
Cre/LoxP Genetic Recombination Sustains Cartilage Anabolic Factor Expression in Hyaluronan Encapsulated MSCs Alleviates Intervertebral Disc Degeneration

 ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Construction of Recombinant Baculovirus and Titer Determination
2.2. Cell Culture, Baculoviral Transduction, Western Blot, and Luminescence Examination
2.3. Analysis of Cell Viability, Proliferation, and qRT-PCR
2.4. Rat Coccygeal Disc Degeneration Model, Hyaluronan Encapsulation, BMSCs Transplantation, and Radiographic Imaging
2.5. Histological and Immunohistochemical Staining
2.6. Statistical Analysis
3. Results
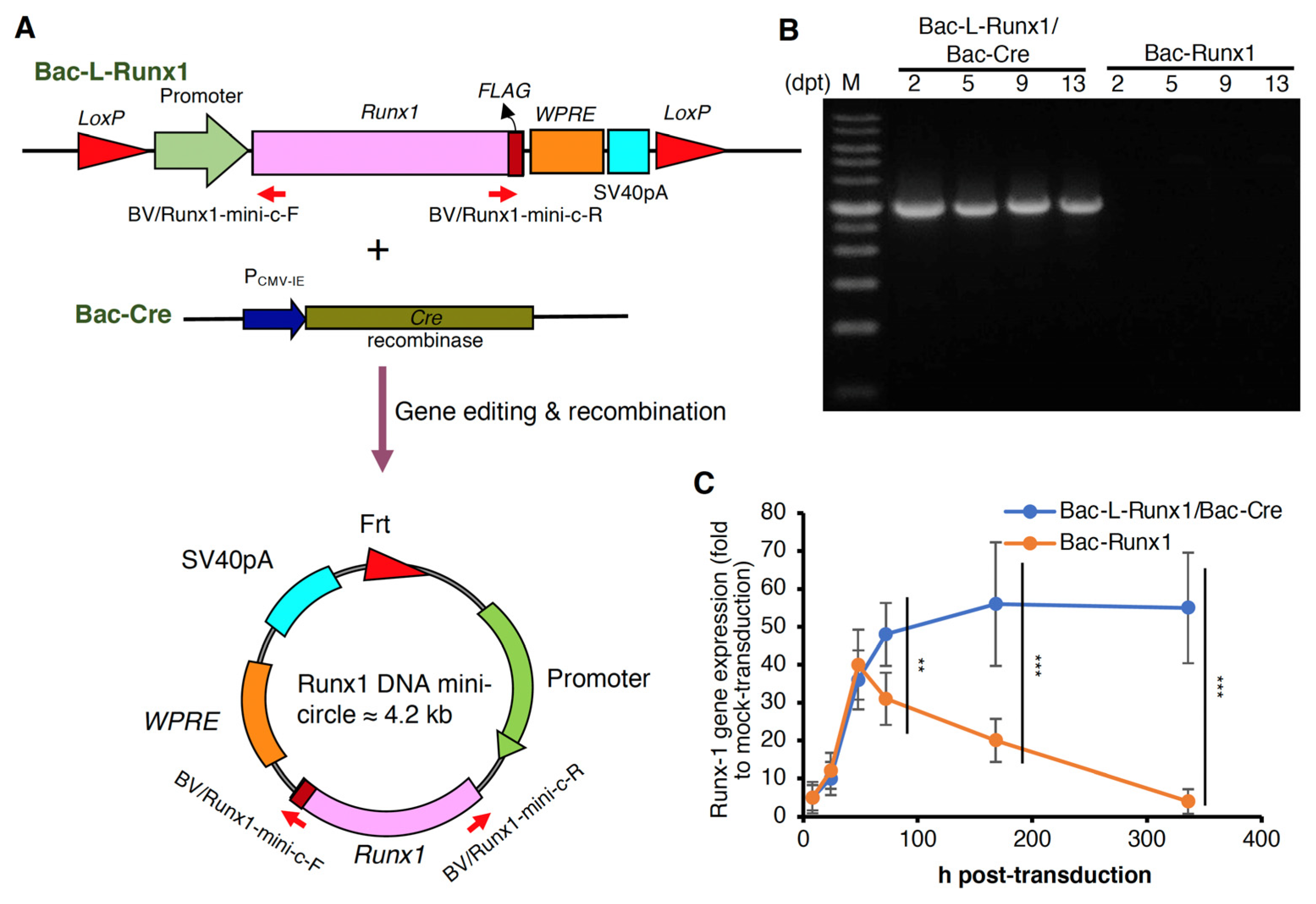
3.1. Sustained Runx1 Expression Mediated by Cre/LoxP Recombinant Baculoviral Vector
3.2. Cre/LoxP Recombinant Baculoviral Vector Elicited Low Cytotoxicity and Safety Concerns
3.3. Chondrogenesis of BMSCs Induced by runx1 Expressed Baculoviral Vector Transduction
3.4. Disc Height Shrinkage Ameliorated by runx1 Overexpressed BMSCs Transplantation in a Disc Degeneration Model in the Rat
3.5. Cre/LoxP Recombinant Baculoviral Vector-Mediated runx1 Sustained Expression Ameliorated Disc Degeneration Progress and ECM Loss
3.6. Transplantation of Cre/LoxP Recombinant Baculoviral Vector Modified BMSCs Elicited Sparse Adaptive Immune Responses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- James, S.L.; Abate, D.; Abate, K.H.; Abay, S.M.; Abbafati, C.; Abbasi, N.; Abbastabar, H.; Abd-Allah, F.; Abdela, J.; Abdelalim, A. Global, regional, and national incidence, prevalence, and years lived with disability for 354 diseases and injuries for 195 countries and territories, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1789–1858. [Google Scholar] [CrossRef]
- Martin, B.I.; Deyo, R.A.; Mirza, S.K.; Turner, J.A.; Comstock, B.A.; Hollingworth, W.; Sullivan, S.D. Expenditures and health status among adults with back and neck problems. JAMA 2008, 299, 656–664. [Google Scholar] [CrossRef] [PubMed]
- Vos, T.; Flaxman, A.D.; Naghavi, M.; Lozano, R.; Michaud, C.; Ezzati, M.; Shibuya, K.; Salomon, J.A.; Abdalla, S.; Aboyans, V.; et al. Years lived with disability (YLDs) for 1160 sequelae of 289 diseases and injuries 1990–2010: A systematic analysis for the Global Burden of Disease Study 2010. Lancet 2012, 380, 2163–2196. [Google Scholar] [CrossRef]
- Cheung, K.M.; Karppinen, J.; Chan, D.; Ho, D.W.; Song, Y.Q.; Sham, P.; Cheah, K.S.; Leong, J.C.; Luk, K.D. Prevalence and pattern of lumbar magnetic resonance imaging changes in a population study of one thousand forty-three individuals. Spine 2009, 34, 934–940. [Google Scholar] [CrossRef] [PubMed]
