
Biocides as Biomedicines against Foodborne Pathogenic Bacteria


 , and
, and 
Abstract
:1. Introduction
2. Biocides
2.1. Metal-Based Biocides
2.2. Aldehydes, Alcohols, Phenols, and Bisphenols
2.3. Halogens, Peroxides, and Organic Acids
2.4. Quaternary Ammonium Compounds (QACs)
2.5. Phytochemicals
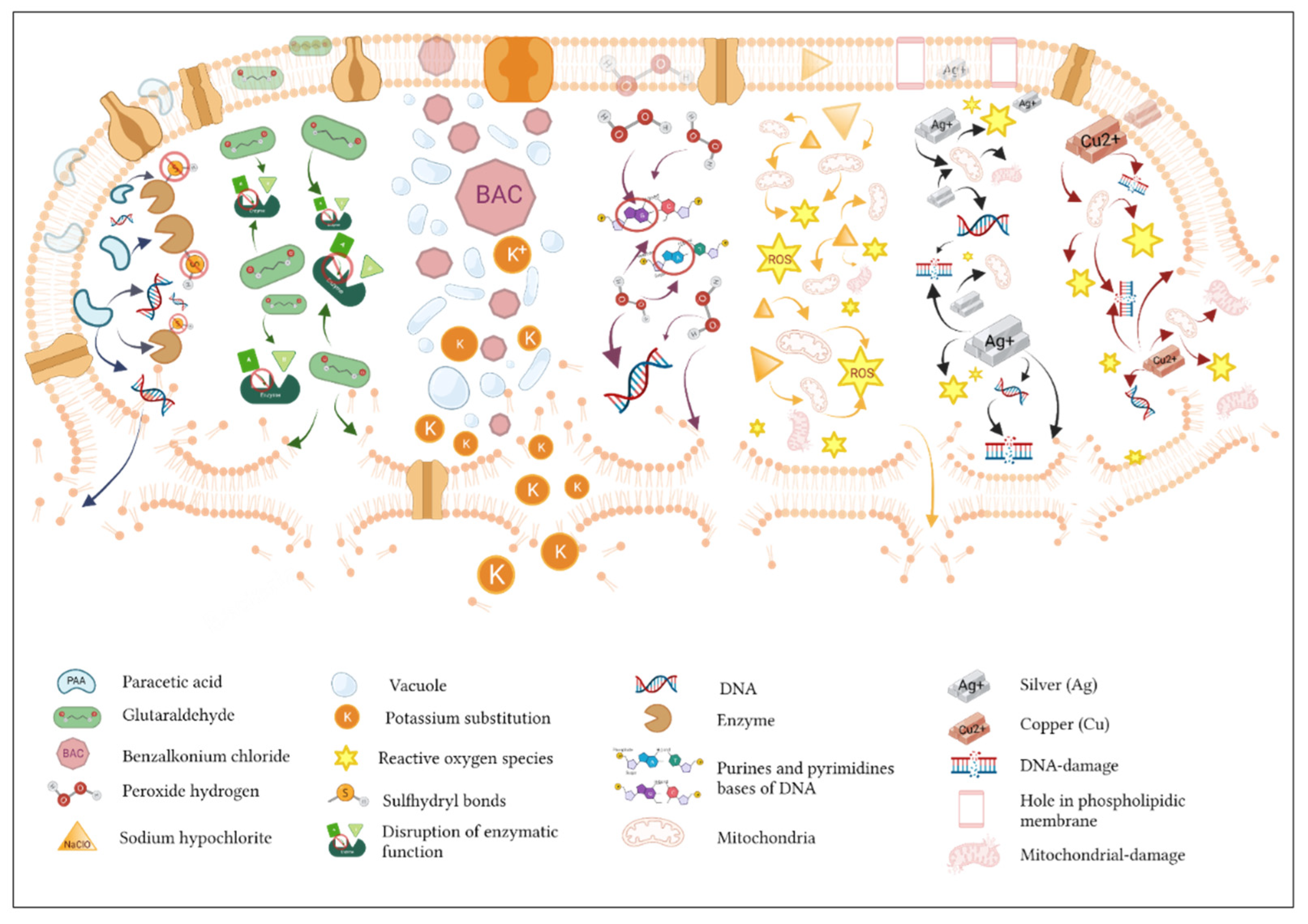
3. Biological Mechanisms of Action of Biocides
3.1. Release-Killing Biocides and Contact-Killing Biocides
3.2. Biocide Efficacy
3.3. Inducing the Coresistance and Tolerance of Microorganisms through the Use of Biocides
4. The Effect of Biocides against Foodborne Pathogenic Bacteria
4.1. Escherichia spp.

{kind=link}
| Biocide | Strain | Concentration | Mechanism/Notes | Gene/Protein | Ref. |
|---|---|---|---|---|---|
| MBQ (magnetic biochar/quaternary phosphonium salt) | E. coli ATCC 25922 | MBC 20 mg/L | Cell wall and membrane penetration; induced vacuolization, loss of cell integrity/permeability, leakage of intracellular components, oxidative stress. | n.i. | [73] |
| Cellulose foam paper with BTCA and GA | E. coli MTCC 40 | MIC 1 g/L | BTCA—crosslinking agent. Reduced pH and growth. GA—cross-binding with cell wall amines interfered with the transport and enzymatic activities and bactericidal effect. | n.i. | [13] |
| Polyvinylidene fluoride membranes with guanidine backbone or sulphonium backbone | E. coli ATCC 25922 | Bacteria lost structural integrity caused by the existing electrostatic interactions, which led to the leakage of intracellular components and caused their death. | n.i. | [12] | |
| Sodium hypochlorite; chlorine dioxide; neutral electrolyzed oxidizing water; sodium dichloroisocyanurate | E. coli CECT 434 | MBC 80 and 100 ppm—planktonic cultures; 50 ppm—biofilms | ROS production in the case of SH and NEOW. | n.i. | [38] |
| KFU-127 (pyridoxine-based quaternary ammonium derivatives of terbinafine 127) | E. coli MG1655 | MIC 8 µg/mL MBC ×16 | Cell membrane damage and membrane potential changes; inhibitory property on pyroxidal-dependent enzymes. | n.i. | [11] |
| Peptide biocides (45 analogs of antimicrobial peptide indolicidin) | E. coli M17; E. coli ATCC 252934 | MIC 0.9–1.8 µg/mL | Pore-forming agents; the introduction of groups Cl, NO2, F into the aromatic ring in the structure of biocides leads to increased antimicrobial activity; analogs with higher hydrophobicity have the possibility of breaking the cell membrane. | n.i. | [71] |
| PAA (peracetic acid) PFA (performic acid) | AmpR E. coli | MIC 80 and 60 mg min/L 10 and 15 mg min/L | Oxidation of sulfur and sulfhydryl bonds in proteins and enzymes; disruption/dislodging the cell walls and modifying the cytoplasmic membrane of lipoproteins, blocked enzymatic and transport processes; formation of hydroxyl peroxide free radicals. | n.i. | [74] |
| PMA (permaleic acid) | E. coli ATCC 25922 | MIC 40 mg/L (50%) 100 mg/L (100%) | Damage and disruption of cell membrane activity. | n.i. | [75] |
| ECASs (1%, 5%, 10%) (electrochemically activated solutions) | E. coli from the water disruption network of NUST | Oxidizing agent; induced damage to the cell membrane; Cl in ECASs increased cell permeability and disrupted protein synthesis; degradation of functional groups (ClO2, H2O2, ozone) over time weakens oxidative stress. | n.i. | [72] | |
| AuNSps (nanosphere) AuNSts (nanostars) AuNCs (nanocubes) | E. coli from Industrial Culture Collection, China | MIC 80 µL; MBCs AuNSps—0.02 and 0.04 µg; AuNCs—0.2 and 0.4 µg | Visible surface damage, disturbance and cell loss by disruption of membrane-bound components, loss of flagella, loss of cell integrity, leakage of cell contents into the environment, death. | n.i. | [76] |
| Ag-iNPs | Homogenized microbial solution of E. coli commercial lyophilized pellet (ATCC) | Affected phospholipids, cytoplasm proteins (GADPH). Created holes on the outer membrane (OM), increased the permeability of the membrane, led to disruption of the breathing cycle leading to its lysis; electrostatic interaction, leading to disruption of the integrity of the OM and activation of OMPLA lipolytic enzymes; ROS formation | ↑PldA ↑cueO ↑copA ↑cusR | [77,78] | |
| AgNO3 | ↓ZntA gene ↓CopA gene ↓CueO gene | [77] | |||
| Cu/SHfNP | E. coli SE4 isolated from hospital wastewaters | MIC 2 mM (98%); 1 mM (91.9%)—reduced biofilm formation | Loss of cellular components of cells; reduced the level of attachment of biofilm cells; disrupted nanowire formation. | n.i. | [79] |
| BDCA-RNM (combination of benzophenone tetracarboxylic dianhydride and chlorogenic acid membranes) | E. coli O157:H7 | MBC 10 µL | ROS production; cellular deformation and surface collapse; lysed and disrupted the bacterial cell walls and membranes; leakage of nucleic acids and proteins. | n.i. | [80] |
