
Shock Waves Enhance Expression of Glycosphingolipid Tumor Antigen on Renal Cell Carcinoma: Dynamics of Physically Unmasking Hidden Intracellular Markers Independent of Gene-Signaling Pathways



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Line
2.2. Shock Wave Generator
2.3. Shock Wave Treatment
2.4. Cell Viability
2.5. Flowcytometry
2.6. Scanning Electron Microscopy
2.7. Immunoelectron Microscopy
2.8. Statistical Analysis
3. Results
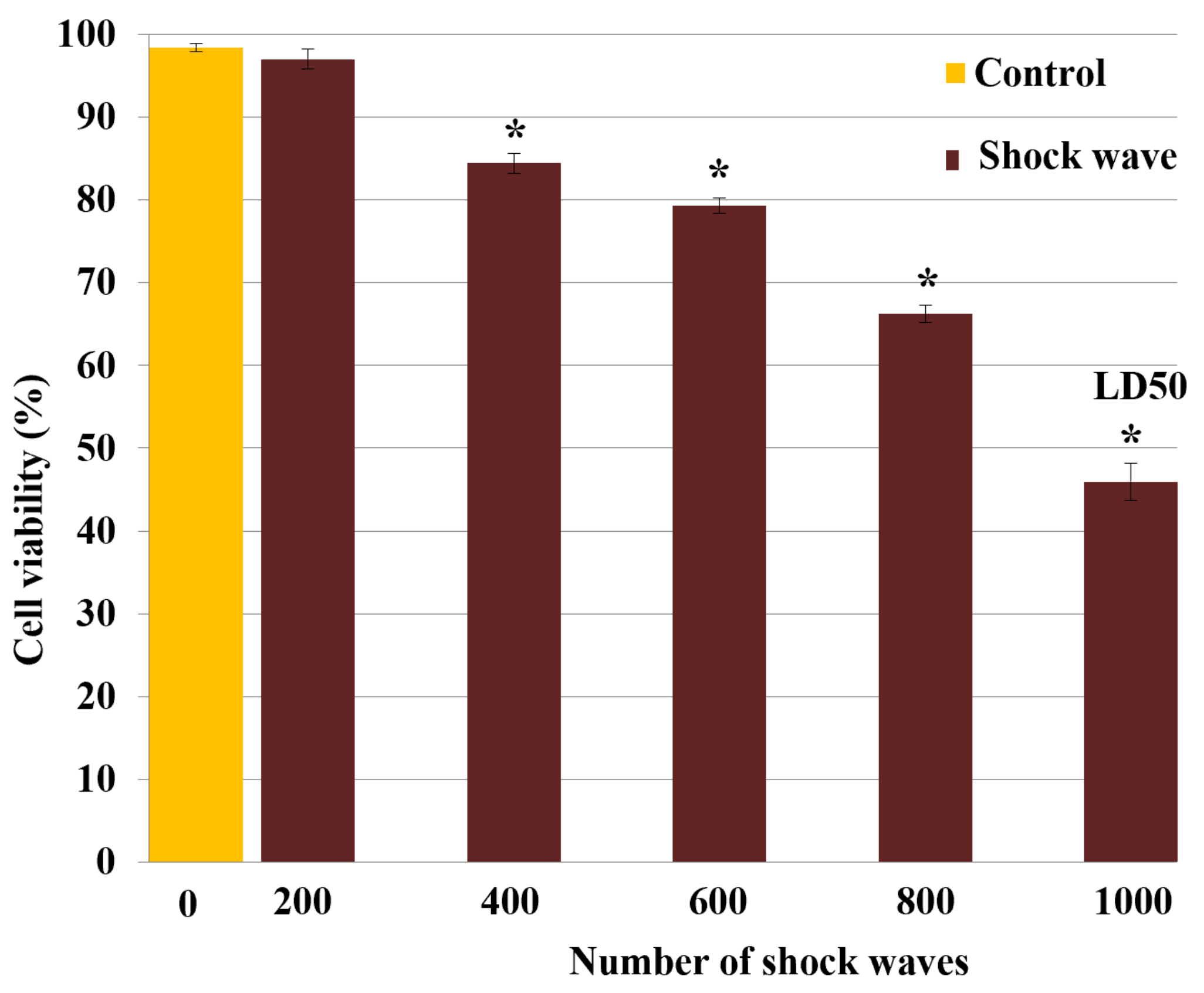
3.1. Effect of Shock Waves on Cell Viability
3.2. Effect of Shock Waves on Particle Displacement and Temperature
3.3. Effect of Shock Waves on Membrane Expression of MSGG Antigen
3.3.1. Flowcytometric Measurement
3.3.2. Immunoelectron Microscopic Analysis
3.4. Morphological Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, Y.; Zhang, Z. The history and advances in cancer immunotherapy: Understanding the characteristics of tumor-infiltrating immune cells and their therapeutic implications. Cell. Mol. Immunol. 2020, 17, 807–821. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.; Chehrazi-Raffle, A.; Reddi, S.; Salgia, R. Development of PD-1 and PD-L1 inhibitors as a form of cancer immunotherapy: A comprehensive review of registration trials and future considerations. J. Immunother. Cancer 2018, 6, 8. [Google Scholar] [CrossRef]
- Waldman, A.D.; Fritz, J.M.; Lenardo, M.J. A guide to cancer immunotherapy: From T cell basic science to clinical practice. Nat. Rev. Immunol. 2020, 20, 651–668. [Google Scholar] [CrossRef] [PubMed]
- Hollingsworth, R.E.; Jansen, K. Turning the corner on therapeutic cancer vaccines. NPJ Vaccines 2019, 4, 7. [Google Scholar] [CrossRef] [PubMed]
- Fares, C.M.; Van Allen, E.M.; Drake, C.G.; Allison, J.P.; Hu-Lieskovan, S. Mechanisms of Resistance to Immune Checkpoint Blockade: Why Does Checkpoint Inhibitor Immunotherapy Not Work for All Patients? Developmental Immunotherapy and Tumor Immunobiology. Am. Soc. Clin. Oncol. Educ. Book 2019, 39, 147–164. [Google Scholar] [CrossRef] [PubMed]
- Bai, R.; Chen, N.; Li, L.; Du, N.; Bai, L.; Lv, Z.; Tian, H.; Cui, J. Mechanisms of cancer resistance to immunotherapy. Front. Oncol. 2020, 10, 1290. [Google Scholar] [CrossRef]
- Amato, R.J. Vaccine therapy for renal cell carcinoma. Rev. Urol. 2003, 5, 65–71. [Google Scholar]
- Hakomori, S. Cancer-associated glycosphingolipid antigens: Their structure, organization, and function. Cells Tissues Organs 1998, 161, 79–90. [Google Scholar] [CrossRef]
- Satoh, M.; Moosavi Nejad, F.; Nakano, O.; Ito, A.; Kawamura, S.; Oyama, C.; Saito, S.; Orikasa, S. Four new human renal cell carcinoma cell lines expressing globe-series gangliosides. Tohoku J. Exp. Med. 1999, 189, 95–105. [Google Scholar] [CrossRef]
- Yu, J.; Hung, J.T.; Wang, S.H.; Cheng, J.Y.; Yu, A.L. Targeting glycosphingolipids for cancer immunotherapy. FEBS Lett. 2020, 594, 3602–3618. [Google Scholar] [CrossRef]
