
Cell Shortening and Calcium Homeostasis Analysis in Adult Cardiomyocytes via a New Software Tool

 , , , and
, , , and
Abstract

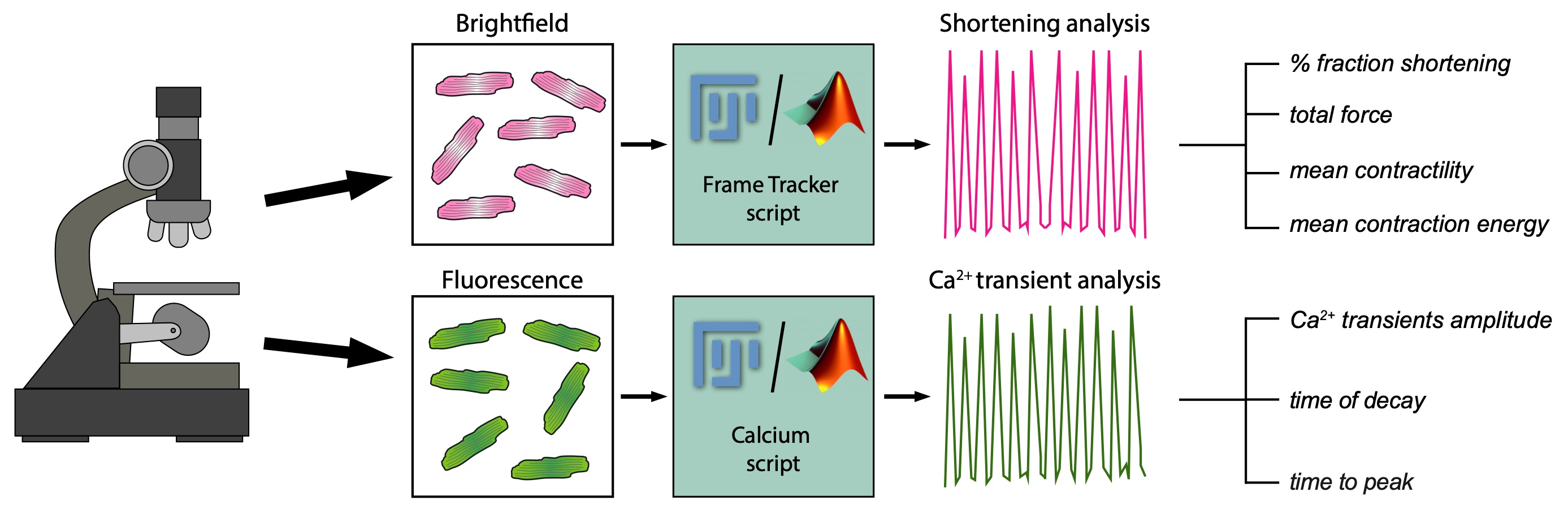{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Mouse Husbandry
2.2. Isolation of Primary Adult Cardiomyocytes
2.3. Calcium Transient and Contraction
2.4. Statistical Analysis
3. Results
3.1. Analysis of Cell Shortening and Kinematics/Dynamics of Primary Isolated Adult Cardiomyocytes
3.2. Analysis of Ca2+ Transient in Primarily Isolated Adult Cardiomyocytes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bers, D.M. Calcium Cycling and Signaling in Cardiac Myocytes. Annu. Rev. Physiol. 2008, 70, 23–49. [Google Scholar] [CrossRef] [PubMed]
- Bers, D.M. Calcium fluxes involved in control of cardiac myocyte contraction. Circ. Res. 2000, 87, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Lanner, J.T.; Georgiou, D.K.; Joshi, A.D.; Hamilton, S.L. Ryanodine receptors: Structure, expression, molecular details, and function in calcium release. Cold Spring Harb. Perspect. Biol. 2010, 2, a003996. [Google Scholar] [CrossRef] [PubMed]
- Berridge, M.J.; Bootman, M.D.; Roderick, H.L. Calcium signalling: Dynamics, homeostasis and remodelling. Nat. Rev. Mol. Cell Biol. 2003, 4, 517–529. [Google Scholar] [CrossRef]
- Eisner, D.A.; Caldwell, J.L.; Kistamás, K.; Trafford, A.W. Calcium and Excitation-Contraction Coupling in the Heart. Circ. Res. 2017, 121, 181–195. [Google Scholar] [CrossRef]
- Kranias, E.G.; Bers, D.M. Calcium and cardiomyopathies. Subcell. Biochem. 2007, 45, 523–537. [Google Scholar] [CrossRef]
- Molkentin, J.D. Calcineurin-NFAT signaling regulates the cardiac hypertrophic response in coordination with the MAPKs. Cardiovasc. Res. 2004, 63, 467–475. [Google Scholar] [CrossRef]
- Marx, S.O.; Reiken, S.; Hisamatsu, Y.; Jayaraman, T.; Burkhoff, D.; Rosemblit, N.; Marks, A.R. PKA phosphorylation dissociates FKBP12.6 from the calcium release channel (ryanodine receptor): Defective regulation in failing hearts. Cell 2000, 101, 365–376. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, Y.; Fu, Q.; Xu, B.; Zhang, Y.; Kim, S.; Tan, R.; Barbagallo, F.; West, T.; Anderson, E.; et al. Inhibiting insulin-mediated β 2-adrenergic receptor activation prevents diabetes-associated cardiac dysfunction. Circulation 2017, 135, 73–88. [Google Scholar] [CrossRef]
- Barbagallo, F.; Xu, B.; Reddy, G.R.; West, T.; Wang, Q.; Fu, Q.; Li, M.; Shi, Q.; Ginsburg, K.S.; Ferrier, W.; et al. Genetically Encoded Biosensors Reveal PKA Hyperphosphorylation on the Myofilaments in Rabbit Heart Failure. Circ. Res. 2016, 119, 931–943. [Google Scholar] [CrossRef]