4.2. Pseudomonas spp.
| Biocide | Strain | Concentration | Mechanism/Notes | Gene/Protein | Ref. |
|---|---|---|---|---|---|
| Cellulose foam filter paper with GA and BTCA | P. aeruginosa NCDC 105 | MIC 1 g/L | BTCA—crosslinking agent. Possible mechanism—its acidic nature reduces the pH of the medium, which inhibits bacterial growth; Glutaraldehyde—cross-binding of the molecule with the amines of bacterial cells, interfering in the transport and enzymatic activities, which disrupts the work of the main functions, causing bactericidal effect. | n.i. | [13] |
| AuNSps AuNSts AuNCs | P. aeruginosa obtained from the China Center for Industrial Culture Collection | MIC: 80 µL; MBCs: AuNSps—0.02 and 0.04 µg; AuNCs—0.2 and 0.4 µg | Surface damage, disturbance, cell loss by disruption of membrane-bound components, loss of flagella, leakage of cell contents into the environment. | n.i. | [76] |
| KFU-127 | P. aeruginosa ATCC 27853 | MIC/MBC 64 µg/mL | Cell membrane damage and changes; inhibitory property on pyroxidal-dependent enzymes. | n.i. | [11] |
| CHX—1% BKC—1% Kohrsolin extra SEPTI-Turbo | P. aeruginosa NCTC 10662 and P. aeruginosa isolates collected from different hospitals in Hamadan city, Iran | MICs: CHX—8–128 µg/mL; BKC—8–64 µg/mL; Kohrsolin extra—8–32 µg/mL; SEPTI-Turbo—8–128 µg/mL | n.i. | ↓ cepA gene ↓ qacEΔ1 gene ↓ qacE gene | [86] |
| PHMG-Cl | P. aeruginosa ATCC 27853 | PHMG-Cl attaches high molecular weight DNA and plasmid DNA, resumes the process of inactivation of DNA from surfaces. | ↓ eDNA gene | [87] | |
| PAA SH CHDN | P. aeruginosa and P. fluorescens isolated from a cheese processing line at a dairy industry located in Sao Paulo, Brazil | MBCs: PAA—300 mg/L; SH—100 mg/L; CHDN—400 mg/L | PAA—bactericidal action linked to hypochlorous acid, crosses the cell membrane, oxidizes the sulfhydryl groups of certain enzymes; SH—strong oxidizing agent of the cytoplasmic membrane, deactivating physiological functions; CHDN—reacts with the negatively charged microbial cell surface, destroying the cell membrane, penetrates into the cell and causes leakage of intracellular components. | n.i. | [83] |
| LGPcide | P. fluorescens isolated from natural seawater from Praia dos Anjos Bay, Brazil | MBC reduction >99.5% after 24 h | High concentrations cause damage to cell membranes; disturbance of membrane structure and fluidity. | n.i. | [84] |
| GA GO | P. fluorescens | MBCs: GA—1000 µg/mL; GO—15000 µg/mL | GA—active membrane and oxidizing agent, reversible on the cell envelope if applied in low concentrations; GO—effects absent on bacterial surface, cell replication inhibitor (irreversible effects). | n.i. | [85] |
| ILs | P. aeruginosa PAO1 and PA14 | Permeabilization and disrupting the bacterial OM; interact with the lipid portion of the phospholipid, destabilizing the bilayer by breaking hydrophobic interaction between lipids responsible for the integrity of the membrane. | n.i. | [88] |
4.3. Klebsiella spp.
| Biocide | Strain | Concentration | Mechanism/Notes | Gene/Protein | Ref. |
|---|---|---|---|---|---|
| Chlorhexidine Octenidine | K. pneumoniae MGH 78578 | MICs: 8 mg/L and 1 mg/L; MBCs: 128 mg/L and 16 mg/L | n.i. | ↑ smvA ↑ smvR | [96] |
| Chlorhexidine Isopropanol | Klebsiella spp. from collections of the Universidad Cientifica de Sur in Lima, Peru. K. pneumoniae from HN2M, INMP, and HNGAI, Lima, Peru | MICs: 32 µg/mL (MIC50) and 64 µg/mL (MIC90) | n.i. | [93] | |
| Derdevice plus Y I&D Sept | K. pneumoniae CRKP isolates obtained from Gazi University, collection of the Microbiology Laboratory | MBC 1/300 dilution and 100% | n.i. | [97] | |
| Au, Cu, Pt, Pd, Ag | K. pneumoniae NCTC9633 | MICs: Au—5.85 mg/L; Cu—15.62 mg/L; Pt—3.90 mg/L; Pd—5.85 mg/L; Ag—11.71 mg/L; MBCs: Au—3.90 mg/L; Cu—15.62 mg/L; Pt—3.90 mg/L; Au—3.90 mg/L; Pd—3.90 mg/L | Toxic effects of metals can cause DNA damage, antioxidant depletion, disturbance of membrane function, structural changes in the cell wall, increased cell permeability, lysis of the cell. | n.i. | [94] |
| Cellulose foam paper with GA and BTCA | K. pneumoniae NCDC 138 | MIC 1–2 g/L | BTCA—crosslinking agent, reduces pH of the medium and inhibits bacterial growth; GA—cross-binding of the molecule with OM of bacterial cells interferes in the enzymatic activities and transport of the bacterial cell. | n.i. | [13] |
| Filter paper with Ca(OCl)2/AgNPs | K. pneumoniae isolated from different water supplies | MBC 2.0 mg/L (70%) 1.0 mg/L (50%) | Ag+ ions can bind and penetrate the cell membrane, increasing permeability. | n.i. | [95] |