- Takatalo, J.; Karppinen, J.; Niinimaki, J.; Taimela, S.; Nayha, S.; Mutanen, P.; Sequeiros, R.B.; Kyllonen, E.; Tervonen, O. Does lumbar disc degeneration on magnetic resonance imaging associate with low back symptom severity in young Finnish adults? Spine 2011, 36, 2180–2189. [Google Scholar] [CrossRef]
- Panjabi, M.M.; Oxland, T.R.; Yamamoto, I.; Crisco, J.J. Mechanical behavior of the human lumbar and lumbosacral spine as shown by three-dimensional load-displacement curves. J. Bone Jt. Surg. 1994, 76, 413–424. [Google Scholar] [CrossRef]
- Pattappa, G.; Li, Z.; Peroglio, M.; Wismer, N.; Alini, M.; Grad, S. Diversity of intervertebral disc cells: Phenotype and function. J. Anat. 2012, 221, 480–496. [Google Scholar] [CrossRef]
- Miele, V.J.; Panjabi, M.M.; Benzel, E.C. Anatomy and biomechanics of the spinal column and cord. Handb. Clin. Neurol. 2012, 109, 31–43. [Google Scholar] [CrossRef]
- Johnson, K.; Zhu, S.; Tremblay, M.S.; Payette, J.N.; Wang, J.; Bouchez, L.C.; Meeusen, S.; Althage, A.; Cho, C.Y.; Wu, X.; et al. A stem cell-based approach to cartilage repair. Science 2012, 336, 717–721. [Google Scholar] [CrossRef]
- Yano, F.; Hojo, H.; Ohba, S.; Fukai, A.; Hosaka, Y.; Ikeda, T.; Saito, T.; Hirata, M.; Chikuda, H.; Takato, T.; et al. A novel disease-modifying osteoarthritis drug candidate targeting Runx1. Ann. Rheum. Dis. 2013, 72, 748–753. [Google Scholar] [CrossRef]
- Wang, Y.; Belflower, R.M.; Dong, Y.F.; Schwarz, E.M.; O’Keefe, R.J.; Drissi, H. Runx1/AML1/Cbfa2 mediates onset of mesenchymal cell differentiation toward chondrogenesis. J. Bone Miner. Res. 2005, 20, 1624–1636. [Google Scholar] [CrossRef]
- Wotton, S.; Terry, A.; Kilbey, A.; Jenkins, A.; Herzyk, P.; Cameron, E.; Neil, J.C. Gene array analysis reveals a common Runx transcriptional programme controlling cell adhesion and survival. Oncogene 2008, 27, 5856–5866. [Google Scholar] [CrossRef]
- Lin, C.Y.; Crowley, S.T.; Uchida, S.; Komaki, Y.; Kataoka, K.; Itaka, K. Treatment of Intervertebral Disk Disease by the Administration of mRNA Encoding a Cartilage-Anabolic Transcription Factor. Mol. Ther. Nucleic Acids 2019, 16, 162–171. [Google Scholar] [CrossRef]
- Belzhelarskaia, S.N. Baculovirus expression systems for recombinant protein production in insect and mammalian cells. Mol. Biol. 2011, 45, 142–159. [Google Scholar]
