Ono, F.; Takino, N.; Ito, M.; Ozawa, K.; Muramatsu, S.I. Persistent Expression of Dopamine-Synthesizing Enzymes 15 Years After Gene Transfer in a Primate Model of Parkinson’s Disease. Hum. Gene Clin. Dev. 2017, 28, 74–79. [Google Scholar] [CrossRef]
- Lakhan, R.; Baylink, D.J.; Lau, K.H.; Tang, X.; Sheng, M.H.; Rundle, C.H.; Qin, X. Local administration of AAV-DJ pseudoserotype expressing COX2 provided early onset of transgene expression and promoted bone fracture healing in mice. Gene 2015, 22, 721–728. [Google Scholar] [CrossRef] [Green Version]
- Shimamura, M.; Sato, N.; Oshima, K.; Aoki, M.; Kurinami, H.; Waguri, S.; Uchiyama, Y.; Ogihara, T.; Kaneda, Y.; Morishita, R. Novel therapeutic strategy to treat brain ischemia: Overexpression of hepatocyte growth factor gene reduced ischemic injury without cerebral edema in rat model. Circulation 2004, 109, 424–431. [Google Scholar] [CrossRef]
- Wang, L.S.; Wang, H.; Zhang, Q.L.; Yang, Z.J.; Kong, F.X.; Wu, C.T. Hepatocyte Growth Factor Gene Therapy for Ischemic Diseases. Hum. Gene 2018, 29, 413–423. [Google Scholar] [CrossRef]
- Kawabori, M.; Shichinohe, H.; Kuroda, S.; Houkin, K. Clinical Trials of Stem Cell Therapy for Cerebral Ischemic Stroke. Int. J. Mol. Sci. 2020, 21, 7380. [Google Scholar] [CrossRef] [PubMed]
- Hatakeyama, M.; Ninomiya, I.; Otsu, Y.; Omae, K.; Kimura, Y.; Onodera, O.; Fukushima, M.; Shimohata, T.; Kanazawa, M. Cell Therapies under Clinical Trials and Polarized Cell Therapies in Pre-Clinical Studies to Treat Ischemic Stroke and Neurological Diseases: A Literature Review. Int. J. Mol. Sci. 2020, 21, 6194. [Google Scholar] [CrossRef] [PubMed]
- Shichinohe, H.; Houkin, K. Cell Therapy for Stroke: Review of Previous Clinical Trials and Introduction of Our New Trials. Neurol. Med. Chir. 2016, 56, 592–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houkin, K.; Shichinohe, H.; Abe, K.; Arato, T.; Dezawa, M.; Honmou, O.; Horie, N.; Katayama, Y.; Kudo, K.; Kuroda, S.; et al. Accelerating Cell Therapy for Stroke in Japan: Regulatory Framework and Guidelines on Development of Cell-Based Products. Stroke 2018, 49, e145–e152. [Google Scholar] [CrossRef]
- Abe, K.; Yamashita, T.; Takizawa, S.; Kuroda, S.; Kinouchi, H.; Kawahara, N. Stem cell therapy for cerebral ischemia: From basic science to clinical applications. J. Cereb. Blood Flow Metab. 2012, 32, 1317–1331. [Google Scholar] [CrossRef] [Green Version]
- Kondziolka, D.; Steinberg, G.K.; Wechsler, L.; Meltzer, C.C.; Elder, E.; Gebel, J.; Decesare, S.; Jovin, T.; Zafonte, R.; Lebowitz, J.; et al. Neurotransplantation for patients with subcortical motor stroke: A phase 2 randomized trial. J. Neurosurg. 2005, 103, 38–45. [Google Scholar] [CrossRef]
- Savitz, S.I.; Dinsmore, J.; Wu, J.; Henderson, G.V.; Stieg, P.; Caplan, L.R. Neurotransplantation of fetal porcine cells in patients with basal ganglia infarcts: A preliminary safety and feasibility study. Cereb. Dis. 2005, 20, 101–107. [Google Scholar] [CrossRef]
- Lee, J.; Chang, W.H.; Chung, J.-W.; Kim, S.J.; Kim, S.-K.; Lee, J.S.; Sohn, S.-I.; Kim, Y.-H.; Bang, O.Y. Efficacy of Intravenous Mesenchymal Stem Cells for Motor Recovery After Ischemic Stroke: A Neuroimaging Study. Stroke 2022, 53, 20–28. [Google Scholar] [CrossRef]
- Jaillard, A.; Hommel, M.; Moisan, A.; Zeffiro, T.A.; Favre-Wiki, I.M.; Barbieux-Guillot, M.; Vadot, W.; Marcel, S.; Lamalle, L.; Grand, S.; et al. Autologous Mesenchymal Stem Cells Improve Motor Recovery in Subacute Ischemic Stroke: A Randomized Clinical Trial. Transl. Stroke Res. 2020, 11, 910–923. [Google Scholar] [CrossRef]
- Haque, M.E.; Gabr, R.E.; George, S.D.; Boren, S.B.; Vahidy, F.S.; Zhang, X.; Arevalo, O.D.; Alderman, S.; Narayana, P.A.; Hasan, K.M.; et al. Serial Cerebral Metabolic Changes in Patients With Ischemic Stroke Treated With Autologous Bone Marrow Derived Mononuclear Cells. Front. Neurol. 2019, 10, 141. [Google Scholar] [CrossRef] [Green Version]