4.4. Staphylococcus spp.
| Biocide | Strain | Concentration | Mechanism/Notes | Gene/Protein | Ref. |
|---|---|---|---|---|---|
| Benzalkonium chloride, H2O2, Biocide 1, Biocide 2, Biocide 3, mitomycin C | S. aureus 8325-4 and phages 8325-4Φ13 (CG1); RN420 8325-4Φ13-kana; MW2-ΦSa3mw; MW2c; 61599; 93616; DC10B; 8325-4Φ13attBmut; RN4220Φ13attBmut | MICs: BC—2.67 µg/mL; H2O2—0.03% w/w; Biocide 1—0.02%; Biocide 2—0.02%; Biocide 3—5.00 %; mitomycin C—0.12 mg/L | Known as hlb-converting phages which integrate in the hlb gene at the attB attachment site. | n.i. | [98] |
| Chlorhexidine | S. aureus isolates ST36 (qacA+ and qacA−); ST22 (qacA+ and qacA−); ST239-TW (qacA+) | ST22 clones—decrease 45% (1.47 mg/L); ST36 clones—1.75 mg/L | Increased binding to fibrinogen and fibronectin, increased adhesion and internalization into monolayers of keratinocytes, and confirms phenomenon of survival in vivo after chlorhexidine exposure. | n.i. | [105] |
| Hypochlorite and Phenol | MRSA and MSSA S. aureus clinical isolates, Egypt | Association between the presence of antiseptic resistance genes and the MICs | n.i. | ↑qacA/B, qacC genes | [100] |
| Chlorhexidine | 201 MRSA isolates from Portuguese hospitals, strain collection at ITQB-NOVA in Oeiras, Portugal | MICs 0.125–4 mg/L, one of them presented MICs 0.5–1 mg/L MBCs 0.125–8 mg/L | n.i. | sepA and mepA (100%); lmrS (87.1%); qacAB (22.4%); smr (1.0%) | [106] |
| F10SC Hexacon | S. pseudintermedius (MRSP, MSSP) and S. aureus (MRSA) clinical isolates, Sydney, Australia | MBCs: F10SC—1.05–16.87 mg/L; Hexacon—7.81–31.25 mg/L | n.i. | qacA/B genes—were only in MRSA isolates. qacJ (54%), qacG (29%), smr (7%) MRSP isolates. | [107] |
| Cu/SHfNP | S. aureus ATCC 6538 | Reduced cell growth at the highest concentration (1 mM) by 86% | Damage cell membrane, increase membrane’s permeability, disrupt cell membrane at high dose (2 mM). | n.i. | [79] |
| Dex-MA | S. aureus (MRSA) (ATCC 33591) | MICs: 1–2 µg/mL; MBCs: Dex-5—1.0 mg, Dex-10—1.9 mg, Dex-20—4.1 mg | n.i. | n.i. | [108] |
| PAA Chlorhexidine digluconate | S. aureus BZ012 and Sa30 from Brazilian dairies | MICs: PAA—0.075% (4.6 log) and 0.015% (1.1 log) mixed cultures; chlorhexidine dicluconate—0.0001953125 and 0.025% mixed cultures. | n.i. | n.i. | [109] |
| T-C@AgNPs | S. aureus (Bap-MRSA) | n.i. | Damage and destabilization of membrane of bacterial cell. Bioelectrical changes caused by biocide (intramembrane space to outside the cell) create pores—dose-dependent. | ↓Coa, ↓Eap, ↓SpA, ↓Bap genes | [102] |
| GTAgNPs | S. aureus 090 (MRSA090) | MIC 20 mg/mL | Presents antioxidant dose-dependent activity. Neutralizing ROS. | ↓Coa, ↓Eap, ↓SpA genes | [103] |
| PVDF/GN and PVDF/SP | S. aureus ATCC 25923 | Caused a 6-log reduction | Bactericidal action confirmed with the presence of intracellular ROS. | n.i. | [12] |
| MBQ (magnetic biochar/quaternary phosphonium salt) | S. aureus ATCC 6538 | MBC: 2 mg/L (50%), 50 mg/L (90%), dose-dependent effects | Loss of cell integrity, the appearance of vacuolization, rupture of cell surface and leakage of intracellular substances, Induced oxidative stress, penetrates through the lipid bilayers, and increases membrane structural destabilization. | n.i. | [73] |
| AuNSps AuNSts AuNCs | S. aureus from CICC, Beijing China | MICs 0.04 µg | Surface damage, disturbance, and cell loss by disruption of membrane-bound components, loss of flagella, cell integrity, leakage of cell contents into environment. | n.i. | [76] |
| CTAB | S. aureus (MRSA) | Cubic and tetrapod nanoparticles—14.19 log10 CFU/plate eradication; spheric nanoparticles—7.3 ± 0.2 log10 decrease | n.i. | n.i. | [104] |
| KFU-127 | S. aureus ATCC 29213, S. epidermidis clinical isolate from Kazan Institute, Russia | MICs: 4 µg/mL; MBCs: 8 µg/mL | Membrane potential changes and cell membrane damage; inhibitory property on pyroxidal-dependent enzymes. | n.i. | [11] |
4.5. Listeria spp.
4.6. Campylobacter spp.
4.7. Salmonella spp.
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Schirone, M.; Visciano, P.; Tofalo, R.; Suzzi, G. Editorial: Foodborne Pathogens: Hygiene and Safety. Front. Microbiol. 2019, 10, 1974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzales-Barron, U.; Thébault, A.; Kooh, P.; Watier, L.; Sanaa, M.; Cadavez, V. Strategy for systematic review of observational studies and meta-analysis modelling of risk factors for sporadic foodborne diseases. Microb. Risk Anal. 2021, 17, 100082. [Google Scholar] [CrossRef] [Green Version]
- Poole, K. Mechanisms of bacterial biocide and antibiotic resistance. J. Appl. Microbiol. 2002, 92, 55S–64S. [Google Scholar] [CrossRef] [PubMed]
- Santajit, S.; Indrawattana, N. Mechanisms of Antimicrobial Resistance in ESKAPE Pathogens. BioMed Res. Int. 2016, 2016, 2475067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rice, L.B. Progress and Challenges in Implementing the Research on ESKAPE Pathogens. Infect. Control Hosp. Epidemiol. 2010, 31, S7–S10. [Google Scholar] [CrossRef]
- Meade, E.; Slattery, M.; Garvey, M. Biocidal Resistance in Clinically Relevant Microbial Species: A Major Public Health Risk. Pathogens 2021, 10, 598. [Google Scholar] [CrossRef]
- Fernando, S.; Gunasekara, T.; Holton, J. Antimicrobial Nanoparticles: Applications and mechanisms of action. Sri Lankan J. Infect. Dis. 2018, 8, 2. [Google Scholar] [CrossRef]
- Fink, J.K. Hydraulic Fracturing Chemicals and Fluids Technology; Gulf Professional Publishing: Oxford, UK, 2013; pp. 1–235. [Google Scholar]
- McDonnell, G.; Russell, A.D. Antiseptics and Disinfectants: Activity, Action, and Resistance. Clin. Microbiol. Rev. 1999, 12, 147–179. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, P.; McBain, A.J.; Bloomfield, S.F. Biocide abuse and antimicrobial resistance: Being clear about the issues. J. Antimicrob. Chemother. 2002, 50, 137–139. [Google Scholar] [CrossRef]
- Garipov, M.R.; Sabirova, A.E.; Pavelyev, R.S.; Shtyrlin, N.V.; Lisovskaya, S.A.; Bondar, O.V.; Laikov, A.V.; Romanova, J.G.; Bogachev, M.I.; Kayumov, A.R.; et al. Targeting pathogenic fungi, bacteria and fungal-bacterial biofilms by newly synthesized quaternary ammonium derivative of pyridoxine and terbinafine with dual action profile. Bioorg. Chem. 2020, 104, 104306. [Google Scholar] [CrossRef]