- Butler, L.; Cros, C.; Oldman, K.L.; Harmer, A.R.; Pointon, A.; Pollard, C.E.; Abi-Gerges, N. Enhanced characterization of contractility in cardiomyocytes during early drug safety assessment. Toxicol. Sci. 2015, 145, 396–406. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Wold, L.E. Measurement of cardiac mechanical function in isolated ventricular myocytes from rats and mice by computerized video-based imaging. Biol. Proced. Online 2001, 3, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Delbridge, L.M.D.; Roos, K.P. Optical methods to evaluate the contractile function of unloaded isolated cardiac myocytes. J. Mol. Cell. Cardiol. 1997, 29, 11–25. [Google Scholar] [CrossRef]
- Fagundes, D.L.; Goulart, J.T. ContHeart: Software for monitoring isolated cardiomyocyte shortening. SoftwareX 2020, 12, 100547. [Google Scholar] [CrossRef]
- Bazan, C.; Barba, D.T.; Blomgren, P.; Paolini, P. Image processing techniques for assessing contractility in isolated adult cardiac myocytes. Int. J. Biomed. Imaging 2009, 2009, 352954. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, T.; Kunihiro, T.; Ando, T.; Kobayashi, S.; Matsui, E.; Yada, H.; Kanda, Y.; Kurokawa, J.; Furukawa, T. Image-based evaluation of contraction-relaxation kinetics of human-induced pluripotent stem cell-derived cardiomyocytes: Correlation and complementarity with extracellular electrophysiology. J. Mol. Cell. Cardiol. 2014, 77, 178–191. [Google Scholar] [CrossRef] [PubMed]
- Toepfer, C.N.; Sharma, A.; Cicconet, M.; Garfinkel, A.C.; Mücke, M.; Neyazi, M.; Willcox, J.A.L.; Agarwal, R.; Schmid, M.; Rao, J.; et al. SarcTrack: An adaptable software tool for efficient large-scale analysis of sarcomere function in hiPSC-cardiomyocytes. Circ. Res. 2019, 124, 1172–1183. [Google Scholar] [CrossRef]
- Pasqualin, C.; Gannier, F.; Yu, A.; Malécot, C.O.; Bredeloux, P.; Maupoil, V. SarcOptiM for ImageJ: High-frequency online sarcomere length computing on stimulated cardiomyocytes. Am. J. Physiol.-Cell Physiol. 2016, 311, C277–C283. [Google Scholar] [CrossRef]
- Muto, A.; Ohkura, M.; Abe, G.; Nakai, J.; Kawakami, K. Real-time visualization of neuronal activity during perception. Curr. Biol. 2013, 23, 307–311. [Google Scholar] [CrossRef]
- Fassina, L.; Rozzi, G.; Rossi, S.; Scacchi, S.; Galetti, M.; Lo Muzio, F.P.; Del Bianco, F.; Colli Franzone, P.; Petrilli, G.; Faggian, G.; et al. Cardiac kinematic parameters computed from video of in situ beating heart. Sci. Rep. 2017, 7, 46143. [Google Scholar] [CrossRef]