- Kost, T.A.; Condreay, J.P.; Jarvis, D.L. Baculovirus as versatile vectors for protein expression in insect and mammalian cells. Nat. Biotechnol. 2005, 23, 567–575. [Google Scholar] [CrossRef]
- Goehring, A.; Lee, C.H.; Wang, K.H.; Michel, J.C.; Claxton, D.P.; Baconguis, I.; Althoff, T.; Fischer, S.; Garcia, K.C.; Gouaux, E. Screening and large-scale expression of membrane proteins in mammalian cells for structural studies. Nat. Protoc. 2014, 9, 2574–2585. [Google Scholar] [CrossRef]
- Thimiri Govinda Raj, D.B.; Khan, N.A.; Venkatachalam, S.; Arumugam, S. BacMam System for Rapid Recombinant Protein Expression in Mammalian Cells. Methods Mol. Biol. 2020, 2125, 205–208. [Google Scholar] [CrossRef]
- Chen, C.Y.; Lin, C.Y.; Chen, G.Y.; Hu, Y.C. Baculovirus as a gene delivery vector: Recent understandings of molecular alterations in transduced cells and latest applications. Biotechnol. Adv. 2011, 29, 618–631. [Google Scholar] [CrossRef]
- Lin, C.Y.; Wang, Y.H.; Li, K.C.; Sung, L.Y.; Yeh, C.L.; Lin, K.J.; Yen, T.C.; Chang, Y.H.; Hu, Y.C. Healing of massive segmental femoral bone defects in minipigs by allogenic ASCs engineered with FLPo/Frt-based baculovirus vectors. Biomaterials 2015, 50, 98–106. [Google Scholar] [CrossRef]
- Li, K.C.; Lo, S.C.; Sung, L.Y.; Liao, Y.H.; Chang, Y.H.; Hu, Y.C. Improved calvarial bone repair by hASCs engineered with Cre/loxP-based baculovirus conferring prolonged BMP-2 and MiR-148b co-expression. J. Tissue Eng. Regen. Med. 2017, 11, 3068–3077. [Google Scholar] [CrossRef]
- Aini, H.; Itaka, K.; Fujisawa, A.; Uchida, H.; Uchida, S.; Fukushima, S.; Kataoka, K.; Saito, T.; Chung, U.I.; Ohba, S. Messenger RNA delivery of a cartilage-anabolic transcription factor as a disease-modifying strategy for osteoarthritis treatment. Sci. Rep. 2016, 6, 18743. [Google Scholar] [CrossRef] [PubMed]
- O’Reilly, D.; Miller, L.; Luckow, V. Baculovirus Expression Vectors: A Laboratory Manual; W.H. Freeman and Co.: New York, NY, USA, 1992; pp. 109–215. [Google Scholar]
- Han, S.; Wang, B.; Li, X.; Xiao, Z.; Han, J.; Zhao, Y.; Fang, Y.; Yin, Y.; Chen, B.; Dai, J. Bone marrow-derived mesenchymal stem cells in three-dimensional culture promote neuronal regeneration by neurotrophic protection and immunomodulation. J. Biomed. Mater. Res. A 2016, 104, 1759–1769. [Google Scholar] [CrossRef]