- Samantaray, P.K.; Kumar, S.; Bose, S. ‘Polycation’ modified PVDF based antibacterial and antifouling membranes and ‘point-of-use supports’ for sustainable and effective water decontamination. J. Water Process. Eng. 2020, 38, 101536. [Google Scholar] [CrossRef]
- Heydarifard, S.; Taneja, K.; Bhanjana, G.; Dilbaghi, N.; Nazhad, M.M.; Kim, K.-H.; Kumar, S. Modification of cellulose foam paper for use as a high-quality biocide disinfectant filter for drinking water. Carbohydr. Polym. 2018, 181, 1086–1092. [Google Scholar] [CrossRef] [PubMed]
- Hughes, L.; Roberts, W.; Johnson, D. The impact of DNA adenine methyltransferase knockout on the development of triclosan resistance and antibiotic cross-resistance in Escherichia coli. Access Microbiol. 2021, 3, 000178. [Google Scholar] [CrossRef]
- Mann, R.; Holmes, A.; McNeilly, O.; Cavaliere, R.; Sotiriou, G.A.; Rice, S.A.; Gunawan, C. Evolution of biofilm-forming pathogenic bacteria in the presence of nanoparticles and antibiotic: Adaptation phenomena and cross-resistance. J. Nanobiotechnol. 2021, 19, 291. [Google Scholar] [CrossRef] [PubMed]
- Puangseree, J.; Jeamsripong, S.; Prathan, R.; Pungpian, C.; Chuanchuen, R. Resistance to widely-used disinfectants and heavy metals and cross resistance to antibiotics in Escherichia coli isolated from pigs, pork and pig carcass. Food Control 2021, 124, 107892. [Google Scholar] [CrossRef]
- Amsalu, A.; Sapula, S.A.; Lopes, M.D.B.; Hart, B.J.; Nguyen, A.H.; Drigo, B.; Turnidge, J.; Leong, L.E.; Venter, H. Efflux Pump-Driven Antibiotic and Biocide Cross-Resistance in Pseudomonas aeruginosa Isolated from Different Ecological Niches: A Case Study in the Development of Multidrug Resistance in Environmental Hotspots. Microorganisms 2020, 8, 1647. [Google Scholar] [CrossRef]
- Kahrilas, G.A.; Blotevogel, J.; Stewart, P.S.; Borch, T. Biocides in Hydraulic Fracturing Fluids: A Critical Review of Their Usage, Mobility, Degradation, and Toxicity. Environ. Sci. Technol. 2014, 49, 16–32. [Google Scholar] [CrossRef]
- Morente, E.O.; Fernández-Fuentes, M.A.; Burgos, M.J.G.; Abriouel, H.; Perez-Pulido, R.; Galvez, A. Biocide tolerance in bacteria. Int. J. Food Microbiol. 2013, 162, 13–25. [Google Scholar] [CrossRef]
- Balta, I.; Linton, M.; Pinkerton, L.; Kelly, C.; Ward, P.; Stef, L.; Pet, I.; Horablaga, A.; Gundogdu, O.; Corcionivoschi, N. The effect of natural antimicrobials on the Campylobacter coli T6SS+/− during in vitro infection assays and on their ability to adhere to chicken skin and carcasses. Int. J. Food Microbiol. 2021, 338, 108998. [Google Scholar] [CrossRef]
- Tabak, M.; Scher, K.; Hartog, E.; Romling, U.; Matthews, K.R.; Chikindas, M.L.; Yaron, S. Effect of triclosan on Salmonella typhimurium at different growth stages and in biofilms. FEMS Microbiol. Lett. 2007, 267, 200–206. [Google Scholar] [CrossRef] [Green Version]
- Balta, I.; Linton, M.; Pinkerton, L.; Kelly, C.; Stef, L.; Pet, I.; Stef, D.; Criste, A.; Gundogdu, O.; Corcionivoschi, N. The effect of natural antimicrobials against Campylobacter spp. and its similarities to Salmonella spp., Listeria spp., Escherichia coli, Vibrio spp., Clostridium spp. and Staphylococcus spp. Food Control 2020, 121, 107745. [Google Scholar] [CrossRef]
- Turner, R.J. Metal-based antimicrobial strategies. Microb. Biotechnol. 2017, 10, 1062–1065. [Google Scholar] [CrossRef] [PubMed]
- Harrison, J.J.; Ceri, H.; Stremick, C.A.; Turner, R.J. Biofilm susceptibility to metal toxicity. Environ. Microbiol. 2004, 6, 1220–1227. [Google Scholar] [CrossRef] [PubMed]
- Pal, C.; Bengtsson-Palme, J.; Kristiansson, E.; Larsson, D.G.J. Co-occurrence of resistance genes to antibiotics, biocides and metals reveals novel insights into their co-selection potential. BMC Genom. 2015, 16, 964. [Google Scholar] [CrossRef] [Green Version]
- Piddock, L.J.V. The crisis of no new antibiotics—What is the way forward? Lancet Infect. Dis. 2012, 12, 249–253. [Google Scholar] [CrossRef]
- Kayumov, A.R.; Nureeva, A.A.; Trizna, E.Y.; Gazizova, G.R.; Bogachev, M.; Shtyrlin, N.; Pugachev, M.; Sapozhnikov, S.V.; Shtyrlin, Y. New Derivatives of Pyridoxine Exhibit High Antibacterial Activity against Biofilm-Embedded Staphylococcus Cells. BioMed Res. Int. 2015, 2015, 890968. [Google Scholar] [CrossRef]
- Cheng, L.; Zhang, K.; Melo, M.; Weir, M.; Zhou, X.; Xu, H. Anti-biofilm Dentin Primer with Quaternary Ammonium and Silver Nanoparticles. J. Dent. Res. 2012, 91, 598–604. [Google Scholar] [CrossRef] [Green Version]
- Tischer, M.; Pradel, G.; Ohlsen, K.; Holzgrabe, U. Quaternary ammonium salts and their antimicrobial potential: Targets or nonspecific interactions? ChemMedChem 2012, 7, 22–31. [Google Scholar] [CrossRef]
- Shtyrlin, N.V.; Pugachev, M.V.; Sapozhnikov, S.V.; Garipov, M.R.; Vafina, R.M.; Grishaev, D.Y.; Pavelyev, R.S.; Kazakova, R.R.; Agafonova, M.N.; Iksanova, A.G.; et al. Novel Bis-Ammonium Salts of Pyridoxine: Synthesis and Antimicrobial Properties. Molecules 2020, 25, 4341. [Google Scholar] [CrossRef]
- Evans, A.; Kavanagh, K.A. Evaluation of metal-based antimicrobial compounds for the treatment of bacterial pathogens. J. Med. Microbiol. 2021, 70, 001363. [Google Scholar] [CrossRef]
- Juszkiewicz, M.; Walczak, M.; Woźniakowski, G. Characteristics of selected active substances used in disinfectants and their virucidal activity against ASFV. J. Veter. Res. 2019, 63, 17–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giuliano, C.A.; Rybak, M.J. Efficacy of Triclosan as an Antimicrobial Hand Soap and Its Potential Impact on Antimicrobial Resistance: A Focused Review. Pharmacother. J. Hum. Pharmacol. Drug Ther. 2015, 35, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Macri, D. Worldwide Use of Triclosan: Can Dentistry Do Without this Antimicrobial? Contemp. Clin. Dent. 2017, 8, 7–8. [Google Scholar] [CrossRef] [PubMed]
- Saleh, S.; Haddadin, R.; Baillie, S.; Collier, P. Triclosan—An update. Lett. Appl. Microbiol. 2010, 52, 87–95. [Google Scholar] [CrossRef]
