
Polycystic Ovarian Syndrome: A Complex Disease with a Genetics Approach


 , , , , ,
, , , , ,
Abstract
1. Introduction
2. Epidemiologic Profile of PCOS
3. Window of Susceptibility
3.1. PCOS in Adolescence
3.2. PCOS in Reproductive Age
3.3. PCOS in Menopausal Age
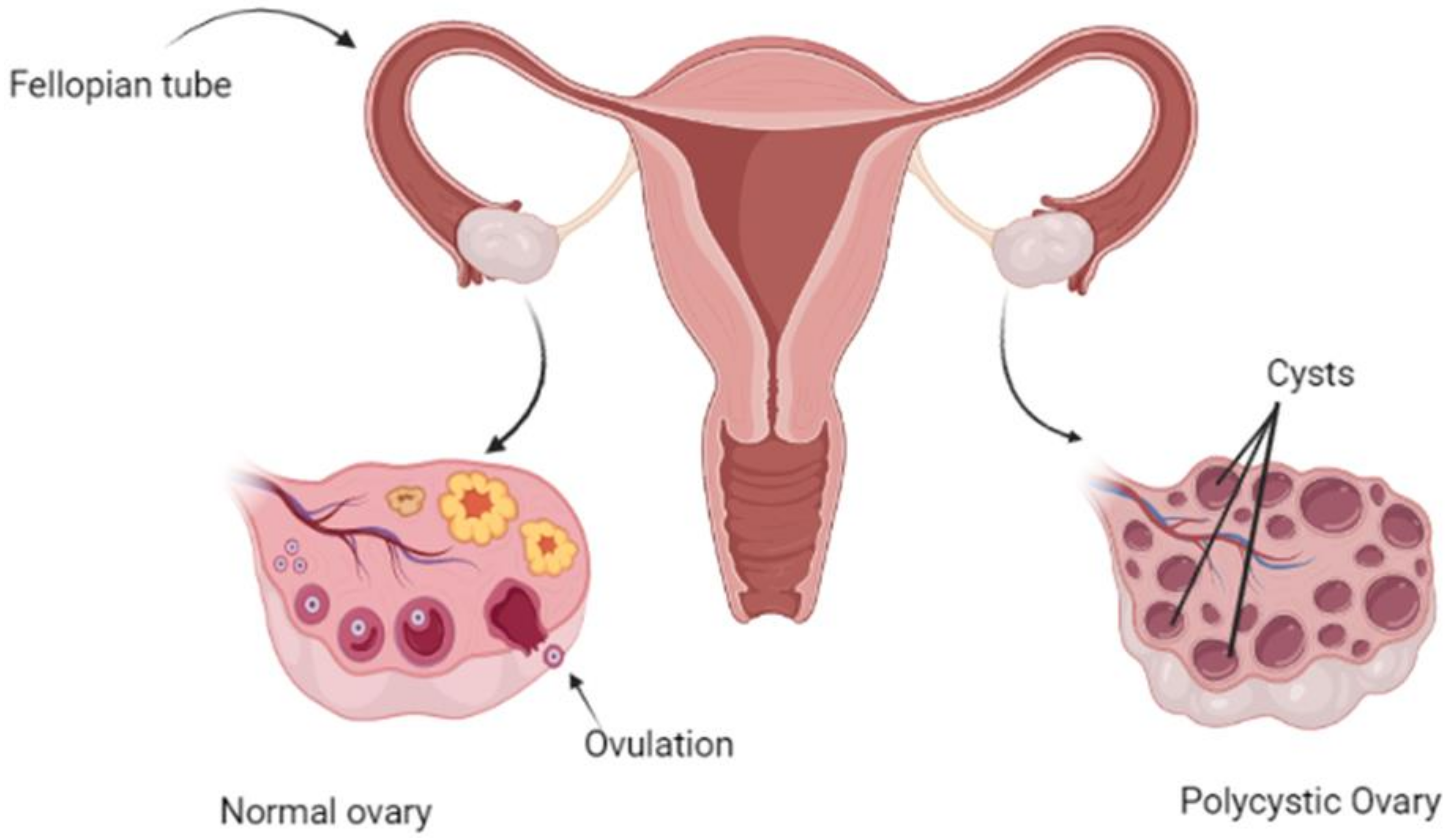
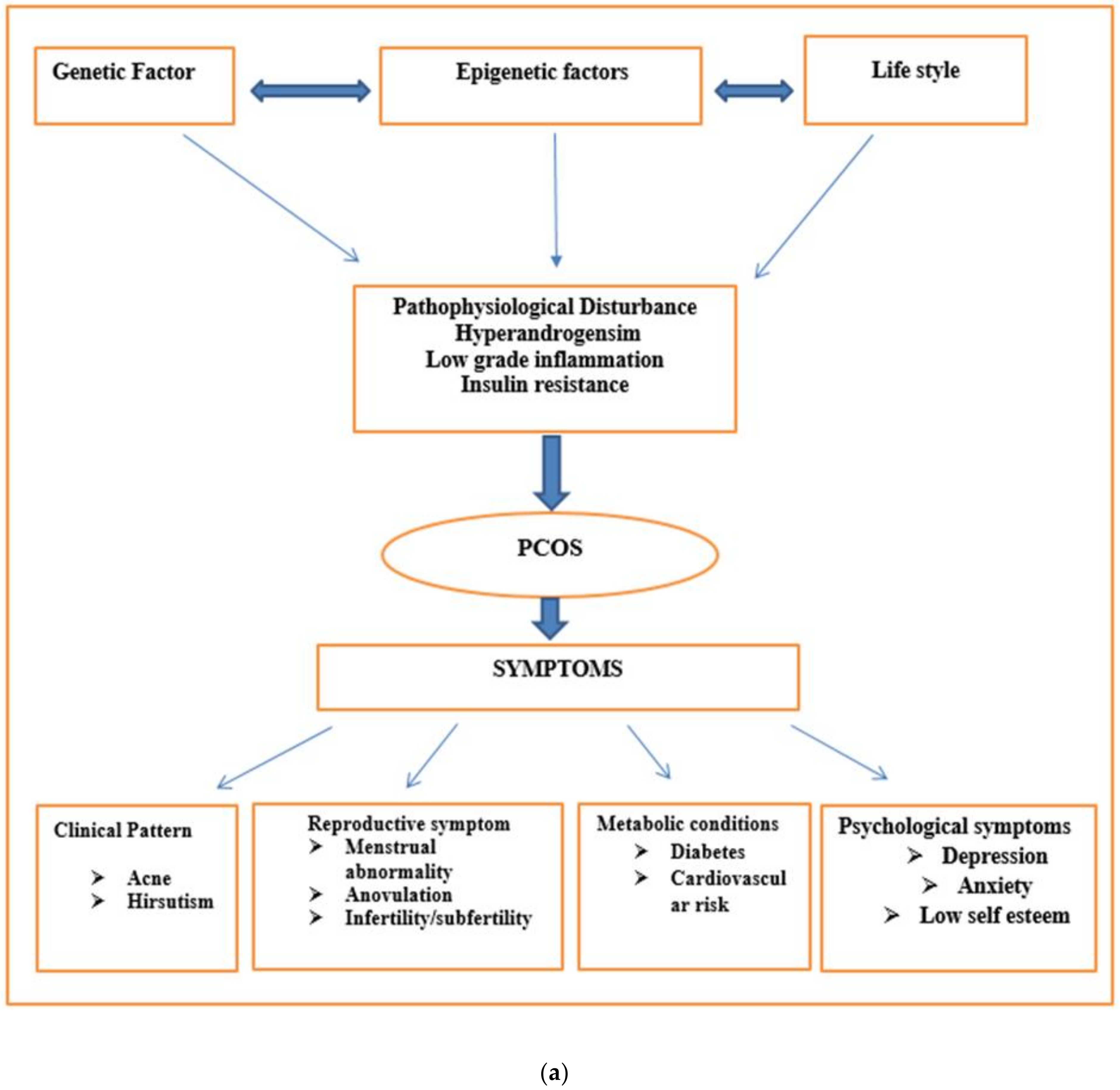
4. Clinical Pattern of Disease
5. Phenotypic Classification of PCOS
5.1. Classic Phenotype
5.2. Ovulatory PCOS
5.3. Non-Hyperandrogenic PCOS
- Hyperandrogenism, oligo anovulation, various polycystic ovaries morphology (through ultrasound)
- Hyperandrogenism, oligo anovulation, normal ovaries morphology (through ultrasound)
- Hyperandrogenism, normal menstruation cycle, various polycystic ovaries morphology (through ultrasound)
- Normal androgen, oligo anovulation, and polycystic ovaries morphology (through ultrasound)
6. PCOS Pathogenesis
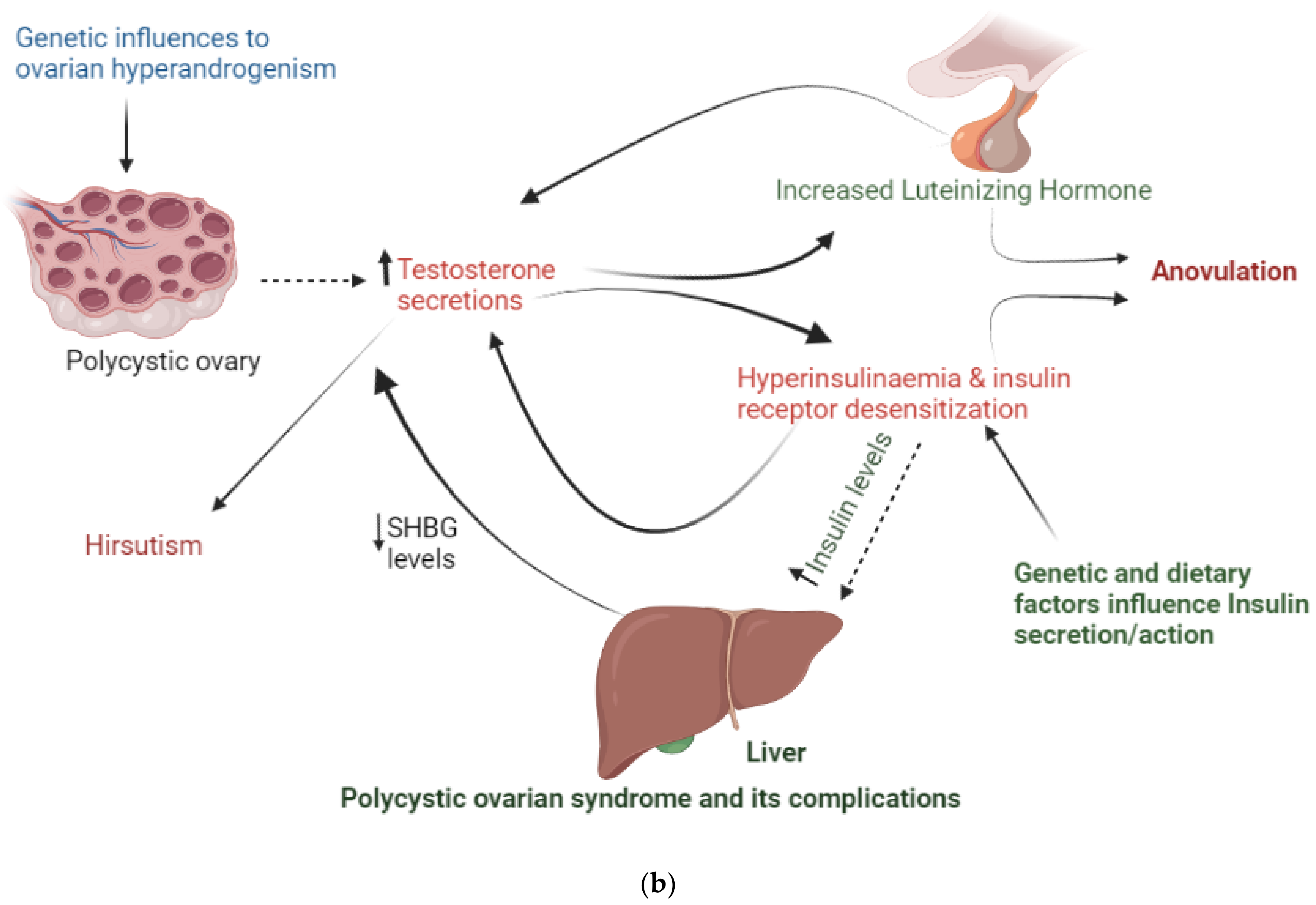
6.1. Hyperandrogenism
6.2. Ovarian Follicular Dysfunction
6.3. Neuroendocrine Imbalance
6.4. Insulin Resistance
7. Inheritance in PCOS
8. Genetic Heritability Reported in GWAS Studies
9. Specialized Genes Involved in PCOS Pathophysiology
9.1. Gene Involved in Ovarian and Adrenal Steroidogenesis
9.2. Epigenetics of PCOS
9.2.1. Peripubertal Diet and Epigenetics of PCOS
9.2.2. Prenatal Diet and Epigenetics of PCOS
9.3. Genes Involved in Insulin Action and Secretion
9.4. Genes Involved in Steroid Hormone Effect
9.4.1. Androgen Receptor Gene
9.4.2. Sex Hormone-Binding Globulin Gene
9.5. Gene Involved in Gonadotropin
9.5.1. Lutein Hormone and Receptor
9.5.2. Follicular Stimulating Hormone Receptor
9.5.3. Anti-Mullerian Hormone
9.6. Other Genes
9.6.1. PCO
9.6.2. SRD5A and SRD5B
9.6.3. Fat Mass Obesity (FTO)
10. Application of Genetic Identification
11. Treatment
11.1. Non-Pharmacological Approach
11.1.1. Weight Reduction
11.1.2. Diet
11.1.3. Exercise
11.1.4. Complementary and Alternative Medicines (CAM)
Acupuncture
Diet Therapy (Health Supplementation)
11.2. Medicinal Approach
12. Conclusions
13. Future Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hardiman, P.; Pillay, O.S.; Atiomo, W. Polycystic ovary syndrome and endometrial carcinoma. Lancet 2003, 361, 1810–1812. [Google Scholar] [CrossRef]
- Diamanti-Kandarakis, E.; Piperi, C.; Spina, J.; Papanastasiou, L.; Bergiele, A.; Panidis, D. Polycystic ovary syndrome: The influence of environmental and genetic factors. Hormones 2006, 5, 17. [Google Scholar] [CrossRef] [PubMed]
- Krysiak, R.; Okopie, B.; Gdula-Dymek, A.; Herman, Z.S. Update on the management of polycystic ovary syndrome. Pharmacol. Rep. 2006, 58, 614. [Google Scholar] [PubMed]
- Liu, A.L.; Xie, H.J.; Xie, H.Y.; Liu, J.; Yin, J.; Hu, J.S.; Peng, C.Y. Association between fat mass and obesity associated (FTO) gene rs9939609 A/T polymorphism and polycystic ovary syndrome: A systematic review and meta-analysis. BMC Med. Genet. 2017, 18, 89. [Google Scholar] [CrossRef]
- Deeks, A.A.; Gibson-Helm, M.E.; Paul, E.; Teede, H.J. Is having polycystic ovary syndrome a predictor of poor psychological function including anxiety and depression? Hum. Reprod. 2011, 26, 1399–1407. [Google Scholar] [CrossRef]
- Lowe, W.L.; Reddy, T.E. Genomic approaches for understanding the genetics of complex disease. Genome Res. 2015, 25, 1432–1441. [Google Scholar] [CrossRef]
- Feinberg, A.P. Phenotypic plasticity and the epigenetics of human disease. Nature 2007, 447, 433–440. [Google Scholar] [CrossRef]
- Hanash, S. Disease proteomics. Nature 2003, 422, 226–232. [Google Scholar] [CrossRef]
- Zawadzski, J.K. Diagnostic criteria for polycystic ovary syndrome: Towards a rational approach. In Polycystic Ovary Syndrome; Blackwell Scientific: Boston, MA, USA, 1992; pp. 39–50. [Google Scholar]
- Azziz, R.; Carmina, E.; Dewailly, D.; Diamanti-Kandarakis, E.; Escobar-Morreale, H.F.; Futterweit, W.; Janssen, O.E.; Legro, R.S.; Norman, R.J.; Taylor, A.E.; et al. Positions statement: Criteria for defining polycystic ovary syndrome as a predominantly hyperandrogenic syndrome: An Androgen Excess Society guideline. J. Clin. Endocrinol. Metab. 2006, 91, 4237–4245. [Google Scholar] [CrossRef]
- Eshre, T.R. ASRM-Sponsored PCOS Consensus Workshop Group. Revised 2003 consensus on diagnostic criteria and long-term health risks related to polycystic ovary syndrome. Fertil. Steril. 2004, 81, 19–25. [Google Scholar]
- Knochenhauer, E.S.; Key, T.J.; Kahsar-Miller, M.; Waggoner, W.; Boots, L.R.; Azziz, R. Prevalence of the polycystic ovary syndrome in unselected black and white women of the Southeastern United States: A prospective study. J. Clin. Endocrinol. Metab. 1998, 83, 3078–3082. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.L.; Kar, S.; Vanky, E.; Morin-Papunen, L.; Piltonen, T.; Puurunen, J.; Tapanainen, J.S.; Maciel, G.A.; Hayashida, S.A.; Soares, J.M., Jr.; et al. Racial and ethnic differences in the prevalence of metabolic syndrome and its components of metabolic syndrome in women with polycystic ovary syndrome: A regional cross-sectional study. Am. J. Obstet. Gynecol. 2017, 217, 189.e1–189.e8. [Google Scholar] [CrossRef] [PubMed]
- Asunción, M.; Calvo, R.M.; Njls, M.; Sancho, J.; Avila, S.; Escobar-Morreale, H.F. A prospective study of the prevalence of the polycystic ovary syndrome in unselected Caucasian women from Spain. J. Clin. Endocrinol. Metab. 2000, 85, 2434–2438. [Google Scholar] [CrossRef]
- Khan, M.J.; Ullah, A.; Basit, S. Genetic Basis of Polycystic Ovary Syndrome (PCOS) Current Perspectives. Appl. Clin. Genet. 2019, 12, 249. [Google Scholar] [CrossRef]
- Li, R.; Zhang, Q.; Yang, D.; Li, S.; Lu, S.; Wu, X.; Wei, Z.; Song, X.; Wang, X.; Fu, S.; et al. Prevalence of polycystic ovary syndrome in women in China: A large community-based study. Hum. Reprod. 2013, 28, 2562–2569. [Google Scholar] [CrossRef]
- Nidhi, R.; Padmalatha, V.; Nagarathna, R.; Amritanshu, R. Prevalence of polycystic ovarian syndrome in Indian adolescents. J. Pediatr. Adolesc. Gynecol. 2011, 24, 223–227. [Google Scholar] [CrossRef]
- Roya, R.; Mohammad Akbar, A.; Wajeeda, T.; Avinash, B.; Humaira, M.; Avvari, B.B.; Ayapati, M.G.; Vikram, A.A.; Taalia, N.A.; Mohammed, S.A. An Indian Evidence-Based Study of Prevalence, Phenotypic Features, Lifestyle Modifications of Polycystic Ovarian Syndrome Patients. J. Gynecol. Women’s Health 2021, 21, 556069. [Google Scholar] [CrossRef]
- Akram, M.; Roohi, N. Endocrine correlates of polycystic ovary syndrome in Pakistani women. J. Coll. Physicians Surg. Pak. 2015, 25, 22–26. [Google Scholar]
- Baqai, Z.; Khanam, M.; Parveen, S. Prevalence of PCOS in infertile patients. Med. Chanel 2010, 16, 437–440. [Google Scholar]
- Louwers, Y.V.; Laven, J.S. Characteristics of polycystic ovary syndrome throughout life. Ther. Adv. Reprod. Health 2020, 14. [Google Scholar] [CrossRef]
- Apter, D. Endocrine and metabolic abnormalities in adolescents with a PCOS-like condition: Consequences for adult reproduction. Trends. Endocrinol. Metab. 1998, 9, 58–61. [Google Scholar] [CrossRef]
- van Hooff, M.H.; Voorhorst, F.J.; Kaptein, M.B.; Hirasing, R.A.; Koppenaal, C.; Schoemaker, J. Predictive value of menstrual cycle pattern, body mass index, hormone levels and polycystic ovaries at age 15 years for oligo-amenorrhoea at age 18 years. Hum. Reprod. 2004, 19, 383–392. [Google Scholar] [CrossRef]
- Teede, H.J.; Misso, M.L.; Costello, M.F.; Dokras, A.; Laven, J.; Moran, L.; Piltonen, T.; Norman, R.J. Recommendations from the international evidence-based guideline for the assessment and management of polycystic ovary syndrome. Hum. Reprod. 2018, 33, 1602–1618. [Google Scholar] [CrossRef]