- Liao, J.C. Cell Therapy Using Bone Marrow-Derived Stem Cell Overexpressing BMP-7 for Degenerative Discs in a Rat Tail Disc Model. Int. J. Mol. Sci. 2016, 17, 147. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.Y.; Chang, Y.H.; Lin, K.J.; Yen, T.C.; Tai, C.L.; Chen, C.Y.; Lo, W.H.; Hsiao, I.T.; Hu, Y.C. The healing of critical-sized femoral segmental bone defects in rabbits using baculovirus-engineered mesenchymal stem cells. Biomaterials 2010, 31, 3222–3230. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.Y.; Chang, Y.H.; Kao, C.Y.; Lu, C.H.; Sung, L.Y.; Yen, T.C.; Lin, K.J.; Hu, Y.C. Augmented healing of critical-size calvarial defects by baculovirus-engineered MSCs that persistently express growth factors. Biomaterials 2012, 33, 3682–3692. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.; Yin, Y.; Xia, Q.J.; Lin, N.; Wang, Y.C.; Liu, J.; Wang, H.P.; Lim, A.; Wang, T.H. Bone Marrow Stromal Cells Promote Neuronal Restoration in Rats with Traumatic Brain Injury: Involvement of GDNF Regulating BAD and BAX Signaling. Cell. Physiol. Biochem. 2016, 38, 748–762. [Google Scholar] [CrossRef] [PubMed]
- Lo, W.-H.; Hwang, S.-M.; Chuang, C.-K.; Chen, C.-Y.; Hu, Y.-C. Development of a hybrid baculoviral vector for sustained transgene expression. Mol. Ther. 2009, 17, 658–666. [Google Scholar] [CrossRef]
- Bian, L.; Guvendiren, M.; Mauck, R.L.; Burdick, J.A. Hydrogels that mimic developmentally relevant matrix and N-cadherin interactions enhance MSC chondrogenesis. Proc. Natl. Acad. Sci. USA 2013, 110, 10117–10122. [Google Scholar] [CrossRef]
- Inoue, H.; Montgomery, S.R.; Aghdasi, B.; Kaner, T.; Tan, Y.; Tian, H.; Terrell, R.; Wang, J.C.; Daubs, M.D. The effect of bone morphogenetic protein-2 injection at different time points on intervertebral disk degeneration in a rat tail model. J. Spinal Disord. Tech. 2015, 28, E35–E44. [Google Scholar] [CrossRef]
- Sakai, D.; Andersson, G.B. Stem cell therapy for intervertebral disc regeneration: Obstacles and solutions. Nat. Reviews. Rheumatol. 2015, 11, 243–256. [Google Scholar] [CrossRef]
- Lin, C.Y.; Lin, K.J.; Li, K.C.; Sung, L.Y.; Hsueh, S.; Lu, C.H.; Chen, G.Y.; Chen, C.L.; Huang, S.F.; Yen, T.C.; et al. Immune responses during healing of massive segmental femoral bone defects mediated by hybrid baculovirus-engineered ASCs. Biomaterials 2012, 33, 7422–7434. [Google Scholar] [CrossRef] [PubMed]
- Murphy, K.; Weaver, C. Janeway’s Immunobiology; Garland Science: New York, NY, USA, 2016. [Google Scholar]
- Kong, L.; Cao, J.; Wang, L.; Shen, Y. Prevalence of adjacent segment disease following cervical spine surgery: A PRISMA-compliant systematic review and meta-analysis. Medicine 2016, 95, e4171. [Google Scholar] [CrossRef]