- Fassina, L.; di Grazia, A.; Naro, F.; Monaco, L.; de Angelis, M.G.C.; Magenes, G. Video evaluation of the kinematics and dynamics of the beating cardiac syncytium: An alternative to the langendorff method. Int. J. Artif. Organs 2011, 34, 546–558. [Google Scholar] [CrossRef] [PubMed]
- Lo Muzio, F.P.; Rozzi, G.; Rossi, S.; Gerbolés, A.G.; Fassina, L.; Pelà, G.; Luciani, G.B.; Miragoli, M. In-situ optical assessment of rat epicardial kinematic parameters reveals frequency-dependent mechanic heterogeneity related to gender. Prog. Biophys. Mol. Biol. 2019, 154, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Rozzi, G.; Lo Muzio, F.P.; Sandrini, C.; Rossi, S.; Fassina, L.; Faggian, G.; Miragoli, M.; Luciani, G.B. Real-time video kinematic evaluation of the in situ beating right ventricle after pulmonary valve replacement in patients with tetralogy of Fallot: A pilot study. Interact. Cardiovasc. Thorac. Surg. 2019, 29, 625–631. [Google Scholar] [CrossRef] [PubMed]
- Fassina, L.; Magenes, G.; Gimmelli, R.; Naro, F. Modulation of the cardiomyocyte contraction inside a hydrostatic pressure bioreactor: In vitro verification of the frank-starling law. Biomed. Res. Int. 2015, 2015, 542105. [Google Scholar] [CrossRef] [PubMed]
- West, T.M.; Wang, Q.; Deng, B.; Zhang, Y.; Barbagallo, F.; Reddy, G.R.; Chen, D.; Phan, K.S.; Xu, B.; Isidori, A.; et al. Phosphodiesterase 5 Associates With β2 Adrenergic Receptor to Modulate Cardiac Function in Type 2 Diabetic Hearts. J. Am. Heart Assoc. 2019, 8, e012273. [Google Scholar] [CrossRef] [PubMed]
- Raake, P.W.; Vinge, L.E.; Gao, E.; Boucher, M.; Rengo, G.; Chen, X.; Degeorge, B.R.; Matkovich, S.; Houser, S.R.; Most, P.; et al. G protein-coupled receptor kinase 2 ablation in cardiac myocytes before or after myocardial infarction prevents heart failure. Circ. Res. 2008, 103, 413–422. [Google Scholar] [CrossRef] [PubMed]
- Bers, D.M. Cardiac excitation-contraction coupling. Nature 2002, 415, 198–205. [Google Scholar] [CrossRef]
- Gorski, P.A.; Kho, C.; Oh, J.G. Measuring cardiomyocyte contractility and calcium handling in vitro. Methods Mol. Biol. 2018, 1816, 93–104. [Google Scholar] [CrossRef]
- Sala, L.; Van Meer, B.J.; Tertoolen, L.G.J.; Bakkers, J.; Bellin, M.; Davis, R.P.; Denning, C.; Dieben, M.A.E.; Eschenhagen, T.; Giacomelli, E.; et al. Musclemotion: A versatile open software tool to quantify cardiomyocyte and cardiac muscle contraction in vitro and in vivo. Circ. Res. 2018, 122, e5–e16. [Google Scholar] [CrossRef]