- Brassard, M.; AinMelk, Y.; Baillargeon, J.P. Basic infertility including polycystic ovary syndrome. Med. Clin. N. Am. 2008, 92, 1163–1192. [Google Scholar] [CrossRef]
- Joham, A.E.; Teede, H.J.; Ranasinha, S.; Zoungas, S.; Boyle, J. Prevalence of infertility and use of fertility treatment in women with polycystic ovary syndrome: Data from a large community-based cohort study. J. Womens Health 2015, 24, 299–307. [Google Scholar] [CrossRef]
- Christ, J.P.; Gunning, M.N.; Meun, C.; Eijkemans, M.J.; van Rijn, B.B.; Bonsel, G.J.; Laven, J.S.; Fauser, B.C. Preconception characteristics predict obstetrical and neonatal outcomes in women with polycystic ovary syndrome. J. Clin. Endocrinol. Metab. 2019, 104, 809–818. [Google Scholar] [CrossRef]
- Qin, J.Z.; Pang, L.H.; Li, M.J.; Fan, X.J.; Huang, R.D.; Chen, H.Y. Obstetric complications in women with polycystic ovary syndrome: A systematic review and meta-analysis. Reprod. Biol. Endocrinol. 2013, 11, 56. [Google Scholar] [CrossRef]
- Glueck, C.J.; Goldenberg, N. Characteristics of obesity in polycystic ovary syndrome: Etiology, treatment, and genetics. Metabolism 2019, 92, 108–120. [Google Scholar] [CrossRef]
- Brown, Z.A.; Louwers, Y.V.; Fong, S.L.; Valkenburg, O.; Birnie, E.; de Jong, F.H.; Fauser, B.C.; Laven, J.S. The phenotype of polycystic ovary syndrome ameliorates with aging. Fertil. Steril. 2011, 96, 1259–1265. [Google Scholar] [CrossRef]
- Pellatt, L.; Hanna, L.; Brincat, M.; Galea, R.; Brain, H.; Whitehead, S.; Mason, H. Granulosa cell production of anti-Müllerian hormone is increased in polycystic ovaries. J. Clin. Endocrinol. Metab. 2007, 92, 240–245. [Google Scholar] [CrossRef]
- Azziz, R.; Carmina, E.; Dewailly, D.; Diamanti-Kandarakis, E.; Escobar-Morreale, H.F.; Futterweit, W.; Janssen, O.E.; Legro, R.S.; Norman, R.J.; Taylor, A.E.; et al. The androgen excess and PCOS Society criteria for the polycystic ovary syndrome: The complete task force report. Fertil. Steril. 2009, 91, 456–488. [Google Scholar] [CrossRef]
- Villarroel, C.; Merino, P.M.; López, P.; Eyzaguirre, F.C.; Van Velzen, A.; Iniguez, G.; Codner, E. Polycystic ovarian morphology in adolescents with regular menstrual cycles is associated with elevated anti-Mullerian hormone. Hum. Reprod. 2011, 26, 2861–2868. [Google Scholar] [CrossRef]
- Webber, L.J.; Stubbs, S.; Stark, J.; Trew, G.H.; Margara, R.; Hardy, K.; Franks, S. Formation and early development of follicles in the polycystic ovary. Lancet 2003, 362, 1017–1021. [Google Scholar] [CrossRef]
- Das, M.; Djahanbakhch, O.; Hacihanefioglu, B.; Saridogan, E.; Ikram, M.; Ghali, L.; Raveendran, M.; Storey, A. Granulosa cell survival and proliferation are altered in polycystic ovary syndrome. J. Clin. Endocrinol. Metab. 2008, 93, 881–887. [Google Scholar] [CrossRef]
- Dunaif, A. Insulin resistance and the polycystic ovary syndrome: Mechanism and implications for pathogenesis. Endocr. Rev. 1997, 7, 345–349. [Google Scholar]
- Cortón, M.; Botella-Carretero, J.I.; Benguría, A.; Villuendas, G.; Zaballos, A.; San Millán, J.L.; Escobar-Morreale, H.F.; Peral, B. Differential gene expression profile in omental adipose tissue in women with polycystic ovary syndrome. J. Clin. Endocrinol. Metab. 2007, 92, 328–337. [Google Scholar] [CrossRef]
- González, F.; Rote, N.S.; Minium, J.; Kirwan, J.P. Reactive oxygen species-induced oxidative stress in the development of insulin resistance and hyperandrogenism in polycystic ovary syndrome. J. Clin. Endocrinol. Metab. 2006, 91, 336–340. [Google Scholar] [CrossRef]
- Lizneva, D.; Suturina, L.; Walker, W.; Brakta, S.; Gavrilova-Jordan, L.; Azziz, R. Criteria, prevalence, and phenotypes of polycystic ovarysyndrome. Fertil. Steril. 2016, 106, 6–15. [Google Scholar] [CrossRef]
- Goverde, A.J.; Van Koert, A.J.B.; Eijkemans, M.J.; Knauff, E.A.; Westerveld, H.E.; Fauser, B.C.; Broekmans, F.J. Indicators for metabolic disturbances in anovulatory women with polycystic ovary syndrome diagnosed according to the Rotterdam consensus criteria. Hum. Reprod. 2009, 24, 710–717. [Google Scholar] [CrossRef]
- Sahmay, S.; Atakul, N.; Oncul, M.; Tuten, A.; Aydogan, B.; Seyisoglu, H. Serumanti-mullerian hormone levels in the main phenotypes of polycystic ovary syndrome. Eur. J. Obstet. Gynecol. Reprod. Biol. 2013, 170, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Di Fede, G.; Mansueto, P.; Longo, R.A.; Rini, G.; Carmina, E. Influence of sociocultural factors on the ovulatory status of polycystic ovary syndrome. Fertil. Steril. 2009, 91, 1853–1856. [Google Scholar] [CrossRef] [PubMed]
- Guastella, E.; Longo, R.A.; Carmina, E. Clinical and endocrine characteristics of the main polycystic ovary syndrome phenotypes. Fertil. Steril. 2010, 94, 2197–2201. [Google Scholar] [CrossRef]
- Zhang, H.Y.; Zhu, F.F.; Xiong, J.; Shi, X.B.; Fu, S.X. Characteristics of different phenotypes of polycystic ovary syndrome based on the Rotterdam criteria in a large-scale Chinese population. BJOG Int. J. Obs. Gynaecol. 2009, 116, 1633–1639. [Google Scholar] [CrossRef]
- Yilmaz, M.; Isaoglu, U.; Delibas, I.B.; Kadanali, S. Anthropometric, clinical and laboratory comparison of four phenotypes of polycystic ovary syndrome based on Rotterdam criteria. J. Obstet. Gynaecol. Res. 2011, 37, 1020–1026. [Google Scholar] [CrossRef]
- Moghetti, P.; Tosi, F.; Bonin, C.; Di Sarra, D.; Fiers, T.; Kaufman, J.M.; Giagulli, V.A.; Signori, C.; Zambotti, F.; Dall’Alda, M.; et al. Divergences in insulin resistance between the different phenotypes of the polycystic ovary syndrome. J. Clin. Endocrinol. Metab. 2013, 98, E628–E637. [Google Scholar] [CrossRef] [PubMed]
- Dewailly, D.; Catteau-Jonard, S.; Reyss, A.-C.; Leroy, M.; Pigny, P. Oligoanovulation with polycystic ovaries but not overt hyperandrogenism. J. Clin. Endocrinol. Metab. 2006, 91, 3922–3927. [Google Scholar] [CrossRef]
- Jamil, A.S.; Alalaf, S.K.; Al-Tawil, N.G.; Al-Shawaf, T. Comparison of clinical and hormonal characteristics among four phenotypes of polycystic ovary syndrome based on the Rotterdam criteria. Arch. Gynecol. Obstet. 2016, 293, 447–456. [Google Scholar] [CrossRef]
- Panidis, D.; Tziomalos, K.; Papadakis, E.; Chatzis, P.; Kandaraki, E.A.; Tsourdi, E.A.; Macut, D.; Bjekic-Macut, J.; Marthopoulos, A.; Katsikis, I. Associations of menstrual cycle irregularities with age, obesity and phenotype in patients with polycystic ovary syndrome. Hormones 2015, 14, 431–437. [Google Scholar] [CrossRef]
- Rosenfield, R.L.; Ehrmann, D.A. The Pathogenesis of Polycystic Ovary Syndrome (PCOS): The Hypothesis of PCOS as Functional Ovarian Hyperandrogenism Revisited. Endocr. Rev. 2016, 37, 467–520. [Google Scholar] [CrossRef]
- Nelson, V.L.; Legro, R.S.; Strauss, J.F.; McAllister, J.M. Augmented androgen production is a stable steroidogenic phenotype of propagated theca cells from polycystic ovaries. Mol. Endocrinol. 1999, 13, 946–957. [Google Scholar] [CrossRef]
- Goodarzi, M.O.; Carmina, E.; Azziz, R. DHEA, DHEAS and PCOS. J. Steroid. Biochem. Mol. Biol. 2015, 145, 213–225. [Google Scholar] [CrossRef] [PubMed]
- Louwers, Y.V.; de Jong, F.H.; van Herwaarden, N.A.A.; Stolk, L.; Fauser, B.C.J.M.; Uitterlinden, A.G.; Laven, J.S.E. Variants in SULT2A1 affect the DHEA sulphate to DHEA ratio in patients with polycystic ovary syndrome but not the hyperandrogenic phenotype. J. Clin. Endocrinol. Metab. 2013, 98, 3848–3855. [Google Scholar] [CrossRef] [PubMed]