- Köhler, A.T.; Rodloff, A.C.; Labahn, M.; Reinhardt, M.; Truyen, U.; Speck, S. Efficacy of sodium hypochlorite against multidrug-resistant Gram-negative bacteria. J. Hosp. Infect. 2018, 100, e40–e46. [Google Scholar] [CrossRef]
- Kampf, G.; Todt, D.; Pfaender, S.; Steinmann, E. Persistence of coronaviruses on inanimate surfaces and their inactivation with biocidal agents. J. Hosp. Infect. 2020, 104, 246–251, Corrigendum in J. Hosp. Infect. 2020, 105, 587. [Google Scholar] [CrossRef] [Green Version]
- Vereshchagin, A.; Frolov, N.; Egorova, K.; Seitkalieva, M.; Ananikov, V. Quaternary Ammonium Compounds (QACs) and Ionic Liquids (ILs) as Biocides: From Simple Antiseptics to Tunable Antimicrobials. Int. J. Mol. Sci. 2021, 22, 6793. [Google Scholar] [CrossRef]
- Shaban, S.M.; Saied, A.; Tawfik, S.M.; Abd-Elaal, A.; Aiad, I. Corrosion inhibition and Biocidal effect of some cationic surfactants based on Schiff base. J. Ind. Eng. Chem. 2013, 19, 2004–2009. [Google Scholar] [CrossRef]
- Borges, A.; Abreu, A.C.; Dias, C.; Saavedra, M.J.; Borges, F.; Simões, M. New Perspectives on the Use of Phytochemicals as an Emergent Strategy to Control Bacterial Infections Including Biofilms. Molecules 2016, 21, 877. [Google Scholar] [CrossRef]
- Shin, S.-A.; Joo, B.J.; Lee, J.; Ryu, G.; Han, M.; Kim, W.-Y.; Park, H.H.; Lee, J.; Lee, C.S. Phytochemicals as Anti-Inflammatory Agents in Animal Models of Prevalent Inflammatory Diseases. Molecules 2020, 25, 5932. [Google Scholar] [CrossRef]
- Paulus, W. Directory of Microbicides for the Protection of Materials: A Handbook; Springer Science & Business Media: Berlin, Germany, 2005. [Google Scholar]
- Denyer, S.P.; Stewart, G.S.A.B. Mechanisms of action of disinfectants. Int. Biodeterior. Biodegrad. 1998, 41, 261–268. [Google Scholar] [CrossRef]
- Araújo, P.; Lemos, M.; Mergulhão, F.; Melo, L.; Simões, M. Antimicrobial resistance to disinfectants in biofilms. Sci. Against Microb. Pathog. Commun. Curr. Res. Technol. Adv. 2011, 3, 826–834. [Google Scholar]
- Mitra, D.; Kang, E.-T.; Neoh, K.G. Antimicrobial Copper-Based Materials and Coatings: Potential Multifaceted Biomedical Applications. ACS Appl. Mater. Interfaces 2019, 12, 21159–21182. [Google Scholar] [CrossRef]
- Jamaati, H.; Mortaz, E.; Pajouhi, Z.; Folkerts, G.; Movassaghi, M.; Moloudizargari, M.; Adcock, I.M.; Garssen, J. Nitric Oxide in the Pathogenesis and Treatment of Tuberculosis. Front. Microbiol. 2017, 8, 2008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalantari, K.; Afifi, A.M.; Jahangirian, H.; Webster, T.J. Biomedical applications of chitosan electrospun nanofibers as a green polymer—Review. Carbohydr. Polym. 2018, 207, 588–600. [Google Scholar] [CrossRef]
- Jiao, Y.; Niu, L.-N.; Ma, S.; Li, J.; Tay, F.R.; Chen, J.-H. Quaternary ammonium-based biomedical materials: State-of-the-art, toxicological aspects and antimicrobial resistance. Prog. Polym. Sci. 2017, 71, 53–90. [Google Scholar] [CrossRef]
- Chai, D.; Liu, W.; Hao, X.; Wang, H.; Wang, H.; Hao, Y.; Gao, Y.; Qu, H.; Wang, L.; Dong, A.; et al. Mussel-inspired synthesis of magnetic N-Halamine nanoparticles for antibacterial recycling. Colloids Interface Sci. Commun. 2020, 39, 100320. [Google Scholar] [CrossRef]
- Qian, Y.X.; Zhang, D.F.; Wu, Y.M.; Chen, Q.; Liu, R.H. The design, synthesis and biological activity study of nylon-3 polymers as mimics of host defense peptides. Acta Polym. Sin. 2016, 10, 1300–1311. [Google Scholar]
- Ding, X.; Duan, S.; Ding, X.; Liu, R.; Xu, F.J. Versatile Antibacterial Materials: An Emerging Arsenal for Combatting Bacterial Pathogens. Adv. Funct. Mater. 2018, 28, 1802140. [Google Scholar] [CrossRef]
- Thallinger, B.; Prasetyo, E.N.; Nyanhongo, G.S.; Guebitz, G.M. Antimicrobial enzymes: An emerging strategy to fight microbes and microbial biofilms. Biotechnol. J. 2013, 8, 97–109. [Google Scholar] [CrossRef]
- Dong, A.; Wang, Y.-J.; Gao, Y.; Gao, T.; Gao, G. Chemical Insights into Antibacterial N-Halamines. Chem. Rev. 2017, 117, 4806–4862. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.J.; Muszanska, A.; Xiang, F.; Falkenberg, R.; Van De Belt-Gritter, B.; Loontjens, T.J.A. Contact Killing of Gram-Positive and Gram-Negative Bacteria on PDMS Provided with Immobilized Hyperbranched Antibacterial Coatings. Langmuir 2019, 35, 14108–14116. [Google Scholar] [CrossRef] [PubMed]
- Russell, A. Biocide use and antibiotic resistance: The relevance of laboratory findings to clinical and environmental situations. Lancet Infect. Dis. 2003, 3, 794–803. [Google Scholar] [CrossRef]
- Gilbert, P.; McBain, A.J. Potential Impact of Increased Use of Biocides in Consumer Products on Prevalence of Antibiotic Resistance. Clin. Microbiol. Rev. 2003, 16, 189–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cloete, T. Resistance mechanisms of bacteria to antimicrobial compounds. Int. Biodeterior. Biodegrad. 2003, 51, 277–282. [Google Scholar] [CrossRef] [Green Version]
- Forbes, S.; Cowley, N.; Humphreys, G.; Mistry, H.; Amézquita, A.; McBain, A.J. Formulation of Biocides Increases Antimicrobial Potency and Mitigates the Enrichment of Nonsusceptible Bacteria in Multispecies Biofilms. Appl. Environ. Microbiol. 2017, 83, e03054-16. [Google Scholar] [CrossRef] [Green Version]
- White, D.G.; McDermott, P.F. Biocides, drug resistance and microbial evolution. Curr. Opin. Microbiol. 2001, 4, 313–317. [Google Scholar] [CrossRef]
- Fleitas, O.; Franco, O.L. Induced Bacterial Cross-Resistance toward Host Antimicrobial Peptides: A Worrying Phenomenon. Front. Microbiol. 2016, 7, 381. [Google Scholar] [CrossRef] [Green Version]
- Calvert, M.B.; Jumde, V.R.; Titz, A. Pathoblockers or antivirulence drugs as a new option for the treatment of bacterial infections. Beilstein J. Org. Chem. 2018, 14, 2607–2617. [Google Scholar] [CrossRef]
- Davin-Regli, A.; Pagès, J.M. Cross-resistance between biocides and antimicrobials: An emerging question. Rev. Sci. Tech. 2012, 31, 89–104. [Google Scholar] [CrossRef]