- Ando, T.; Ito, H.; Arioka, Y.; Ogiso, H.; Seishima, M. Combination therapy with α-galactosylceramide and a toll-like receptor agonist exerts an augmented suppressive effect on lung tumor metastasis in a mouse model. Oncol. Rep. 2015, 33, 826–832. [Google Scholar] [CrossRef] [PubMed]
- Tong, W.; Maira, M.; Roychoudhury, R.; Galan, A.; Brahimi, F.; Gilbert, M.; Cunningham, A.M.; Josephy, S.; Pirvulescu, I.; Moffett, S.; et al. Vaccination with tumor-ganglioside glycomimetics activates a selective immunity that affords cancer therapy. Cell Chem. Biol. 2019, 26, 1013–1026.e4. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Deng, W.; Li, N.; Neri, S.; Sharma, A.; Jiang, W.; Lin, S.H. Combining immunotherapy and radiotherapy for cancer treatment: Current challenges and future directions. Front. Pharmacol. 2018, 9, 185. [Google Scholar] [CrossRef] [PubMed]
- Moosavi-Nejad, S.F.; Hosseini, S.H.R.; Satoh, M.; Takayama, K. Shock wave induced cytoskeletal and morphological deformations in a human renal carcinoma cell line. Cancer Sci. 2006, 97, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Xiaoyu, J.; Yu, X.; Cui, Z.; Zhao, M. The clinical and immunological features of the post-extracorporeal shock wave lithotripsy anti-glomerular basement membrane disease. Ren. Fail. 2021, 43, 149–155. [Google Scholar] [CrossRef]
- Banner, B.; Ziesmer, D.; Collins, L.A. Proliferative glomerulopathy following extracorporeal shock wave lithotripsy in the pig. J. Unrol. 1991, 146, 1425–1428. [Google Scholar] [CrossRef]
- Van den Bijgaart, R.J.; Eikelenboom, D.C.; Hoogenboom, M.; Fütterer, J.J.; den Brok, M.H.; Adema, G.J. Thermal and mechanical high-intensity focused ultrasound: Perspectives on tumor ablation, immune effects and combination strategies. Cancer Immunol. Immunother. 2017, 66, 247–258. [Google Scholar] [CrossRef]
- Cranfield, A.; Mathavakkannan, S. Goodpasture’s disease following extracorporeal shock wave lithotripsy: A case report & literature review. Clin. Case Rep. 2015, 3, 160–164. [Google Scholar] [CrossRef]
- Oshita, D.; Hosseini, S.H.R.; Okuda, Y.; Miyamoto, Y.; Sakugawa, T.; Katsuki, S.; Akiyama, H. Time-Resolved High-Speed Visualization and Analysis of Underwater Shock Wave Focusing Generated by a Magnetic Pulse Compression Unit. IEEE Trans. Plasma Sci. 2012, 40, 2395–2400. [Google Scholar] [CrossRef]
- Ohtani, H.; Sasano, N. Characterization of Microvasculature in the Stroma of Human Colorectal Carcinoma: An Immunoelectron Microscopic Study on Factor VIII/von Willebrand Factor. J. Electron. Microsc. 1987, 36, 204–212. [Google Scholar] [CrossRef]
- Gojani, A.B.; Ohtani, K.; Takayama, K.; Hosseini, S.H.R. Shock Hugoniot and equations of states of water, castor oil, and aqueous solutions of sodium chloride, sucrose and gelatin. Shock Waves 2016, 26, 63–68. [Google Scholar] [CrossRef]