- Pan, A.; Hai, Y.; Yang, J.; Zhou, L.; Chen, X.; Guo, H. Adjacent segment degeneration after lumbar spinal fusion compared with motion-preservation procedures: A meta-analysis. Eur. Spine J. 2016, 25, 1522–1532. [Google Scholar] [CrossRef]
- Ren, C.; Song, Y.; Liu, L.; Xue, Y. Adjacent segment degeneration and disease after lumbar fusion compared with motion-preserving procedures: A meta-analysis. Eur. J. Orthop. Surg. Traumatol. 2014, 24, S245–S253. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.P.; Chen, H.L.; Cheng, H.B. Prevalence of adjacent segment degeneration after spine surgery: A systematic review and meta-analysis. Spine 2013, 38, 597–608. [Google Scholar] [CrossRef] [PubMed]
- Mochida, J.; Nishimura, K.; Nomura, T.; Toh, E.; Chiba, M. The importance of preserving disc structure in surgical approaches to lumbar disc herniation. Spine 1996, 21, 1556–1563. [Google Scholar] [CrossRef]
- Fontana, G.; See, E.; Pandit, A. Current trends in biologics delivery to restore intervertebral disc anabolism. Adv. Drug Deliv. Rev. 2015, 84, 146–158. [Google Scholar] [CrossRef]
- Urban, J.P.; Roberts, S. Degeneration of the intervertebral disc. Arthritis Res. 2003, 5, 120–130. [Google Scholar] [CrossRef]
- Chan, D.; Song, Y.; Sham, P.; Cheung, K.M. Genetics of disc degeneration. Eur. Spine J. 2006, 15, S317–S325. [Google Scholar] [CrossRef]
- Blanco, F.J.; Ruiz-Romero, C. New targets for disease modifying osteoarthritis drugs: Chondrogenesis and Runx1. Ann. Rheum. Dis. 2013, 72, 631–634. [Google Scholar] [CrossRef]
- Vergroesen, P.P.; Kingma, I.; Emanuel, K.S.; Hoogendoorn, R.J.; Welting, T.J.; van Royen, B.J.; van Dieen, J.H.; Smit, T.H. Mechanics and biology in intervertebral disc degeneration: A vicious circle. Osteoarthr. Cartil. 2015, 23, 1057–1070. [Google Scholar] [CrossRef] [PubMed]
- Norcross, J.P.; Lester, G.E.; Weinhold, P.; Dahners, L.E. An in vivo model of degenerative disc disease. J. Orthop. Res. 2003, 21, 183–188. [Google Scholar] [CrossRef]
- Gurung, S.; Khan, F.; Gunassekaran, G.R.; Yoo, J.D.; Poongkavithai Vadevoo, S.M.; Permpoon, U.; Kim, S.H.; Kim, H.J.; Kim, I.S.; Han, H.; et al. Phage display-identified PD-L1-binding peptides reinvigorate T-cell activity and inhibit tumor progression. Biomaterials 2020, 247, 119984. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Qin, Z.; Zhao, J.; He, Y.; Ren, E.; Zhu, Y.; Liu, G.; Mao, C.; Zheng, L. Cartilage-targeting and dual MMP-13/pH responsive theranostic nanoprobes for osteoarthritis imaging and precision therapy. Biomaterials 2019, 225, 119520. [Google Scholar] [CrossRef] [PubMed]