- Draper, N.; Walker, E.A.; Bujalska, I.J.; Tomlinson, J.W.; Chalder, S.M.; Arlt, W.; Lavery, G.G.; Bedendo, O.; Ray, D.W.; Laing, I.; et al. Mutations in the genes encoding 11beta-hydroxysteroid dehydrogenase type 1 and hexose-6-phosphate dehydrogenase interact to cause cortisone reductase deficiency. Nat. Genet. 2003, 34, 434–439. [Google Scholar] [CrossRef] [PubMed]
- Garg, D.; Tal, R. The role of AMH in the pathophysiology of polycystic ovarian syndrome. Reprod. Biomed. Online 2016, 33, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Azziz, R.; Carmina, E.; Chen, Z.; Dunaif, A.; Laven, J.S.; Legro, R.S.; Lizneva, D.; Natterson-Horowtiz, B.; Teede, H.J.; Yildiz, B.O. Polycystic ovary syndrome. Nat. Rev. Dis. Primers 2016, 2, 685–697. [Google Scholar] [CrossRef] [PubMed]
- Fallat, M.E.; Siow, Y.; Marra, M.; Cook, C.; Carrillo, A. Müllerian-inhibiting substance in follicular fluid and serum: A comparison of patients with tubal factor infertility, polycystic ovary syndrome, and endometriosis. Fertil. Steril. 1997, 67, 962–965. [Google Scholar] [CrossRef]
- Dixit, H.; Rao, L.K.; Padmalatha, V.V.; Kanakavalli, M.; Deenadayal, M.; Gupta, N.; Chakrabarty, B.; Singh, L. Missense mutations in the BMP15 gene are associated with ovarian failure. Hum. Genet. 2006, 119, 408–415. [Google Scholar] [CrossRef]
- Mikaeili, S.; Rashidi, B.H.; Safa, M.; Najafi, A.; Sobhani, A.; Asadi, E.; Abbasi, M. Altered FoxO3 expression and apoptosis in granulosa cells of women with polycystic ovary syndrome. Arch. Gynecol. Obstet. 2016, 294, 185–192. [Google Scholar] [CrossRef]
- Pastor, C.L.; Griffin-Korf, M.L.; Aloi, J.A.; Evans, W.S.; Marshall, J.C. Polycystic ovary syndrome: Evidence for reduced sensitivity of the gonadotropin-releasing hormone pulse generator to inhibition by estradiol and progesterone. J. Clin. Endocrinol. Metab. 1998, 83, 582–590. [Google Scholar] [CrossRef]
- Goodarzi, M.O.; Dumesic, D.A.; Chazenbalk, G.; Azziz, R. Polycystic ovary syndrome: Etiology, pathogenesis and diagnosis. Nat. Rev. Endocrinol. 2011, 7, 219–231. [Google Scholar] [CrossRef] [PubMed]
- Trikudanathan, S. Polycystic ovarian syndrome. Med. Clin. N. Am. 2015, 99, 221–235. [Google Scholar] [CrossRef] [PubMed]
- Hayes, M.G.; Urbanek, M.; Ehrmann, D.A.; Armstrong, L.L.; Lee, J.Y.; Sisk, R.; Karaderi, T.; Barber, T.M.; McCarthy, M.I.; Franks, S.; et al. Genome-wide association of polycystic ovary syndrome implicates alterations in gonadotropin secretion in European ancestry populations. Nat. Commun. 2015, 6, 7502. [Google Scholar] [CrossRef]
- Day, F.R.; Hinds, D.A.; Tung, J.Y.; Stolk, L.; Styrkarsdottir, U.; Saxena, R.; Bjonnes, A.; Broer, L.; Dunger, D.B.; Halldorsson, B.V.; et al. Causal mechanisms and balancing selection inferred from genetic associations with polycystic ovary syndrome. Nat. Commun. 2015, 6, 8464. [Google Scholar] [CrossRef]
- Shi, Y.; Zhao, H.; Shi, Y.; Cao, Y.; Yang, D.; Li, Z.; Zhang, B.; Liang, X.; Li, T.; Chen, J.; et al. Genome-wide association study identifies eight new risk loci for polycystic ovary syndrome. Nat. Genet. 2012, 44, 1020–1025. [Google Scholar] [CrossRef]
- Chen, Z.J.; Zhao, H.; He, L.; Shi, Y.; Qin, Y.; Shi, Y.; Li, Z.; You, L.; Zhao, J.; Liu, J.; et al. Genome-wide association study identifies susceptibility loci for polycystic ovary syndrome on chromosome 2p16.3, 2p21 and 9q33.3. Nat. Gene 2011, 43, 55–59. [Google Scholar] [CrossRef]
- Day, F.; Karaderi, T.; Jones, M.R.; Meun, C.; He, C.; Drong, A.; Kraft, P.; Lin, N.; Huang, H.; Broer, L.; et al. Large-scale genome-wide meta-analysis of polycystic ovary syndrome suggests shared genetic architecture for di_fferent diagnosis criteria. PLoS Genet. 2018, 14, e1007813. [Google Scholar] [CrossRef]
- Hwang, J.Y.; Lee, E.J.; Jin, G.M.; Sung, Y.A.; Lee, H.J.; Heon, K.S.; Jang, H.C.; Soo Park, K.; Lee, H.J.; Byul Jang, H.; et al. Genome-wide association study identifies GYS2 as a novel genetic factor for polycystic ovary syndrome through obesity-related condition. J. Hum. Genet. 2012, 57, 660–664. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Oh, J.Y.; Sung, Y.A.; Chung, H.; Kim, H.L.; Kim, G.S.; Cho, Y.S.; Kim, J.T. Genome-wide association study identified new susceptibility loci for polycystic ovary syndrome. Hum. Reprod. 2015, 30, 723–731. [Google Scholar] [CrossRef]
- Wilkening, S.; Chen, B.; Bermejo, J.L.; Canzian, F. Is there still a need for candidate gene approaches in the era of genome-wide association studies? Genomics 2009, 93, 415–419. [Google Scholar] [CrossRef]
- Mykhalchenko, K.; Lizneva, D.; Trofimova, T.; Walker, W.; Suturina, L.; Diamond, M.P.; Azziz, R. Genetics of polycystic ovary syndrome. Expert Rev. Mol. Diagn. 2017, 17, 723–733. [Google Scholar] [CrossRef] [PubMed]
- Zondervan, K.T.; Cardon, L.R. Designing candidate gene and genome-wide case-control association studies. Nat. Protoc. 2007, 2, 2492–2501. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.J.; Williams, M.G.; Eynon, N.; Ashton, K.J.; Little, J.P.; Wislo, U.; Coombes, J.S. Genes to predict VO2max trainability: A systematic review. BMC Genom. 2017, 18, 81–110. [Google Scholar] [CrossRef]
- Sagoo, G.S.; Little, J.; Higgins, J.P.T. Systematic Reviews of Genetic Association Studies. PLoS Med. 2009, 6, e1000028. [Google Scholar] [CrossRef] [PubMed]
- Lewis, C.M. Genetic association studies: Design, analysis and interpretation. Brief. Bioinform. 2002, 3, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, B.; Vellanki, P.; Ata, B.; Yildiz, B.O. Metabolic syndrome, hypertension, and hyperlipidemia in mothers, fathers, sisters, and brothers of women with polycystic ovary syndrome: A systematic review and meta-analysis. Fertil. Steril. 2018, 109, 356–364. [Google Scholar] [CrossRef] [PubMed]
- McAllister, J.M.; Modi, B.; Miller, B.A.; Biegler, J.; Bruggeman, R.; Legro, R.S.; Strauss, J.F. Overexpression of a DENND1A isoform produces a polycystic ovary syndrome theca phenotype. Proc. Natl. Acad. Sci. USA 2014, 111, E1519–E1527. [Google Scholar] [CrossRef]
- McAllister, J.M.; Legro, R.S.; Modi, B.P.; Strauss, J.F. Functional genomics of PCOS: From GWAS to molecular mechanisms. Trends. Endocrinol. Metab. 2015, 26, 118–124. [Google Scholar] [CrossRef]
- Toledo, S.P.; Brunner, H.G.; Kraaij, R.; Post, M.; Dahia, P.L.; Hayashida, C.Y.; Kremer HThemmen, A.P. An inactivating mutation of the luteinizing hormone receptor causes amenorrhea in a 46,XX female. J. Clin. Endocrinol. Metab. 1996, 81, 3850–3854. [Google Scholar]
- Latronico, A.C.; Chai, Y.; Arnhold, I.J.; Liu, X.; Mendonca, B.B.; Segaloff, D.L. A homozygous microdeletion in helix 7 of the luteinizing hormone receptor associated with familial testicular and ovarian resistance is due to both decreased cell surface expression and impaired effector activation by the cell surface receptor. Mol. Endocrinol. 1998, 12, 442–450. [Google Scholar] [CrossRef]
- Latronico, A.C.; Lins, T.S.; Brito, V.N.; Arnhold, I.J.; Mendonca, B.B. The effect of distinct activating mutations of the luteinizing hormone receptor gene on the pituitary-gonadal axis in both sexes. Clin. Endocrinol. 2000, 53, 609–613. [Google Scholar] [CrossRef]
- Overbeek, A.; Kuijper, E.A.; Hendriks, M.L.; Blankenstein, M.A.; Ketel, I.J.; Twisk, J.W.; Hompes, P.G.; Homburg, R.; Lambalk, C.B. Clomiphene citrate resistance in relation to follicle-stimulating hormone receptor Ser680Ser-polymorphism in polycystic ovary syndrome. Hum. Reprod. 2009, 24, 2007–2013. [Google Scholar] [CrossRef] [PubMed]
- Voight, B.F.; Scott, L.J.; Steinthorsdottir, V.; Morris, A.P.; Dina, C.; Welch, R.P.; Zeggini, E.; Huth, C.; Aulchenko, Y.S.; Thorleifsson, G.; et al. Twelve type 2 diabetes susceptibility loci identified through large-scale association analysis. Nat. Genet. 2010, 42, 579–589. [Google Scholar] [CrossRef] [PubMed]