- Hosseini, H.; Moosavi-Nejad, S.; Akiyama, H.; Menezes, V. Shock wave interaction with interfaces between materials having different acoustic impedances. Appl. Phys. Lett. 2014, 104, 103701. [Google Scholar] [CrossRef]
- Menezes, V.; Hosseini, H.; Moosavi-Nejad, S.; Irimpan, K.J.; Akiyama, H. Motion of free-surface of shock-compressed water on emergence of rarefaction. Appl. Phys. Lett. 2015, 107, 143701. [Google Scholar] [CrossRef]
- Nejad, S.M.; Hosseini, H.; Akiyama, H.; Tachibana, K. Reparable Cell Sonoporation in Suspension: Theranostic Potential of Microbubble. Theranostics 2016, 6, 446–455. [Google Scholar] [CrossRef]
- Wang, F.S.; Yang, K.D.; Chen, R.F.; Wang, C.J.; Sheen-Chen, S.M. Extracorporeal shock wave promotes growth and differentiation of bone-marrow stromal cells towards osteoprogenitors associated with induction of TGF-beta. J. Bone Jt. Surg. 2002, 84, 457–461. [Google Scholar] [CrossRef]
- Merrill, A.H., Jr. Sphingolipid and Glycosphingolipid Metabolic Pathways in the Era of Sphingolipidomics. Chem. Rev. 2011, 111, 6387–6422. [Google Scholar] [CrossRef]
- Tsaklis, P.V. Presentation of Acoustic Waves Propagation and Their Effects through Human Body Tissues. Hum. Mov. 2010, 11, 58–65. [Google Scholar] [CrossRef]
- Hosano, N.; Hosano, H. Prospects and challenges of physical manipulation of the cell membrane: Comment on “Shock wave-induced permeabilization of mammalian cells” by López-Marín et al. Phys. Life Rev. 2018, 26, 43–46. [Google Scholar] [CrossRef]
- Hosseini, S.; Takayama, K. Implosion of a spherical shock wave reflected from a spherical wall. J. Fluid Mech. 2005, 530, 223–239. [Google Scholar] [CrossRef]
- Hosseini, S.; Takayama, K. Experimental study of Richtmyer-Meshkov instability induced by cylindrical shock waves. Phys. Fluids. 2005, 17, 084101. [Google Scholar] [CrossRef]
- Loske, A.M. Medical and Biomedical Applications of Shock Waves; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Prentice, P.; Cuschieri, A.; Dholakia, K.; Prausnitz, M.; Campbell, P. Membrane disruption by optically controlled microbubble cavitation. Nat. Phys. 2005, 1, 107–110. [Google Scholar] [CrossRef]
- Srivastava, R.C.; Leutloff, D.; Takayama, K.; Grönig, H. (Eds.) Shock Focusing Effect in Medical Science and Sonoluminescence; Springer: Berlin/Heidelberg, Germany, 2003. [Google Scholar]
- White, C.R.; Frangos, J.A. The shear stress of it all: The cell membrane and mechanochemical transduction. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2007, 362, 1459–1467. [Google Scholar] [CrossRef] [PubMed]
- Suhr, D.; Brümmer, F.; Irmer, U.; Hülser, D.F. Disturbance of cellular calcium homeostasis by in vitro application of shock waves. Ultrasound Med. Biol. 1996, 22, 671–679. [Google Scholar] [CrossRef]