- Sonbol, F.I.; El-Banna, T.E.; El-Aziz, A.A.; El-Ekhnawy, E. Impact of triclosan adaptation on membrane properties, efflux and antimicrobial resistance of Escherichia coli clinical isolates. J. Appl. Microbiol. 2018, 126, 730–739. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Miller, W.; Arias, C.A. Mechanisms of antimicrobial resistance among hospital-associated pathogens. Expert Rev. Anti-Infect. Ther. 2018, 16, 269–287. [Google Scholar] [CrossRef] [PubMed]
- Baker-Austin, C.; Wright, M.S.; Stepanauskas, R.; McArthur, J. Co-selection of antibiotic and metal resistance. Trends Microbiol. 2006, 14, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Guentzel, M.N. Escherichia, Klebsiella, Enterobacter, Serratia, Citrobacterv, and Proteus. In Medical Microbiology; Baron, S., Ed.; University of Texas Medical Branch at Galveston: Galveston, TX, USA, 1996. [Google Scholar]
- Stratakos, A.C.; Sima, F.; Ward, P.; Linton, M.; Kelly, C.; Pinkerton, L.; Stef, L.; Pet, I.; Corcionivoschi, N. The in vitro effect of carvacrol, a food additive, on the pathogenicity of O157 and non-O157 Shiga-toxin producing Escherichia coli. Food Control 2017, 84, 290–296. [Google Scholar] [CrossRef]
- Foglarova, M.; Prokop, J.; Milichovsky, M. Oxidized cellulose: An application in the form of sorption filter materials. J. Appl. Polym. Sci. 2009, 112, 669–678. [Google Scholar] [CrossRef]
- Lee, A.; Elam, J.W.; Darling, S.B. Membrane materials for water purification: Design, development, and application. Environ. Sci. Water Res. Technol. 2016, 2, 17–42. [Google Scholar] [CrossRef]
- Meireles, A.; Ferreira, C.; Melo, L.; Simões, M. Comparative stability and efficacy of selected chlorine-based biocides against Escherichia coli in planktonic and biofilm states. Food Res. Int. 2017, 102, 511–518. [Google Scholar] [CrossRef]
- Smirnova, M.; Kolodkin, N.; Kolobov, A.; Afonin, V.; Afonina, I.; Stefanenko, L.; Shpen’, V.; Shamova, O. Indolicidin analogs with broad-spectrum antimicrobial activity and low hemolytic activity. Peptides 2020, 132, 170356. [Google Scholar] [CrossRef]
- Ali, S.; Khan, S.J.; Allan, R.; Hashmi, I. Biocidal potential of electrochemically activated solutions (ECAS) against Aeromonas sp. Enterobacter sp. and Escherichia coli in tap water. J. Water Process Eng. 2020, 36, 101328. [Google Scholar] [CrossRef]
- Fu, Y.; Wang, F.; Sheng, H.; Xu, M.; Liang, Y.; Bian, Y.; Hashsham, S.A.; Jiang, X.; Tiedje, J.M. Enhanced antibacterial activity of magnetic biochar conjugated quaternary phosphonium salt. Carbon 2020, 163, 360–369. [Google Scholar] [CrossRef]
- Campo, N.; De Flora, C.; Maffettone, R.; Manoli, K.; Sarathy, S.; Santoro, D.; Gonzalez-Olmos, R.; Auset, M. Inactivation kinetics of antibiotic resistant Escherichia coli in secondary wastewater effluents by peracetic and performic acids. Water Res. 2019, 169, 115227. [Google Scholar] [CrossRef] [PubMed]
- Zarrella, I.; Falivene, L.; Galiakberov, V.; Fiorentino, A.; Cucciniello, R.; Motta, O.; Rizzo, L.; Krasnogorskaya, N.; Proto, A. Effect of the aqueous matrix on the inactivation of E. coli by permaleic acid. Sci. Total Environ. 2020, 767, 144395. [Google Scholar] [CrossRef] [PubMed]
- Hameed, S.; Wang, Y.; Zhao, L.; Xie, L.; Ying, Y. Shape-dependent significant physical mutilation and antibacterial mechanisms of gold nanoparticles against foodborne bacterial pathogens (Escherichia coli, Pseudomonas aeruginosa and Staphylococcus aureus) at lower concentrations. Mater. Sci. Eng. C 2019, 108, 110338. [Google Scholar] [CrossRef] [PubMed]
- Theofilou, S.; Antoniou, C.; Potamiti, L.; Hadjisavvas, A.; Panayiotidis, M.; Savva, P.; Costa, C.; Fotopoulos, V. Immobilized Ag-nanoparticles (iNPs) for environmental applications: Elucidation of immobilized silver-induced inhibition mechanism of Escherichia coli. J. Environ. Chem. Eng. 2021, 9, 106001. [Google Scholar] [CrossRef]
- Santo, C.E.; Taudte, N.; Nies, D.H.; Grass, G. Contribution of Copper Ion Resistance to Survival of Escherichia coli on Metallic Copper Surfaces. Appl. Environ. Microbiol. 2008, 74, 977–986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manoharan, R.K.; Gangadaran, P.; Ayyaru, S.; Ahn, B.-C.; Ahn, Y.-H. Self-healing functionalization of sulfonated hafnium oxide and copper oxide nanocomposite for effective biocidal control of multidrug-resistant bacteria. New J. Chem. 2021, 45, 9506–9517. [Google Scholar] [CrossRef]
- Si, Y.; Zhang, Z.; Wu, W.; Fu, Q.; Huang, K.; Nitin, N.; Ding, B.; Sun, G. Daylight-driven rechargeable antibacterial and antiviral nanofibrous membranes for bioprotective applications. Sci. Adv. 2018, 4, eaar5931. [Google Scholar] [CrossRef] [Green Version]
- Ciofu, O.; Tolker-Nielsen, T. Tolerance and Resistance of Pseudomonas aeruginosa Biofilms to Antimicrobial Agents-How P. aeruginosa Can Escape Antibiotics. Front. Microbiol. 2019, 10, 913. [Google Scholar] [CrossRef] [Green Version]
- Høiby, N.; Ciofu, O.; Bjarnsholt, T. Pseudomonas aeruginosa biofilms in cystic fibrosis. Futur. Microbiol. 2010, 5, 1663–1674. [Google Scholar] [CrossRef]
- Castro, M.S.R.; Fernandes, M.D.S.; Kabuki, D.Y.; Kuaye, A.Y. Modelling Pseudomonas fluorescens and Pseudomonas aeruginosa biofilm formation on stainless steel surfaces and controlling through sanitisers. Int. Dairy J. 2020, 114, 104945. [Google Scholar] [CrossRef]
- Batista, W.R.; Fernandes, F.C.; Neves, M.H.; Nascimento, T.; Lopes, R.; Lopes, C.; Ziegler, G.P.; Figueroa, B.M.S.; Sparks, D.; Fontaine, D.; et al. Synthetic lipids as a biocide candidate for disinfection of ballast water. Mar. Pollut. Bull. 2018, 137, 702–710. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, S.; Gomes, I.B.; Simões, M. Antimicrobial activity of glycolic acid and glyoxal against Bacillus cereus and Pseudomonas fluorescens. Food Res. Int. 2020, 136, 109346. [Google Scholar] [CrossRef]
- Goodarzi, R.; Yousefimashouf, R.; Taheri, M.; Nouri, F.; Asghari, B. Susceptibility to biocides and the prevalence of biocides resistance genes in clinical multidrug-resistant Pseudomonas aeruginosa isolates from Hamadan, Iran. Mol. Biol. Rep. 2021, 48, 5275–5281. [Google Scholar] [CrossRef]