- Barrett, J.C.; Clayton, D.G.; Concannon, P.; Akolkar, B.; Cooper, J.D.; Erlich, H.A.; Julier, C.; Morahan, G.; Nerup, J.; Nierras, C.; et al. Genome-wide association study and meta-analysis find that over 40 loci affect risk of type 1 diabetes. Nat. Genet. 2009, 41, 703–707. [Google Scholar] [CrossRef]
- Li, T.; Zhao, H.; Zhao, X.; Zhang, B.; Cui, L.; Shi, Y.; Li, G.; Wang, P.; Chen, Z.J. Identification of YAP1 as a novel susceptibility gene for polycystic ovary syndrome. J. Med. Genet. 2012, 49, 254–257. [Google Scholar] [CrossRef]
- Takayama, K.; Suzuki, T.; Bulun, S.E.; Sasano, H.; Yilmaz, B.; Sebastian, S. Organization of the human aromatase p450 (CYP19) gene. In Seminars in Reproductive Medicine; Thieme Medical Publishers, Inc.: New York, NY, USA, 2004; Volume 22, pp. 5–9. [Google Scholar]
- Carey, A.H.; Waterworth, D.; Patel, K.; White, D.; Little, J.; Novelli, P.; Franks, S.; Williamson, R. Polycystic ovaries and premature male pattern baldness are associated with one allele of the steroid metabolism gene CYP17. Hum. Mol. Genet. 1994, 3, 1873–1876. [Google Scholar] [CrossRef]
- Rosenfield, R.L.; Barnes, R.B.; Cara, J.F.; Lucky, A.W. Dysregulation of cytochrome P450c17α as the cause of polycystic ovarian syndrome. Fertil. Steril. 1990, 53, 785–791. [Google Scholar] [CrossRef]
- Wickenheisser, J.K.; Quinn, P.G.; Nelson, V.L.; Legro, R.S.; Strauss, J.F.; Mcallister, J.M. Differential activity of the cytochrome P450 17α- hydroxylase and steroidogenic acute regulatory protein gene promoters in normal and polycystic ovary syndrome theca cells. J. Clin. Endocrinol. Metab. 2000, 85, 2304–2311. [Google Scholar] [CrossRef]
- Witchel, S.F.; Aston, C.E. The role of heterozygosity for CYP21 in the polycystic ovary syndrome. J. Pediatr. Endocrinol. Metab. 2000, 13, 1315–1317. [Google Scholar]
- Franks, S.; Gilling-Smith, C.; Gharani, N.; McCarthy, M. Pathogenesis of polycystic ovary syndrome: Evidence for a genetically determined disorder of ovarian androgen production. Hum. Fertil. 2000, 3, 77–79. [Google Scholar] [CrossRef]
- Gharani, N.; Waterworth, D.M.; Batty, S.; White, D.; Gilling-Smith, C.; Conway, G.S.; McCarthy, M.; Franks, S.; Williamson, R. Association of the Steroid Synthesis Gene Cyp11a with Polycystic Ovary Syndrome and Hyperandrogenism. Hum. Mol. Genet. 1997, 6, 397–402. [Google Scholar] [CrossRef] [PubMed]
- Diamanti-Kandarakis, E.; Bartzis, M.I.; Bergiele, A.T.; Tsianateli, T.C.; Kouli, C.R. Microsatellite polymorphism (tttta)(n) at −528 base pairs of gene CYP11α influences hyperandrogenemia in patients with polycystic ovary syndrome. Fertil. Steril. 2000, 73, 735–741. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, X.K.; Cao, Y.; Yi, L.; Chen, J. A microsatellite polymorphism (tttta)n in the promoter of the CYP11a gene in Chinese women with polycystic ovary syndrome. Fertil. Steril. 2006, 86, 223–226. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Azziz, R.; Goodarzi, M.O. Epigenetics in polycystic ovary syndrome: A pilot study of global DNA methylation. Fertil. Steril. 2010, 94, 781–783.e1. [Google Scholar] [CrossRef]
- Wang, F.; Yu, B.; Yang, W.; Liu, J.; Lu, J.; Xia, X. Polycystic ovary syndrome resembling histopathological alterations in ovaries from prenatal androgenized female rats. J. Ovarian Res. 2012, 5, 15. [Google Scholar] [CrossRef]
- Abbott, D.H.; Tarantal, A.F.; Dumesic, D.A. Fetal, infant, adolescent and adult phenotypes of polycystic ovary syndrome in prenatally androgenized female rhesus monkeys. Am. J. Primatol. 2009, 71, 776–784. [Google Scholar] [CrossRef]
- Hague, W.M.; Adams, J.; Rodda, C.; Brook, C.G.D.; DE Bruyn, R.; Grant, D.B.; Jacobs, H.S. The prevalence of polycystic ovaries in patients with congenital adrenal hyperplasia and their close relatives. Clin. Endocrinol. 1990, 33, 501–510. [Google Scholar] [CrossRef]
- Mukherjee, S. Pathomechanisms of polycystic ovary syndrome multidimensional approaches. Front. Biosci. 2018, 10, 384–422. [Google Scholar] [CrossRef]
- Vázquez-Martínez, E.R.; Gómez-Viais, Y.I.; García-Gómez, E.; Reyes-Mayoral, C.; Reyes-Muñoz, E.; Camacho-Arroyo, I.; Cerbón, M.A. DNA Methylation in the Pathogenesis of Polycystic Ovary Syndrome. Reproduction 2019, 158, R27–R40. [Google Scholar] [CrossRef]
- Xu, J.; Bao, X.; Peng, Z.; Wang, L.; Du, L.; Niu, W.; Sun, Y. Comprehensive analysis of genome-wide DNA methylation across human polycystic ovary syndrome ovary granulosa cell. Oncotarget 2016, 7, 27899–27909. [Google Scholar] [CrossRef]
- Stueve, T.R.; Wol, M.S.; Pajak, A.; Teitelbaum, S.L.; Chen, J. Cyp19a1 promoter methylation in saliva associated with milestones of pubertal timing in urban girls. BMC Pediatr. 2014, 14, 78. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Peterson, K.E.; Sánchez, B.N.; Dolinoy, D.C.; Mercado-Garcia, A.; Téllez-Rojo, M.M.; Goodrich, J.M. Association of blood leukocyte DNA methylation at line-1 and growth-related candidate genes with pubertal onset and progression. Epigenetics 2018, 13, 1222–1233. [Google Scholar] [CrossRef] [PubMed]
- Vilchez, P.; Jaramillo, S.; Sanchez-Rodriguez, E.; Vargas, F.; Montoro-Molina, S.; Romero, M.; Espejo-Calvo, J.A.; Olmo-García, L.; Carrasco-Pancorbo, A.; De La Torre, R. Cardioprotective effect of a virgin olive oil enriched with bioactive compounds in spontaneously hypertensive rats. Nutrients 2019, 11, 1728. [Google Scholar] [CrossRef]
- Tomizawa, H.; Matsuzawa, D.; Ishii, D.; Matsuda, S.; Kawai, K.; Mashimo, Y.; Sutoh, C.; Shimizu, E. Methyl-donor deficiency in adolescence affects memory and epigenetic status in the mouse hippocampus. Genes Brain Behav. 2015, 14, 301–309. [Google Scholar] [CrossRef]
- Jia, L.; Li, J.; He, B.; Jia, Y.; Niu, Y.; Wang, C.; Zhao, R. Abnormally activated one-carbon metabolic pathway is associated with mtDNA hypermethylation and mitochondrial malfunction in the oocytes of polycystic gilt ovaries. Sci. Rep. 2016, 6, 19436. [Google Scholar] [CrossRef] [PubMed]
- Clare, C.E.; Brassington, A.H.; Kwong, W.Y.; Sinclair, K.D. One-carbon metabolism: Linking nutritional biochemistry to epigenetic programming of long-term development. Annu. Rev. Anim. Biosci. 2019, 7, 263–287. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, E.E.; Sadler-Riggleman, I.; Skinner, M.K. Environmentally induced epigenetic transgenerational inheritance of disease. Environ. Epigenet. 2018, 4, dvy016. [Google Scholar] [CrossRef]
- Abbott, D.H.; Dumesic, D.A.; Levine, J.E. Hyperandrogenic origins of polycystic ovary syndrome—Implications for pathophysiology and therapy. Expert Rev. Endocrinol. Metab. 2019, 14, 131–143. [Google Scholar] [CrossRef]
- Zhang, D.; Cong, J.; Shen, H.; Wu, Q.; Wu, X. Genome-wide identification of aberrantly methylated promoters in ovarian tissue of prenatally androgenized rats. Fertil. Steril. 2014, 102, 1458–1467. [Google Scholar] [CrossRef]
- Available online: https://www.genecards.org/cgi-bin/carddisp.pl?gene=CAPN10 (accessed on 29 January 2022).
- Sáez, M.E.; González-Sánchez, J.L.; Ramírez-Lorca, R.; Martínez-Larrad, M.T.; Zabena, C.; Gonzalez, A.; Moron, F.J.; Ruiz, A.; Serrano-Ríos, M. The CAPN10 gene is associated with insulin resistance phenotypes in the Spanish population. PLoS ONE 2008, 3, e2953. [Google Scholar] [CrossRef]
- Ajmal, N.; Khan, S.Z.; Shaikh, R. European journal of obstetrics & gynecology and reproductive biology: X polycystic ovary syndrome (PCOS) and genetic predisposition: A review article. Eur. J. Obstet. Gynecol. Reprod. Biol. X 2019, 3, 100060. [Google Scholar] [CrossRef] [PubMed]