- Ho-Shui-Ling, A.; Bolander, J.; Rustom, L.E.; Johnson, A.W.; Luyten, F.P.; Picart, C. Bone regeneration strategies: Engineered scaffolds, bioactive molecules and stem cells current stage and future perspectives. Biomaterials 2018, 180, 143–162. [Google Scholar] [CrossRef]
- Chen, C.Y.; Wu, H.H.; Chen, C.P.; Chern, S.R.; Hwang, S.M.; Huang, S.F.; Lo, W.H.; Chen, G.Y.; Hu, Y.C. Biosafety assessment of human mesenchymal stem cells engineered by hybrid baculovirus vectors. Mol. Pharm. 2011, 8, 1505–1514. [Google Scholar] [CrossRef]
- Luo, W.Y.; Lin, S.Y.; Lo, K.W.; Lu, C.H.; Hung, C.L.; Chen, C.Y.; Chang, C.C.; Hu, Y.C. Adaptive immune responses elicited by baculovirus and impacts on subsequent transgene expression in vivo. J. Virol. 2013, 87, 4965–4973. [Google Scholar] [CrossRef]
- Li, K.C.; Chang, Y.H.; Lin, C.Y.; Hwang, S.M.; Wang, T.H.; Hu, Y.C. Preclinical Safety Evaluation of ASCs Engineered by FLPo/Frt-Based Hybrid Baculovirus: In Vitro and Large Animal Studies. Tissue Eng. Part. A 2015, 21, 1471–1482. [Google Scholar] [CrossRef]
- Shegarfi, H.; Reikeras, O. Review article: Bone transplantation and immune response. J. Orthop. Surg. 2009, 17, 206–211. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chan, L.-Y.; Chang, C.-C.; Lai, P.-L.; Maeda, T.; Hsu, H.-C.; Lin, C.-Y.; Kuo, S.-J. Cre/LoxP Genetic Recombination Sustains Cartilage Anabolic Factor Expression in Hyaluronan Encapsulated MSCs Alleviates Intervertebral Disc Degeneration. Biomedicines 2022, 10, 555. https://doi.org/10.3390/biomedicines10030555
Chan L-Y, Chang C-C, Lai P-L, Maeda T, Hsu H-C, Lin C-Y, Kuo S-J. Cre/LoxP Genetic Recombination Sustains Cartilage Anabolic Factor Expression in Hyaluronan Encapsulated MSCs Alleviates Intervertebral Disc Degeneration. Biomedicines. 2022; 10(3):555. https://doi.org/10.3390/biomedicines10030555
Chicago/Turabian StyleChan, Long-Yi, Cheng-Chung Chang, Po-Liang Lai, Tomoji Maeda, Horng-Chaung Hsu, Chin-Yu Lin, and Shu-Jui Kuo. 2022. "Cre/LoxP Genetic Recombination Sustains Cartilage Anabolic Factor Expression in Hyaluronan Encapsulated MSCs Alleviates Intervertebral Disc Degeneration" Biomedicines 10, no. 3: 555. https://doi.org/10.3390/biomedicines10030555
APA StyleChan, L.-Y., Chang, C.-C., Lai, P.-L., Maeda, T., Hsu, H.-C., Lin, C.-Y., & Kuo, S.-J. (2022). Cre/LoxP Genetic Recombination Sustains Cartilage Anabolic Factor Expression in Hyaluronan Encapsulated MSCs Alleviates Intervertebral Disc Degeneration. Biomedicines, 10(3), 555. https://doi.org/10.3390/biomedicines10030555