- Park, J.; Fan, Z.; Deng, C.X. Effects of shear stress cultivation on cell membrane disruption and intracellular calcium concentration in sonoporation of endothelial cells. J. Biomech. 2011, 44, 164–169. [Google Scholar] [CrossRef]
- Gillespie, P.; Walker, R. Molecular basis of mechanosensory transduction. Nature 2001, 413, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Ruan, R.; Zou, L.; Sun, S.; Liu, J.; Wen, L.; Gao, D.; Ding, W. Cell Blebbing upon Addition of Cryoprotectants: A Self-Protection Mechanism. PLoS ONE 2015, 10, e0125746. [Google Scholar] [CrossRef]
- Martin, C.J.; Gemmell, H.G. A study of ultrasonically induced pulsations of gas-filled channels in Elodea. Phys. Med. Biol. 1979, 24, 600–612. [Google Scholar] [CrossRef]
- McNeil, P.L.; Terasaki, M. Coping with the inevitable: How cells repair a torn surface membrane. Nat. Cell Biol. 2001, 3, E124–E129. [Google Scholar] [CrossRef]
- Meldolesi, J. Surface wound healing: A new, general function of eukaryotic cells. J. Cell Mol. Med. 2003, 7, 197–203. [Google Scholar] [CrossRef]
- Togo, T.; Alderton, J.M.; Bi, G.Q.; Steinhardt, R.A. The mechanism of facilitated cell membrane resealing. J. Cell Sci. 1999, 112, 719–731. [Google Scholar] [CrossRef]
- Schlicher, R.K.; Hutcheson, J.D.; Radhakrishna, H.; Apkarian, R.P.; Prausnitz, M.R. Changes in cell morphology due to plasma membrane wounding by acoustic cavitation. Ultrasound Med. Biol. 2010, 36, 677–692. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hosano, N.; Moosavi-Nejad, Z.; Satoh, M.; Hosano, H. Shock Waves Enhance Expression of Glycosphingolipid Tumor Antigen on Renal Cell Carcinoma: Dynamics of Physically Unmasking Hidden Intracellular Markers Independent of Gene-Signaling Pathways. Biomedicines 2022, 10, 545. https://doi.org/10.3390/biomedicines10030545
Hosano N, Moosavi-Nejad Z, Satoh M, Hosano H. Shock Waves Enhance Expression of Glycosphingolipid Tumor Antigen on Renal Cell Carcinoma: Dynamics of Physically Unmasking Hidden Intracellular Markers Independent of Gene-Signaling Pathways. Biomedicines. 2022; 10(3):545. https://doi.org/10.3390/biomedicines10030545
Chicago/Turabian StyleHosano, Nushin, Zahra Moosavi-Nejad, Makoto Satoh, and Hamid Hosano. 2022. "Shock Waves Enhance Expression of Glycosphingolipid Tumor Antigen on Renal Cell Carcinoma: Dynamics of Physically Unmasking Hidden Intracellular Markers Independent of Gene-Signaling Pathways" Biomedicines 10, no. 3: 545. https://doi.org/10.3390/biomedicines10030545
APA StyleHosano, N., Moosavi-Nejad, Z., Satoh, M., & Hosano, H. (2022). Shock Waves Enhance Expression of Glycosphingolipid Tumor Antigen on Renal Cell Carcinoma: Dynamics of Physically Unmasking Hidden Intracellular Markers Independent of Gene-Signaling Pathways. Biomedicines, 10(3), 545. https://doi.org/10.3390/biomedicines10030545