- Haque, M.E.; Hasan, K.M.; George, S.; Sitton, C.; Boren, S.; Arevalo, O.D.; Vahidy, F.; Zhang, X.; Cox, C.S., Jr.; Alderman, S.; et al. Longitudinal neuroimaging evaluation of the corticospinal tract in patients with stroke treated with autologous bone marrow cells. Stem Cells Transl. Med. 2021, 10, 943–955. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.J.; Niizuma, K.; Mokin, M.; Dezawa, M.; Borlongan, C.V. Cell-Based Therapy for Stroke: Musing With Muse Cells. Stroke 2020, 51, 2854–2862. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, T.; Kushida, Y.; Abe, K.; Dezawa, M. Non-Tumorigenic Pluripotent Reparative Muse Cells Provide a New Therapeutic Approach for Neurologic Diseases. Cells 2021, 10, 961. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Sato, Y.; Kushida, Y.; Tsuji, M.; Wakao, S.; Ueda, K.; Imai, K.; Iitani, Y.; Shimizu, S.; Hida, H.; et al. Intravenously delivered multilineage-differentiating stress enduring cells dampen excessive glutamate metabolism and microglial activation in experimental perinatal hypoxic ischemic encephalopathy. J. Cereb. Blood Flow Metab. 2021, 41, 1707–1720. [Google Scholar] [CrossRef]
- Uchida, H.; Niizuma, K.; Kushida, Y.; Wakao, S.; Tominaga, T.; Borlongan, C.V.; Dezawa, M. Human Muse Cells Reconstruct Neuronal Circuitry in Subacute Lacunar Stroke Model. Stroke 2017, 48, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Niizuma, K.; Borlongan, C.V.; Tominaga, T. Application of Muse Cell Therapy to Stroke. Adv. Exp. Med. Biol. 2018, 1103, 167–186. [Google Scholar] [CrossRef]
- Kuroda, S.; Koh, M.; Hori, E.; Hayakawa, Y.; Akai, T. Muse Cell: A New Paradigm for Cell Therapy and Regenerative Homeostasis in Ischemic Stroke. Adv. Exp. Med. Biol. 2018, 1103, 187–198. [Google Scholar] [CrossRef]
- Nagpal, A.; Kremer, K.L.; Hamilton-Bruce, M.A.; Kaidonis, X.; Milton, A.G.; Levi, C.; Shi, S.; Carey, L.; Hillier, S.; Rose, M.; et al. TOOTH (The Open study Of dental pulp stem cell Therapy in Humans): Study protocol for evaluating safety and feasibility of autologous human adult dental pulp stem cell therapy in patients with chronic disability after stroke. Int. J. Stroke 2016, 11, 575–585. [Google Scholar] [CrossRef]
- Savitz, S.I.; Baron, J.C.; Fisher, M.; Consortium, S.X. Stroke Treatment Academic Industry Roundtable X: Brain Cytoprotection Therapies in the Reperfusion Era. Stroke 2019, 50, 1026–1031. [Google Scholar] [CrossRef]
- Boltze, J.; Modo, M.M.; Mays, R.W.; Taguchi, A.; Jolkkonen, J.; Savitz, S.I.; Consortium, S. Stem Cells as an Emerging Paradigm in Stroke 4: Advancing and Accelerating Preclinical Research. Stroke 2019, 50, 3299–3306. [Google Scholar] [CrossRef]

{kind=link}
| Cell Type | Number of Cells | Animal Model | Delivery Timing | Delivery Route | Results | Reference |
|---|---|---|---|---|---|---|
| Porcine dental pulp side population, progenitor cells | 1 × 106 | Rat MCAO (2 h) | 24 h postischemia induction | Intracerebral | Recovered motor function and infarct volume Differentiation of endogenous neuronal progenitor cells and induction of vasculogenesis | Sugiyama et al. 2011 [28] |
| Human DPSCs | 6 × 105 | Rat MCAO (2 h) | 24 h postischemia induction | Intracerebral (striatum and cortex) | Differentiation into astrocytes Neuroprotection Functional outcomes | Leong et al. 2012 [29] |
| Human DPSCs | 4 × 106 | Rat MCAO (2 h) | 24 h postischemia induction | Intravenous (tail vein) | Reduced infarct volume Improved neurological outcome Differentiation into astrocytes and neuron-like cells Promotion of angiogenesis and inhibition of astrocytes | Song et al. 2017 [30] |
| Rat dental pulp-derived neurospheres | 1 × 106 | Rat severe forebrain ischemia (11 min) | 3 h postischemia induction | Intravenous (tail vein) | Increased survival rate Improved cognitive functional recovery Reduced neuronal death | Kumasaka et al. 2017 [34] |
| Human DPSCs | 1 × 106 | Rat MCAO (90 min) | Immediately or 3 h postischemia | Intravenous (tail vein) | Reduced infarct volume Improved neurological outcome Inflammation modulation | Nito et al. 2018 [27] |
| Human DPSCs, HGF-transfected DPSCs | 1 × 106 | Rat MCAO (90 min) | Immediately postischemia | Intravenous (tail vein) | Reduced infarct and edema volume Improved neurological outcome Inflammation modulation Promotion of angiogenesis | Sowa et al. 2018 [35] |