- Hanis, C.; Boerwinkle, E.; Chakraborty, R.; Ellsworth, D.; Concannon, P.; Stirling, B.; Morrison, V.; Wapelhorst, B.; Spielman, R.; Gogolin-Ewens, K.; et al. A genome–wide search for human non–insulin–dependent (type 2) diabetes genes reveals a major susceptibility locus on chromosome 2. Nat. Genet. 1996, 13, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Xiao, J.; Zhao, X.; Liu, C.; Chen, Q. Four polymorphisms of the CAPN 10 gene and their relationship to polycystic ovary syndrome susceptibility: A meta-analysis. Clin. Endocrinol. 2012, 76, 431–438. [Google Scholar] [CrossRef]
- Shen, W.; Li, T.; Hu, Y.; Liu, H.; Song, M. Calpain-10 genetic polymorphisms and polycystic ovary syndrome risk: A meta-analysis and meta-regression. Gene 2013, 531, 426–434. [Google Scholar] [CrossRef] [PubMed]
- Márquez, L.J.; Pacheco, A.; Valdés, P.; Salazar, L.A. Association between CAPN10 UCSNP-43 gene polymorphism and polycystic ovary syndrome in Chilean women. Clin. Chim. Acta Int. J. Clin. Chem. 2008, 398, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Wiltgen, D.; Furtado, L.; Kohek, M.B.F.; Spritzer, P.M. CAPN10UCSNP-43, UCSNP-19 and UCSNP-63 polymorphisms and metabolic syndrome in polycystic ovary syndrome. Gynecol. Endocrinol. 2007, 23, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, A.; Abril, E.; Roca, A.; Aragón, M.J.; Figueroa, M.J.; Velarde, P.; Ruiz, R.; Fayez, O.; Galán, J.J.; Herreros, J.A.; et al. Specific CAPN10 Gene Haplotypes Influence the Clinical Profile of Polycystic Ovary Patients. J. Clin. Endocrinol. Metab. 2003, 88, 5529–5536. [Google Scholar] [CrossRef] [PubMed]
- Tathapudi, S.; Erukkambattu, J.; Hasan, Q.; Addepally, U.; Kodati, V. Association of calpain 10 gene UCSNP-43 polymorphism (rs3792267) with polycystic ovarian syndrome. Int. J. Reprod. Contracept. Obstet. Gynecol. 2017, 4, 1185–1190. [Google Scholar] [CrossRef]
- Burks, D.J.; White, M.F. IRS proteins and beta-cell function. Diabetes 2001, 50 (Suppl. S1), S140–S145. [Google Scholar] [CrossRef]
- Jellema, A.; Zeegers, M.P.A.; Feskens, E.J.M.; Dagnelie, P.C.; Mensink, R.P. Gly972Arg variant in the insulin receptor substrate-1 gene and association with type 2 diabetes: A metaanalysis of 27 studies. Diabetologia 2003, 46, 990–995. [Google Scholar] [CrossRef]
- El Mkadem, S.A.; Lautier, C.; Macari, F.; Molinari, N.; Lefebvre, P.; Renard, E.; Gris, J.C.; Cros, G.; Daures, J.P.; Bringer, J.; et al. Role of allelic variants Gly972Arg of IRS-1 and Gly1057Asp of IRS-2 in moderate- to-severe insulin resistance of women with polycystic ovary syndrome. Diabetes 2001, 50, 2164–2168. [Google Scholar] [CrossRef] [PubMed]
- Sir-Petermann, T.; Pérez-Bravo, F.; Angel, B.; Maliqueo, M.; Calvillan, M.; Palomino, A. G972R polymorphism of IRS-1 in women with polycystic ovary syndrome. Diabetologia 2001, 44, 1200–1201. [Google Scholar] [PubMed]
- Dilek, S.; Ertunc, D.; Tok, E.C.; Erdal, E.M.; Aktas, A. Association of Gly972Arg variant of insulin receptor substrate-1 with metabolic features in women with polycystic ovary syndrome. Fertil. Steril. 2005, 84, 407–412. [Google Scholar] [CrossRef]
- Ioannidis, A.; Ikonomi, E.; Dimou, N.L.; Douma, L.; Bagos, P.G. Polymorphisms of the insulin receptor and the insulin receptor substrates genes in polycystic ovary syndrome: A Mendelian randomization meta-analysis. Mol. Genet. Metab. 2010, 99, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Y.; Ma, J.; Xie, X. Association of IRS-1 and IRS-2 genes polymorphisms with polycystic ovary syndrome: A meta-analysis. Endocr. J. 2012, 59, 601–609. [Google Scholar] [CrossRef]
- Corbould, A.; Zhao, H.; Mirzoeva, S.; Aird, F.; Dunaif, A. Enhanced mitogenic signaling in skeletal muscle of women with polycystic ovary syndrome. Diabetes 2006, 55, 751–759. [Google Scholar] [CrossRef]
- Lee, M.-H.; Yoon, J.-A.; Kim, H.-R.; Kim, Y.S.; Lyu, S.W.; Lee, B.S.; Song, H.; Choi, D.H. Hyperandrogenic Milieu Dysregulates the Expression of Insulin Signaling Factors and Glucose Transporters in the Endometrium of Patients With Polycystic Ovary Syndrome. Reprod. Sci. 2019, 4, 1637–1647. [Google Scholar] [CrossRef]
- Nardo, L.G.; Patchava, S.; Laing, I. Polycystic ovary syndrome: Pathophysiology, molecular aspects and clinical implications. Panminerva Med. 2008, 50, 267–278. [Google Scholar]
- Munir, I.; Yen, H.-W.; Geller, D.H.; Torbati, D.; Bierden, R.M.; Weitsman, S.R.; Agarwal, S.K.; Magoffin, D.A. Insulin Augmentation of 17α-Hydroxylase Activity Is Mediated by Phosphatidyl Inositol 3-Kinase But Not Extracellular Signal-Regulated Kinase-1/2 in Human Ovarian Theca Cells. Endocrinology 2004, 145, 175–183. [Google Scholar] [CrossRef]
- Poretsky, L.; Cataldo, N.A.; Rosenwaks, Z.; Giudice, L.C. The insulin-related ovarian regulatory system in health and disease. Endocr. Rev. 1999, 20, 535–582. [Google Scholar] [CrossRef]
- Baillargeon, J.-P.; Carpentier, A. Role of insulin in the hyperandrogenemia of lean women with polycystic ovary syndrome and normal insulin sensitivity. Fertil. Steril. 2007, 88, 886–893. [Google Scholar] [CrossRef] [PubMed]
- Junien, C.; Van Heyningen, V.; Gillett, G.T. Report of the committee on the genetic constitution of chromosome 11. Cytogenet. Cell Genet. 1991, 46, 459–554. [Google Scholar] [CrossRef]
- Paquette, J.; Giannoukakis, N.; Polychronakos, C.; Vafiadis, P.; Deal, C. The INS 5′ variable number of tandem repeats is associated with IGF2 expression in humans. J. Biol. Chem. 1998, 273, 14158–14164. [Google Scholar] [CrossRef] [PubMed]
- Waterworth, D.M.; Bennett, S.T.; Gharani, N.; McCarthy, M.I.; Hague, S.; Batty, S.; Conway, G.S.; White, D.; Todd, J.A.; Franks, S.; et al. Linkage and association of insulin gene VNTR regulatory polymorphism with polycystic ovary syndrome. Lancet 1997, 349, 986–990. [Google Scholar] [CrossRef]
- Kashima, K.; Yahata, T.; Fujita, K.; Tanaka, K. Polycystic ovary syndrome: Association of a C/T Single nucleotide polymorphism at tyrosine kinase domain of insulin receptor gene with pathogenesis among lean Japanese women. J. Reprod. Med. 2013, 58, 491–496. [Google Scholar] [PubMed]
- Diamanti-Kandarakis, E.; Papavassiliou, A.G. Molecular mechanisms of insulin resistance in polycystic ovary syndrome. Trends. Mol. Med. 2006, 12, 324–332. [Google Scholar] [CrossRef] [PubMed]
- Urbanek, M.; Woodroffe, A.; Ewens, K.G.; Diamanti-Kandarakis, E.; Legro, R.S.; Strauss, J.F., III; Dunaif, A.; Spielman, R.S. Candidate gene region for polycystic ovary syndrome on chromosome 19p13.2. J. Clin. Endocrinol. Metab. 2005, 90, 6623–6629. [Google Scholar] [CrossRef] [PubMed]
- Daghestani, M.H. Rs1799817 in INSR associates with susceptibility to polycystic ovary syndrome. J. Med. Biochem. 2020, 39, 149. [Google Scholar] [CrossRef]
- Dakshinamoorthy, J.; Jain, P.R.; Ramamoorthy, T.; Ayyappan, R.; Balasundaram, U. Association of GWAS identified INSR variants (rs2059807 & rs1799817) with polycystic ovarian syndrome in Indian women. Int. J. Biol. Macromol. 2020, 144, 663–670. [Google Scholar]
- Tian, Y.; Li, J.; Su, S.; Cao, Y.; Wang, Z.; Zhao, S.; Zhao, H. PCOS-GWAS Susceptibility Variants in THADA, INSR, TOX3, and DENND1A Are Associated With Metabolic Syndrome or Insulin Resistance in Women with PCOS. Front. Endocrinol. 2020, 11, 274. [Google Scholar] [CrossRef]
- Shaaban, Z.; Khoradmehr, A.; Amiri-Yekta, A.; Nowzari, F.; Shirazi, M.R.J.; Tamadon, A. Pathophysiologic Mechanisms of Insulin Secretion and Signaling-Related Genes in Etiology of Polycystic Ovary Syndrome. Genet. Res. 2021, 2021, 7781823. [Google Scholar] [CrossRef] [PubMed]