- Moshynets, O.V.; Baranovskyi, T.P.; Iungin, O.S.; Kysil, N.P.; Metelytsia, L.O.; Pokholenko, I.; Potochilova, V.V.; Potters, G.; Rudnieva, K.L.; Rymar, S.Y.; et al. eDNA Inactivation and Biofilm Inhibition by the PolymericBiocide Polyhexamethylene Guanidine Hydrochloride (PHMG-Cl). Int. J. Mol. Sci. 2022, 23, 731. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.D.T.; Voskian, S.; Brown, P.; Liu, A.; Lu, T.K.; Hatton, T.A.; De La Fuente-Nunez, C. Coatable and Resistance-Proof Ionic Liquid for Pathogen Eradication. ACS Nano 2021, 15, 966–978. [Google Scholar] [CrossRef] [PubMed]
- Rendueles, O. Deciphering the role of the capsule of Klebsiella pneumoniae during pathogenesis: A cautionary tale. Mol. Microbiol. 2020, 113, 883–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, K.J.; Ray, C.G. Sherris Medical Microbiology; Mcgraw-Hill: New York, NY, USA, 2003. [Google Scholar]
- Bagley, S.T. Habitat Association of Klebsiella Species. Infect. Control 1985, 6, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Perez-Palacios, P.; Gual-De-Torrella, A.; Delgado-Valverde, M.; Oteo-Iglesias, J.; Hidalgo-Díaz, C.; Pascual, A.; Fernández-Cuenca, F. Transfer of plasmids harbouring blaOXA-48-like carbapenemase genes in biofilm-growing Klebsiella pneumoniae: Effect of biocide exposure. Microbiol. Res. 2021, 254, 126894. [Google Scholar] [CrossRef] [PubMed]
- Morante, J.; Quispe, A.M.; Ymaña, B.; Moya-Salazar, J.; Luque, N.; Soza, G.; Chirinos, M.R.; Pons, M.J. Tolerance to disinfectants (chlorhexidine and isopropanol) and its association with antibiotic resistance in clinically-related Klebsiella pneumoniae isolates. Pathog. Glob. Health 2021, 115, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Vaidya, M.Y.; McBain, A.J.; Butler, J.; Banks, C.E.; Whitehead, K.A. Antimicrobial Efficacy and Synergy of Metal Ions against Enterococcus faecium, Klebsiella pneumoniae and Acinetobacter baumannii in Planktonic and Biofilm Phenotypes. Sci. Rep. 2017, 7, 5911. [Google Scholar] [CrossRef]
- Mohammed, A.N. Resistance of bacterial pathogens to calcium hypochlorite disinfectant and evaluation of the usability of treated filter paper impregnated with nanosilver composite for drinking water purification. J. Glob. Antimicrob. Resist. 2018, 16, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Wand, M.E.; Jamshidi, S.; Bock, L.; Rahman, K.M.; Sutton, J.M. SmvA is an important efflux pump for cationic biocides in Klebsiella pneumoniae and other Enterobacteriaceae. Sci. Rep. 2019, 9, 1344. [Google Scholar] [CrossRef] [PubMed]
- Ozturk, A.; Güzel, Ö.T.; Abdulmajed, O.; Erdoğan, M.; Kozan, R.; Çağlar, K.; Kalkancı, A. Evaluation of the bactericidal activity of some disinfectant agents against carbapenem-resistant Klebsiella pneumoniae isolates. Int. J. Environ. Health Eng. 2020, 9, 17. [Google Scholar]
- Tang, Y.; Nielsen, L.N.; Hvitved, A.; Haaber, J.K.; Wirtz, C.; Andersen, P.S.; Larsen, J.; Wolz, C.; Ingmer, H. Commercial Biocides Induce Transfer of Prophage Φ13 from Human Strains of Staphylococcus aureus to Livestock CC398. Front. Microbiol. 2017, 8, 2418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, L.B.; Stegger, M.; Hasman, H.; Aziz, M.; Larsen, J.; Andersen, P.S.; Pearson, T.; Waters, A.E.; Foster, J.T.; Schupp, J.; et al. Staphylococcus aureus CC398: Host Adaptation and Emergence of Methicillin Resistance in Livestock. mBio 2012, 3, e00305-11. [Google Scholar] [CrossRef] [Green Version]
- Sabry, W.; Al-Dakhakhni, M.Y.S.; Al-Hoseiny, I.M.; Tawab, R.A.R.A. Detection of Biocides Resistance Genes among Clinical Isolates of Both Methicillin-Resistant and Methicillin-Sensitive Strains of Staphylococcus aureus. Int. J. Med. Arts 2021, 3, 1602–1610. [Google Scholar] [CrossRef]
- Kernberger-Fischer, I.A.; Krischek, C.; Strommenger, B.; Fiegen, U.; Beyerbach, M.; Kreienbrock, L.; Klein, G.; Kehrenberg, C. Susceptibility of Methicillin-Resistant and -Susceptible Staphylococcus aureus Isolates of Various Clonal Lineages from Germany to Eight Biocides. Appl. Environ. Microbiol. 2018, 84, e00799-18. [Google Scholar] [CrossRef] [Green Version]
- Manukumar, H.M.; Chandrasekhar, B.; Rakesh, K.P.; Ananda, A.P.; Nandhini, M.; Lalitha, P.; Sumathi, S.; Qinb, H.-L.; Umesha, S. Novel T-C@AgNPs mediated biocidal mechanism against biofilm associated methicillin-resistant Staphylococcus aureus (Bap-MRSA) 090, cytotoxicity and its molecular docking studies. Medchemcomm 2017, 8, 2181–2194. [Google Scholar] [CrossRef]
- Manukumar, H.; Yashwanth, B.; Umesha, S.; Rao, J.V. Biocidal mechanism of green synthesized thyme loaded silver nanoparticles (GTAgNPs) against immune evading tricky methicillin-resistant Staphylococcus aureus 090 (MRSA090) at a homeostatic environment. Arab. J. Chem. 2020, 13, 1179–1197. [Google Scholar] [CrossRef]
- Nickel, R.; Kazemian, M.R.; Wroczynskyj, Y.; Liu, S.; van Lierop, J. Exploiting shape-selected iron oxide nanoparticles for the destruction of robust bacterial biofilms—Active transport of biocides via surface charge and magnetic field control. Nanoscale 2020, 12, 4328–4333. [Google Scholar] [CrossRef]
- Marbach, H.; Vizcay-Barrena, G.; Memarzadeh, K.; Otter, J.A.; Pathak, S.; Allaker, R.P.; Harvey, R.D.; Edgeworth, J.D. Tolerance of MRSA ST239-TW to chlorhexidine-based decolonization: Evidence for keratinocyte invasion as a mechanism of biocide evasion. J. Infect. 2018, 78, 119–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conceição, T.; de Lencastre, H.; Aires-De-Sousa, M. Prevalence of biocide resistance genes and chlorhexidine and mupirocin non-susceptibility in Portuguese hospitals during a 31-year period (1985–2016). J. Glob. Antimicrob. Resist. 2020, 24, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Worthing, K.; Marcus, A.; Abraham, S.; Trott, D.J.; Norris, J.M. Qac genes and biocide tolerance in clinical veterinary methicillin-resistant and methicillin-susceptible Staphylococcus aureus and Staphylococcus pseudintermedius. Veter.-Microbiol. 2018, 216, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Hoque, J.; Haldar, J. Direct Synthesis of Dextran-Based Antibacterial Hydrogels for Extended Release of Biocides and Eradication of Topical Biofilms. ACS Appl. Mater. Interfaces 2017, 9, 15975–15985. [Google Scholar] [CrossRef] [PubMed]