| Rat DPSCs, combination with BDNF | 1 × 107 | Rat MCAO (2 h) | 24 h postischemia induction | Intravenous (tail vein) | Reduced infarct and edema volume Differentiation into neuron-like cells | Zhang et al. 2018 [32] |
| Rat DPSCs, combination with BDNF | 1 × 106 | Rat MCAO (2 h) | 24 h postischemia induction | Intravenous (tail vein) | Reduced infarct and edema volume Improved neurological outcome Differentiation into neuronal progenitor and neuron-like cells Triggered neurogenesis | Zhang et al. 2018 [31] |
| Human DPSC, PDLSCs | 1× 106 | Rat MCAO (2 h) | 24 h postischemia induction | Intravenous (tail vein) | Reduced cerebral infarct size Recovery of neurological function | Wu et al. 2020 [33] |
| Human DPSCs | 4 × 105 | Rat photothrombosis induces permanent focal ischemia | 3 days postunilateral photothrombotic stroke induction | Intracerebral | Success in a skilled forelimb reaching test Decreased reactive astrogliosis | Yew et al. 2021 [36] |
| Human DPSCs Induced neural cells | 1 × 104 1 × 105 1 × 106 | Murine MCAO | 5 days postischemia induction | Intravenous | Functional recovery Differentiation into neurons and glia cells | Matsumura et al. 2021 [37] |
| Human DPSCs Derived Exosome | Mice MCAO (2 h) | 4 h postischemia induction | Intravenous | Reduced infarct and edema volume Improved neurological outcome Inflammation modulation | Li et al. 2021 [38] |
| 1. MCA ischemic stroke |
| 2. Moderate severity chronic disability—stable level (a modified Rankin Score of 2–4) of chronic motor, sensory, and/or language disability for at least 6 months prior to selection. Dominant hemisphere MCA stroke survivors with aphasia are required to attain an aphasia quotient score of 33–70 on the Western Aphasia Battery (WAB-AQ) to participate |
| 3. Good cognitive function—the participant must achieve a Mini Mental State Examination score of 24 or more. Participants with aphasia must score above 23 on the Raven’s Colored Progressive Matrices |
| 4. All participants must pass a Mini International Neuropsychiatric Interview; in those with aphasia, a score of less than 17 on the Stroke Aphasia Depression Questionaire-21 is required |
| 5. Healthy teeth to grow sufficient autologous dental pulp stem cells |
| Inclusion Criteria |
|---|
| 1. Clinical diagnosis of anterior circulation ischemic stroke by nuclear MRI or computed tomography |
| 2. NIHSS score ≥ 5 to ≤20 at screening |
| 3. Onset of ischemic stroke must have occurred within 48 h prior to the start of administration of the study product |
| 4. A modified Rankin Scale of 0 or 1, by either self-report or family report, prior to the onset of ischemic stroke |
| Exclusion criteria |
| 1. Presence of intracranial hemorrhagic change diagnosed by MRI, which is judged to be clinically important by the investigator at screening |
| 2. Alzheimer’s disease or other dementia, Parkinson’s disease, or any other neurological disorder |
| 3. Planned revascularization treatment including carotid endarterectomy and stenting, by the end of the evaluation (Day 91) |
| 4. After eligibility assessment at screening, the investigator will assess NIHSS again ≥4 h after the assessment at screening. Subjects meeting one or more of the following criteria will be excluded: NIHSS score < 4 or ≥21; change in NIHSS score from screening ≥ 5. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nito, C.; Suda, S.; Nitahara-Kasahara, Y.; Okada, T.; Kimura, K. Dental-Pulp Stem Cells as a Therapeutic Strategy for Ischemic Stroke. Biomedicines 2022, 10, 737. https://doi.org/10.3390/biomedicines10040737
Nito C, Suda S, Nitahara-Kasahara Y, Okada T, Kimura K. Dental-Pulp Stem Cells as a Therapeutic Strategy for Ischemic Stroke. Biomedicines. 2022; 10(4):737. https://doi.org/10.3390/biomedicines10040737
Chicago/Turabian StyleNito, Chikako, Satoshi Suda, Yuko Nitahara-Kasahara, Takashi Okada, and Kazumi Kimura. 2022. "Dental-Pulp Stem Cells as a Therapeutic Strategy for Ischemic Stroke" Biomedicines 10, no. 4: 737. https://doi.org/10.3390/biomedicines10040737
APA StyleNito, C., Suda, S., Nitahara-Kasahara, Y., Okada, T., & Kimura, K. (2022). Dental-Pulp Stem Cells as a Therapeutic Strategy for Ischemic Stroke. Biomedicines, 10(4), 737. https://doi.org/10.3390/biomedicines10040737