- Gottlieb, B.; Beitel, L.K.; Nadarajah, A.; Paliouras, M.; Trifiro, M. The androgen receptor gene mutations database: 2012 update. Hum. Mutat. 2012, 33, 887–894. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.; Polson, D.; Franks, S. Prevalence of polycystic ovaries in women with anovulation and idiopathic hirsutism. Br. Med. J. (Clin. Res. Ed.) 1986, 293, 355–359. [Google Scholar] [CrossRef] [PubMed]
- Xita, N.; Tsatsoulis, A. Fetal programming of polycystic ovary syndrome by androgen excess: Evidence from experimental, clinical, and genetic association studies. J. Clin. Endocrinol. Metab. 2006, 91, 1660–1666. [Google Scholar] [CrossRef]
- Schüring, A.N.; Welp, A.; Gromoll, J.; Zitzmann, M.; Sonntag, B.; Nieschlag, E.; Greb, R.R.; Kiesel, L. Role of the CAG Repeat Polymorphism of the Androgen Receptor Gene in Polycystic Ovary Syndrome (PCOS). Exp. Clin. Endocrinol. Diabetes 2012, 120, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xia, Y.; Che, Y.; Zhang, X.; Zhang, C.; Cao, Y.; Wang, W.; Xu, P.; Wu, X.; Yi, L.; et al. Polymorphic CAG repeat in the androgen receptor gene in polycystic ovary syndrome patients. Mol. Med. Rep. 2012, 5, 1330–1334. [Google Scholar] [CrossRef][Green Version]
- Lin, L.H.; Baracat, M.C.; Maciel, G.A.; Soares, J.M., Jr.; Baracat, E.C. Androgen receptor gene polymorphism and polycystic ovary syndrome. Int. J. Gynecol. Obstet. 2013, 120, 115–118. [Google Scholar] [CrossRef]
- Shah, N.A.; Antoine, H.J.; Pall, M.; Taylor, K.D.; Azziz, R.; Goodarzi, M.O. Association of Androgen Receptor CAG Repeat Polymorphism and Polycystic Ovary Syndrome. J. Clin. Endocrinol. Metab. 2008, 93, 1939–1945. [Google Scholar] [CrossRef]
- Mifsud, A.; Ramirez, S.; Yong, E. Androgen receptor gene CAG trinucleotide repeats in anovulatory infertility and polycystic ovaries. J. Clin. Endocrinol. Metab. 2000, 85, 3484–3488. [Google Scholar] [CrossRef]
- Rajender, S.; Carlus, S.J.; Bansal, S.K.; Negi, M.P.S.; Sadasivam, N.; Sadasivam, M.N.; Thangaraj, K. Androgen Receptor CAG Repeats Length Polymorphism and the Risk of Polycystic Ovarian Syndrome (PCOS). PLoS ONE 2013, 8, e75709. [Google Scholar] [CrossRef]
- Ferk, P.; Perme, M.P.; Teran, N.; Gersak, K. Androgen receptor gene (CAG) n polymorphism in patients with polycystic ovary syndrome. Fertil. Steril. 2008, 90, 860–863. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.J.; Choung, S.H.; Choi, Y.M.; Yoon, S.H.; Kim, S.H.; Moon, S.Y. Androgen receptor gene CAG repeat polymorphism in women with polycystic ovary syndrome. Fertil. Steril. 2008, 90, 2318–2323. [Google Scholar] [CrossRef] [PubMed]
- Skrgatic, L.; Baldani, D.P.; Cerne, J.; Ferk, P.; Gersak, K. CAG repeat polymorphism in androgen receptor gene is not directly associated with polycystic ovary syndrome but influences serum testosterone levels. J. Steroid. Biochem. Mol. Biol. 2012, 128, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Hickey, T.; Chandy, A.; Norman, R.J. The androgen receptor CAG repeat polymorphism and X-chromosome inactivation in Australian Caucasian women with infertility related to polycystic ovary syndrome. J. Clin. Endocrinol. Metab. 2002, 87, 161–165. [Google Scholar] [CrossRef] [PubMed]
- Otsuka, F.; Yao, Z.; Lee, T.-H.; Yamamoto, S.; Erickson, G.F.; Shimasaki, S. Bone morphogenetic protein-15: Identification of target cells and biological functions. J. Biol. Chem. 2000, 275, 39523–39528. [Google Scholar] [CrossRef]
- Otsuka, F.; Shimasaki, S. A novel function of bone morphogenetic protein-15 in the pituitary: Selective synthesis and secretion of FSH by gonadotropes. Endocrinology 2002, 143, 4938–4941. [Google Scholar] [CrossRef]
- Calvo, R.M.; Asuncion, M.; Sancho, J.; San Millan, J.L.; Escobar-Morreale, H.F. The role of the CAG repeat polymorphism in the androgen receptor gene and of skewed X-chromosome inactivation, in the pathogenesis of hirsutism. J. Clin. Endocrinol. Metab. 2000, 85, 1735–1740. [Google Scholar]
- Chaudhary, H.; Patel, J.; Jain, N.K.; Joshi, R. The role of polymorphism in various potential genes on polycystic ovary syndrome susceptibility and pathogenesis. J. Ovarian Res. 2021, 14, 125. [Google Scholar] [CrossRef]
- Bérubé, D.; Séralini, G.E.; Gagné, R.; Hammond, G.L. Localization of the human sex hormone-binding globulin gene (SHBG) to the short arm of chromosome 17 (17p12→p13). Cytogenet. Genome Res. 1990, 54, 65–67. [Google Scholar]
- Hammond, G.L. Molecular properties of corticosteroid binding globulin and the sex-steroid binding proteins. Endocr. Rev. 1990, 11, 65–79. [Google Scholar] [CrossRef]
- Edmunds, S.E.J.; Stubbs, A.P.; Santos, A.A.; Wilkinson, M.L. Estrogen and androgen regulation of sex hormone binding globulin secretion by a human liver cell line. J. Steroid. Biochem. Mol. Biol. 1990, 37, 733–739. [Google Scholar] [CrossRef]
- Nestler, J.E.; Powers, L.P.; Matt, D.W.; Steingold, K.A.; Plymate, S.R.; Rittmaster, R.S.; Clore, J.N.; Blackard, W.G. A Direct Effect of Hyperinsulinemia on Serum Sex Hormone-Binding Globulin Levels in Obese Women with the Polycystic Ovary Syndrome. J. Clin. Endocrinol. Metab. 1991, 72, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Plymate, S.R.; Matej, L.A.; Jones, R.E.; Friedl, K.E. Inhibition of sex hormone-binding globulin production in the human hepatoma (Hep G2) cell line by insulin and prolactin. J. Clin. Endocrinol. Metab. 1988, 67, 460–464. [Google Scholar] [CrossRef] [PubMed]
- Wickham, E.P.; Ewens, K.G.; Legro, R.S.; Dunaif, A.; Nestler, J.E.; Strauss, J.F. Polymorphisms in the SHBG gene influence serum SHBG levels in women with polycystic ovary syndrome. J. Clin. Endocrinol. Metab. 2011, 96, E719–E727. [Google Scholar] [CrossRef]
- Chen, C.; Smothers, J.; Lange, A.; Nestler, J.E.; Strauss Iii, J.F.; Wickham Iii, E.P. Sex hormone-binding globulin genetic variation: Associations with type 2 diabetes mellitus and polycystic ovarysyndrome. Minerva Endocrinol. 2010, 35, 271–280. [Google Scholar]
- Franks, S.; Stark, J.; Hardy, K. Follicle dynamics and anovulation in polycystic ovary syndrome. Hum. Reprod. Update 2008, 14, 539. [Google Scholar] [CrossRef]
- Pigny, P.; Merlen, E.; Robert, Y.; Cortet-Rudelli, C.; Decanter, C.; Jonard, S.; Dewailly, D. Elevated Serum Level of Anti-Mullerian Hormone in Patients with Polycystic Ovary Syndrome: Relationship to the Ovarian Follicle Excess and to the Follicular Arrest. J. Clin. Endocrinol. Metab. 2003, 88, 5957–5962. [Google Scholar] [CrossRef]
- Furui, K.; Suganuma, N.; Tsukahara, S.; Asada, Y.; Kikkawa, F.; Tanaka, M.; Ozawa, T.; Tomoda, Y. Identification of two point mutations in the gene coding luteinizing hormone (LH) beta-subunit, associated with immunologically anomalous LH variants. J. Clin. Endocrinol. Metab. 1994, 78, 107–113. [Google Scholar] [CrossRef]
- Nilsson, C.; Pettersson, K.; Millar, R.P.; Coerver, K.A.; Matzuk, M.M.; Huhtaniemi, I.T. Worldwide frequency of a common genetic variant of luteinizing hormone: An international collaborative research. Fertil. Steril. 1997, 67, 998–1004. [Google Scholar] [CrossRef]
- Roy, A.C.; Liao, W.-X.; Chen, Y.; Arulkumaran, S.; Ratnam, S.S. Identification of seven novel mutations in LH ?-Subunit gene by SSCP. Mol. Cell. Biochem. 1996, 165, 151–153. [Google Scholar] [CrossRef]
- Gromoll, J.; Simoni, M. Genetic complexity of FSH receptor function. Trends Endocrinol. Metab. 2005, 16, 368–373. [Google Scholar] [CrossRef] [PubMed]