- Di Pasquale, E.; Lodola, F.; Miragoli, M.; Denegri, M.; Avelino-Cruz, J.E.; Buonocore, M.; Nakahama, H.; Portararo, P.; Bloise, R.; Napolitano, C.; et al. CaMKII inhibition rectifies arrhythmic phenotype in a patient-specific model of catecholaminergic polymorphic ventricular tachycardia. Cell Death Dis. 2013, 4, e843. [Google Scholar] [CrossRef]
- Greensmith, D.J. Ca analysis: An excel based program for the analysis of intracellular calcium transients including multiple, simultaneous regression analysis. Comput. Methods Programs Biomed. 2014, 113, 241–250. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Psaras, Y.; Margara, F.; Cicconet, M.; Sparrow, A.J.; Repetti, G.G.; Schmid, M.; Steeples, V.; Wilcox, J.A.L.; Bueno-Orovio, A.; Redwood, C.S.; et al. CalTrack: High-Throughput Automated Calcium Transient Analysis in Cardiomyocytes. Circ. Res. 2021, 129, 326–341. [Google Scholar] [CrossRef] [PubMed]
- Gesmundo, I.; Miragoli, M.; Carullo, P.; Trovato, L.; Larcher, V.; Di Pasquale, E.; Brancaccio, M.; Mazzola, M.; Villanova, T.; Sorge, M.; et al. Growth hormone-releasing hormone attenuates cardiac hypertrophy and improves heart function in pressure overload-induced heart failure. Proc. Natl. Acad. Sci. USA 2017, 114, 12033–12038. [Google Scholar] [CrossRef] [PubMed]
- Meraviglia, V.; Wen, J.; Piacentini, L.; Campostrini, G.; Wang, C.; Florio, M.C.; Azzimato, V.; Fassina, L.; Langes, M.; Wong, J.; et al. Higher cardiogenic potential of IPSCs derived from cardiac versus skin stromal cells. Front. Biosci.-Landmark 2016, 21, 719–743. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fassina, L.; Assenza, M.R.; Miragoli, M.; Isidori, A.M.; Naro, F.; Barbagallo, F. Cell Shortening and Calcium Homeostasis Analysis in Adult Cardiomyocytes via a New Software Tool. Biomedicines 2022, 10, 640. https://doi.org/10.3390/biomedicines10030640
Fassina L, Assenza MR, Miragoli M, Isidori AM, Naro F, Barbagallo F. Cell Shortening and Calcium Homeostasis Analysis in Adult Cardiomyocytes via a New Software Tool. Biomedicines. 2022; 10(3):640. https://doi.org/10.3390/biomedicines10030640
Chicago/Turabian StyleFassina, Lorenzo, Maria Rita Assenza, Michele Miragoli, Andrea M. Isidori, Fabio Naro, and Federica Barbagallo. 2022. "Cell Shortening and Calcium Homeostasis Analysis in Adult Cardiomyocytes via a New Software Tool" Biomedicines 10, no. 3: 640. https://doi.org/10.3390/biomedicines10030640
APA StyleFassina, L., Assenza, M. R., Miragoli, M., Isidori, A. M., Naro, F., & Barbagallo, F. (2022). Cell Shortening and Calcium Homeostasis Analysis in Adult Cardiomyocytes via a New Software Tool. Biomedicines, 10(3), 640. https://doi.org/10.3390/biomedicines10030640