- Baban, A.S.S.; Korsheed, S.H.; Al Hayawi, A.Y. The FSHR polymorphisms association with polycystic ovary syndrome in women of Erbil, Kurdistan in North of Iraq. Ibn AL-Haitham J. Pure Appl. Sci. 2018, 262, 257–272. [Google Scholar] [CrossRef]
- Cate, R.; Mattaliano, R.; Hession, C.; Tizard, R.; Farber, N.; Cheung, A.; Ninfa, E.; Frey, A.; Gash, D.; Chow, E.; et al. Isolation of the bovine and human genes for müllerian inhibiting substance and expression of the human gene in animal cells. Cell 1986, 45, 685–698. [Google Scholar] [CrossRef]
- Gorsic, L.K.; Kosova, G.; Werstein, B.; Sisk, R.; Legro, R.; Hayes, M.G.; Teixeira, J.; Dunaif, A.; Urbanek, M. Pathogenic Anti-Müllerian Hormone Variants in Polycystic Ovary Syndrome. J. Clin. Endocrinol. Metab. 2017, 102, 2862–2872. [Google Scholar] [CrossRef] [PubMed]
- Lidija, K.; Gorsic, M.D.; Richard, S.; Legro, M. Geoffrey Hayes, and Margrit Urbanek, Functional Genetic Variation in the AMH Pathway in PCOS. J. Clin. Endocrinol. Metab. 2019, 104, 2855–2874. [Google Scholar]
- Teixeira, J.; Fynn-Thompson, E.; Payne, A.H.; Donahoe, P.K. Müllerian inhibiting substance regulates androgen synthesis at the transcriptional level. Endocrinology 1999, 140, 4732–4738. [Google Scholar] [CrossRef]
- Teixeira, J.; Maheswaran, S.; Donahoe, P.K. Müllerian inhibiting substance: An instructive developmental hormone with diagnostic and possible therapeutic applications. Endocr. Rev. 2001, 22, 657–674. [Google Scholar] [CrossRef]
- Park, J.H.; Maclaughlin, D.T.; Teixeira, J.M. The rate of in vitro maturation of primary follicles from adult mice and the quality of oocytes is improved in the absence of anti-mullerian hormone. Reprod. Sci. 2011, 18, 334–341. [Google Scholar] [CrossRef]
- Maciel, G.A.; Baracat, E.C.; Benda, J.A.; Markham, S.M.; Hensinger, K.; Chang, R.J.; Erickson, G.F. Stockpiling of transitional and classic primary follicles in ovaries of women with polycystic ovary syndrome. J. Clin. Endocrinol. Metab. 2004, 89, 5321–5327. [Google Scholar] [CrossRef]
- Belville, C.; Jamin, S.P.; Picard, J.Y.; Josso, N.; di Clemente, N. Role of type I receptors for anti-Müllerian hormone in the SMAT-1 Sertoli cell line. Oncogene 2005, 24, 4984–4992. [Google Scholar] [CrossRef]
- Racine, C.; Rey, R.; Forest, M.G.; Louis, F.; Ferré, A.; Huhtaniemi, I.; Josso, N.; di Clemente, N. Receptors for anti-Müllerian hormone on Leydig cells are responsible for its effects on steroidogenesis and cell differentiation. Proc. Natl. Acad. Sci. USA 1998, 95, 594–599. [Google Scholar] [CrossRef] [PubMed]
- Fynn-Thompson, E.; Cheng, H.; Teixeira, J. Inhibition of steroidogenesis in Leydig cells by Müllerian-inhibiting substance. Mol. Cell. Endocrinol. 2003, 211, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Grossman, M.P.; Nakajima, S.T.; Fallat, M.E.; Siow, Y. Müllerianinhibiting substance inhibits cytochrome P450 aromatase activity in human granulosa lutein cell culture. Fertil. Steril. 2008, 89 (Suppl. 5), 1364–1370. [Google Scholar] [CrossRef] [PubMed]
- Sèdes, L.; Leclerc, A.; Moindjie, H.; Cate, R.L.; Picard, J.Y.; di Clemente, N.; Jamin, S.P. Anti-Müllerian hormone recruits BMPR-IA in immature granulosa cells. PLoS ONE 2013, 8, e81551. [Google Scholar] [CrossRef]
- Urbanek, M.; Legro, R.S.; Driscoll, D.A.; Azziz, R.; Ehrmann, D.A.; Norman, R.J.; Strauss, J.F., III; Spielman, R.S.; Dunaif, A. Thirty-seven candidate genes for polycystic ovary syndrome: Strongest evidence for linkage is with follistatin. Proc. Natl. Acad. Sci. USA 1999, 96, 8573–8578. [Google Scholar] [CrossRef]
- Jakimiuk, A.J.; Weitsman, S.R.; Magoffin, D.A. 5α-Reductase activity in women with polycystic ovary syndrome. J. Clin. Endocrinol. Metab. 1999, 84, 2414–2418. [Google Scholar]
- Goodarzi, M.O.; Shah, N.A.; Antoine, H.J.; Pall, M.; Guo, X.; Azziz, R. Variants in the 5α-reductase type 1 and type 2 genes are associated with polycystic ovary syndrome and the severity of hirsutism in affected women. J. Clin. Endocrinol. Metab. 2006, 91, 4085–4091. [Google Scholar] [CrossRef]
- Wojciechowski, P.; GIANT Consortium; Lipowska, A.; Rys, P.; Ewens, K.G.; Franks, S.; Tan, S.; Lerchbaum, E.; Vcelak, J.; Attaoua, R.; et al. Impact of FTO genotypes on BMI and weight in polycystic ovary syndrome: A systematic review and meta-analysis. Diabetologia 2012, 55, 2636–2645. [Google Scholar] [CrossRef]
- Rizwan, S.; Ghazanvi, S.; Rasheed, N.; Mi, U. Association of FTO common RS9939609 polymorphism with obesity and association of FTO common RS9939609 polymorphism with obesity and polycystic ovarian syndrome in Pakistani women. J. Med. Res. Biol. Stud. 2018, 1, 101. [Google Scholar]
- Talmud, P.J.; Cooper, J.A.; Morris, R.W.; Dudbridge, F.; Shah, T.; Engmann, J.; Dale, C.; White, J.; McLachlan, S.; Zabaneh, D.; et al. Sixty-Five Common Genetic Variants and Prediction of Type 2 Diabetes. Diabetes 2014, 64, 1830–1840. [Google Scholar] [CrossRef]
- Krarup, N.; Borglykke, A.; Allin, K.; Sandholt, C.; Justesen, J.M.; Andersson, E.; Grarup, N.; Jørgensen, T.; Pedersen, O.; Hansen, T. A genetic risk score of 45 coronary artery disease risk variants associates with increased risk of myocardial infarction in 6041 Danish individuals. Atherosclerosis 2015, 240, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Li, Z.; Ai, F.; Chen, J.; Xing, Q.; Zhou, P.; Wei, Z.; Shi, Y.; He, X.-J.; Cao, Y. Systematic Evaluation of Genetic Variants for Polycystic Ovary Syndrome in a Chinese Population. PLoS ONE 2015, 10, e0140695. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Oh, J.Y.; Sung, Y.A.; Chung, H.W. A genetic risk score is associated with polycystic ovary syndrome–related traits. Hum. Reprod. 2016, 31, 209. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Brower, M.A.; Jones, M.R.; Rotter, J.I.; Krauss, R.M.; Legro, R.S.; Azziz, R.; Goodarzi, M.O. Further investigation in europeans of susceptibility variants for polycystic ovary syndrome discovered in genome-wide association studies of Chinese individuals. J. Clin. Endocrinol. Metab. 2015, 100, E182–E186. [Google Scholar] [CrossRef] [PubMed]
- Zeind, C.S.; Carvalho, M.G. Applied Therapeutics: The Clinical Use of Drugs; Wolters Kluwer Health: Philadelphia, PA, USA, 2017. [Google Scholar]
- Liu, H.-Y.; Liu, J.-Q.; Mai, Z.-X.; Zeng, Y.-T. A Subpathway-Based Method of Drug Reposition for Polycystic Ovary Syndrome. Reprod. Sci. 2014, 22, 423–430. [Google Scholar] [CrossRef]
- Zhang, X.; Zheng, Y.; Guo, Y.; Lai, Z. The Effect of Low Carbohydrate Diet on Polycystic Ovary Syndrome: A Meta-Analysis of Randomized Controlled Trials. Int. J. Endocrinol. 2019, 2019, 4386401. [Google Scholar] [CrossRef]
- Brennan, L.; Teede, H.; Skouteris, H.; Linardon, J.; Hill, B.; Moran, L. Lifestyle and Behavioral Management of Polycystic Ovary Syndrome. J. Women’s Health 2017, 26, 836–848. [Google Scholar] [CrossRef]
- Faghfoori, Z.; Fazelian, S.; Shadnoush, M.; Goodarzi, R. Nutritional management in women with polycystic ovary syndrome: A review study. Diabetes Metab. Syndr. Clin. Res. Rev. 2017, 11, S429–S432. [Google Scholar] [CrossRef]
- Hakimi, O.; Cameron, L.-C. Effect of Exercise on Ovulation: A Systematic Review. Sports Med. 2016, 47, 1555–1567. [Google Scholar] [CrossRef]
- Li, Y.; Zheng, Q.; Sun, D.; Cui, X.; Chen, S.; Bulbul, A.; Liu, S.; Yan, Q. Dehydroepiandrosterone stimulates inflammation and impairs ovarian functions of polycystic ovary syndrome. J. Cell. Physiol. 2018, 234, 7435–7447. [Google Scholar] [CrossRef]
- Rudnicka, E.; Suchta, K.; Grymowicz, M.; Calik-Ksepka, A.; Smolarczyk, K.; Duszewska, A.; Smolarczyk, R.; Meczekalski, B. Chronic Low Grade Inflammation in Pathogenesis of PCOS. Int. J. Mol. Sci. 2021, 22, 3789. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.-Y.; Feng, J.-X.; Li, J.-L.; Liu, F.-Y.; Xie, L.-Z.; Luo, S.-J.; Han, F.-J. The Complementary and Alternative Medicine for Polycystic Ovary Syndrome: A Review of Clinical Application and Mechanism. Evid.-Based Complement. Altern. Med. 2021, 2021, 5555315. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; Jin, B.; Pan, Y.; Han, Y.; You, T.; Zhang, Z.; Qu, Y.; Liu, S.; Zhang, Y. The Effects of Traditional Chinese Medicine-Associated Complementary and Alternative Medicine on Women with Polycystic Ovary Syndrome. Evid.-Based Complement. Altern. Med. 2021, 2021, 6619597. [Google Scholar] [CrossRef] [PubMed]
- Raja-Khan, N.; Stener-Victorin, E.; Wu, X.; Legro, R.S. The physiological basis of complementary and alternative medicines for polycystic ovary syndrome. Am. J. Physiol. Metab. 2011, 301, E1–E10. [Google Scholar] [CrossRef]
- Zhang, Y.; Guo, X.; Ma, S.; Ma, H.; Li, H.; Wang, Y.; Qin, Z.; Wu, X.; Han, Y.; Han, Y. The Treatment with Complementary and Alternative Traditional Chinese Medicine for Menstrual Disorders with Polycystic Ovary Syndrome. Evid.-Based Complement. Altern. Med. 2021, 2021, 6678398. [Google Scholar] [CrossRef]
- Shirvani-Rad, S.; Tabatabaei-Malazy, O.; Mohseni, S.; Hasani-Ranjbar, S.; Soroush, A.-R.; Hoseini-Tavassol, Z.; Ejtahed, H.-S.; Larijani, B. Probiotics as a Complementary Therapy for Management of Obesity: A Systematic Review. Evid.-Based Complement. Altern. Med. 2021, 2021, 6688450. [Google Scholar] [CrossRef]
- Li, Y.; Peng, C.; Cao, G.; Li, W.; Hou, L. Tai chi for overweight/obese adolescent and young women with polycystic ovary syndrome: Study protocol for a randomized controlled trial. Trials 2018, 19, 512. [Google Scholar] [CrossRef]
- Mohseni, M.; Eghbali, M.; Bahrami, H.; Dastaran, F.; Amini, L. Yoga Effects on Anthropometric Indices and Polycystic Ovary Syndrome Symptoms in Women Undergoing Infertility Treatment: A Randomized Controlled Clinical Trial. Evid.-Based Complement. Altern. Med. 2021, 2021, 5564824. [Google Scholar] [CrossRef]
- Legro, R.S.; Duguech, L.M.M. Pharmacologic Treatment of Polycystic Ovary Syndrome: Alternate and Future Paths. Semin. Reprod. Med. 2017, 35, 326–343. [Google Scholar] [CrossRef]
- Ortega, I.; Villanueva, J.A.; Wong, D.H.; Cress, A.B.; Sokalska, A.; Stanley, S.D.; Duleba, A.J. Resveratrol potentiates effects of simvastatin on inhibition of rat ovarian theca-interstitial cells steroidogenesis. J. Ovarian Res. 2014, 7, 21. [Google Scholar] [CrossRef]
- Crandall, J.P.; Oram, V.; Trandafirescu, G.; Reid, M.; Kishore, P.; Hawkins, M.; Cohen, H.W.; Barzilai, N. Pilot Study of Resveratrol in Older Adults With Impaired Glucose Tolerance. J. Gerontol. Ser. A Boil. Sci. Med. Sci. 2012, 67, 1307–1312. [Google Scholar] [CrossRef] [PubMed]
- Rondanelli, M.; Infantino, V.; Riva, A.; Petrangolini, G.; Faliva, M.A.; Peroni, G.; Naso, M.; Nichetti, M.; Spadaccini, D.; Gasparri, C.; et al. Polycystic ovary syndrome management: A review of the possible amazing role of berberine. Arch. Gynecol. Obstet. 2020, 301, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Naka, K.K.; Kalantaridou, S.N.; Kravariti, M.; Bechlioulis, A.; Kazakos, N.; Calis, K.A.; Makrigiannakis, A.; Katsouras, C.S.; Chrousos, G.P.; Tsatsoulis, A.; et al. Effect of the insulin sensitizers metformin and pioglitazone on endothelial function in young women with polycystic ovary syndrome: A prospective randomized study. Fertil. Steril. 2011, 95, 203–209. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| S.N | Diagnosis Criteria | Symptoms | Recommendations |
|---|---|---|---|
| 1. | National Institute of Health (1990) |
| Both criteria are required |
| 2. | Rotterdam Consensus (2003) |
| Two of three criteria required |
| 3. | Androgen Excess Society (2006) |
| Both criteria required |
| Phenotype | Hyperandrogenism | Chronic Anovulation | Polycystic Ovaries Morphology | NIH 1990 | Rotterdam | AE-PCOS 2006 |
|---|---|---|---|---|---|---|
| Phenotype A | Yes | Yes | Yes | - | - | - |
| Phenotype B | Yes | Yes | No | - | - | - |
| Phenotype C | Yes | No | Yes | - | - | |
| Phenotype D | No | Yes | Yes | - |
| S.N | Diagnostic Criteria | Gene Locus | SNPs | Nearest Gene | Study |
|---|---|---|---|---|---|
| 1. | Rotterdam | 2p16.3 | rs13405728 | LHCGHR STON1-GTF2A1L | Chen et al., 2011 [66] |
| 2p21 | rs12468394 | ||||
| rs13429458 | THADA | ||||
| rs12478601 | |||||
| 9q33.3 | rs10818854 | DENND1A | |||
| rs10986105 | |||||
| rs24779106 | |||||
| 2. | NIH | 8p32.1 | rs804279 | NEIL2, GATA4 | Hayes et al., 2015 [63] |
| 9q22.32 | rs10993397 | C9orf3 | |||
| 11p14.1 | rs11031006 | ARL14EP, FSHB | |||
| 3 | Rotterdam | 12p12.2 | rs10841843 | GYS2 | Hwang et al., 2012 [68] |
| rs6487237 | |||||
| rs7485509 | |||||
| 4 | NIH | 2q.34 | rs1351592 | ERBB4 | Day et at., 2015 [67] |
| 11q22.1 | rs11225154 | YAP1 | |||
| 2q21 | rs7563201 | THADA | |||
| 11p14.1 | rs11031006 | FSHB | |||
| 5q31.1 | rs13164856 | RAD50 | |||
| 12q21.2 | rs1275468 | KRR1 | |||
| 5. | Rotterdam | 8q24.2 | rs10505648 | KHDRBS3 LICE02055 | Lee et al., 2012 [69] |
| S.N | Different Gene Categories Involved in PCOS | Genes under Categories |
|---|---|---|
| 1. | Genes involved in ovarian and adrenal steroidogenesis |
|
| 2. | Epigenetics of PCOS |
|
| 3. | Gene involved in insulin action and secretion |
|
| 4. | Gene involved in steroid hormone effect |
|
| 5. | Gene involved in gonadotropin |
|
| 6. | Other genes |
|
| S.N | Gene | Type of Polymorphism | Genetic Marker | Physiological Function | Type of Study with Study Population | Reference |
|---|---|---|---|---|---|---|
| 1 | C A P N 10 | UCSNP-43 | UCSNP-43, 19 and 63 | Calcium medium, intracellular signaling, insulin secretion | Case control study on Chilean women | [117] |
| 2 | UCSNP43 | UCSNP-43, 19 and 63 | Cross sectional study on Brazilian women | [118] | ||
| 3 | UCSNP44 | CAPN10 haplotypes | Haplotype phenotype correlation study on Spanish women | [119] | ||
| 4 | UCSNP-19,45, and 63 | SNPs | Meta-analysis on Asian women | [116] | ||
| 5 | UCSNP-19, 43, and 44 | UCSNP-19, 43, 44 and 63 | Cross sectional study on Spanish women | [112] | ||
| 6 | UCSNP-19,63 | UCSNP-19, 63, 43 and 44 | Meta-analysis Different population | [115] | ||
| 7 | - | UCSNP-43, rs 3792267 | Case control on Indian women | [120] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nautiyal, H.; Imam, S.S.; Alshehri, S.; Ghoneim, M.M.; Afzal, M.; Alzarea, S.I.; Güven, E.; Al-Abbasi, F.A.; Kazmi, I. Polycystic Ovarian Syndrome: A Complex Disease with a Genetics Approach. Biomedicines 2022, 10, 540. https://doi.org/10.3390/biomedicines10030540
Nautiyal H, Imam SS, Alshehri S, Ghoneim MM, Afzal M, Alzarea SI, Güven E, Al-Abbasi FA, Kazmi I. Polycystic Ovarian Syndrome: A Complex Disease with a Genetics Approach. Biomedicines. 2022; 10(3):540. https://doi.org/10.3390/biomedicines10030540
Chicago/Turabian StyleNautiyal, Himani, Syed Sarim Imam, Sultan Alshehri, Mohammed M. Ghoneim, Muhammad Afzal, Sami I. Alzarea, Emine Güven, Fahad A. Al-Abbasi, and Imran Kazmi. 2022. "Polycystic Ovarian Syndrome: A Complex Disease with a Genetics Approach" Biomedicines 10, no. 3: 540. https://doi.org/10.3390/biomedicines10030540
APA StyleNautiyal, H., Imam, S. S., Alshehri, S., Ghoneim, M. M., Afzal, M., Alzarea, S. I., Güven, E., Al-Abbasi, F. A., & Kazmi, I. (2022). Polycystic Ovarian Syndrome: A Complex Disease with a Genetics Approach. Biomedicines, 10(3), 540. https://doi.org/10.3390/biomedicines10030540