- Oxaran, V.; Dittmann, K.K.; Lee, S.H.I.; Chaul, L.T.; de Oliveira, C.A.F.; Corassin, C.H.; Alves, V.F.; De Martinis, E.C.P.; Gram, L. Behavior of Foodborne Pathogens Listeria monocytogenes and Staphylococcus aureus in Mixed-Species Biofilms Exposed to Biocides. Appl. Environ. Microbiol. 2018, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-López, P.; Rodríguez-Herrera, J.J.; Vázquez-Sánchez, D.; Cabo, M.L. Current Knowledge on Listeria monocytogenes Biofilms in Food-Related Environments: Incidence, Resistance to Biocides, Ecology and Biocontrol. Foods 2018, 7, 85. [Google Scholar] [CrossRef] [Green Version]
- Stratakos, A.C.; Ijaz, U.Z.; Ward, P.; Linton, M.; Kelly, C.; Pinkerton, L.; Scates, P.; McBride, J.; Pet, I.; Criste, A.; et al. In vitro and in vivo characterisation of Listeria monocytogenes outbreak isolates. Food Control 2020, 107. [Google Scholar] [CrossRef]
- Duze, S.T.; Marimani, M.; Patel, M. Tolerance of Listeria monocytogenes to biocides used in food processing environments. Food Microbiol. 2021, 97, 103758. [Google Scholar] [CrossRef]
- Roedel, A.; Dieckmann, R.; Brendebach, H.; Hammerl, J.A.; Kleta, S.; Noll, M.; Al Dahouk, S.; Vincze, S. Biocide-Tolerant Listeria monocytogenes Isolates from German Food Production Plants Do Not Show Cross-Resistance to Clinically Relevant Antibiotics. Appl. Environ. Microbiol. 2019, 85, e01253-19. [Google Scholar] [CrossRef] [Green Version]
- Guérin, A.; Bridier, A.; Le Grandois, P.; Sévellec, Y.; Palma, F.; Félix, B.; Roussel, S.; Soumet, C.; LISTADAPT Study Group. Exposure to Quaternary Ammonium Compounds Selects Resistance to Ciprofloxacin in Listeria monocytogenes. Pathogens 2021, 10, 220. [Google Scholar] [CrossRef]
- Noll, M.; Trunzer, K.; Vondran, A.; Vincze, S.; Dieckmann, R.; Al Dahouk, S.; Gold, C. Benzalkonium Chloride Induces a VBNC State in Listeria monocytogenes. Microorganisms 2020, 8, 184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kropac, A.; Eshwar, A.K.; Stephan, R.; Tasara, T. New Insights on the Role of the pLMST6 Plasmid in Listeria monocytogenes Biocide Tolerance and Virulence. Front. Microbiol. 2019, 10, 1538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mafuna, T.; Matle, I.; Magwedere, K.; Pierneef, R.E.; Reva, O.N. Whole Genome-Based Characterization of Listeria monocytogenes Isolates Recovered from the Food Chain in South Africa. Front. Microbiol. 2021, 12, 669287. [Google Scholar] [CrossRef] [PubMed]
- Dygico, L.K.; Gahan, C.G.; Grogan, H.; Burgess, C.M. Examining the efficacy of mushroom industry biocides on Listeria monocytogenes biofilm. J. Appl. Microbiol. 2020, 130, 1106–1116. [Google Scholar] [CrossRef]
- Luque-Sastre, L.; Fox, E.; Jordan, K.; Fanning, S. A Comparative Study of the Susceptibility of Listeria Species to Sanitizer Treatments When Grown under Planktonic and Biofilm Conditions. J. Food Prot. 2018, 81, 1481–1490. [Google Scholar] [CrossRef]
- Haubert, L.; Zehetmeyr, M.L.; Da Silva, W.P. Resistance to benzalkonium chloride and cadmium chloride in Listeria monocytogenes isolates from food and food-processing environments in southern Brazil. Can. J. Microbiol. 2019, 65, 429–435. [Google Scholar] [CrossRef]
- Rodríguez-Melcón, C.; Capita, R.; Rodríguez-Jerez, J.J.; Martínez-Suárez, J.V.; Alonso-Calleja, C. Effect of Low Doses of Disinfectants on the Biofilm-Forming Ability of Listeria monocytogenes. Foodborne Pathog. Dis. 2019, 16, 262–268. [Google Scholar] [CrossRef]
- Vázquez-Sánchez, D.; Galvão, J.A.; Oetterer, M. Contamination sources, serogroups, biofilm-forming ability and biocide resistance of Listeria monocytogenes persistent in tilapia-processing facilities. J. Food Sci. Technol. 2017, 54, 3867–3879. [Google Scholar] [CrossRef]
- Rodríguez-Melcón, C.; Riesco-Peláez, F.; Fernández, M.D.C.G.; Alonso-Calleja, C.; Capita, R. Susceptibility of Listeria monocytogenes planktonic cultures and biofilms to sodium hypochlorite and benzalkonium chloride. Food Microbiol. 2019, 82, 533–540. [Google Scholar] [CrossRef]
- Møretrø, T.; Schirmer, B.C.; Heir, E.; Fagerlund, A.; Hjemli, P.; Langsrud, S. Tolerance to quaternary ammonium compound disinfectants may enhance growth of Listeria monocytogenes in the food industry. Int. J. Food Microbiol. 2017, 241, 215–224. [Google Scholar] [CrossRef] [Green Version]
- López-Alonso, V.; Ortiz, S.; Corujo, A.; Martínez-Suárez, J.V. Analysis of Benzalkonium Chloride Resistance and Potential Virulence of Listeria monocytogenes Isolates Obtained from Different Stages of a Poultry Production Chain in Spain. J. Food Prot. 2020, 83, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Karki, A.B.; Wells, H.; Fakhr, M.K. Retail liver juices enhance the survivability of Campylobacter jejuni and Campylobacter coli at low temperatures. Sci. Rep. 2019, 9, 2733. [Google Scholar] [CrossRef] [PubMed]
- Beier, R.C.; Byrd, J.A.; Andrews, K.; Caldwell, D.; Crippen, T.L.; Anderson, R.C.; Nisbet, D.J. Disinfectant and antimicrobial susceptibility studies of the foodborne pathogen Campylobacter jejuni isolated from the litter of broiler chicken houses. Poult. Sci. 2020, 100, 1024–1033. [Google Scholar] [CrossRef] [PubMed]
- Beier, R.C.; Harvey, R.B.; Hernandez, C.A.; Andrews, K.; Droleskey, R.E.; Hume, M.E.; Davidson, M.K.; Bodeis-Jones, S.; Young, S.; Anderson, R.C.; et al. Disinfectant and Antimicrobial Susceptibility Profiles of Campylobacter coli Isolated in 1998 to 1999 and 2015 from Swine and Commercial Pork Chops. J. Food Sci. 2019, 84, 1501–1512. [Google Scholar] [CrossRef] [PubMed]
- Muhandiramlage, G.K.; McWhorter, A.R.; Chousalkar, K.K. Chlorine Induces Physiological and Morphological Changes on Chicken Meat Campylobacter Isolates. Front. Microbiol. 2020, 11, 503. [Google Scholar] [CrossRef]
- Talukdar, P.K.; Turner, K.L.; Crockett, T.M.; Lu, X.; Morris, C.F.; Konkel, M.E. Inhibitory Effect of Puroindoline Peptides on Campylobacter jejuni Growth and Biofilm Formation. Front. Microbiol. 2021, 12, 702762. [Google Scholar] [CrossRef]
- Paliy, A.B.T.; Stegniy, V.I.; Bolotin, K.A.; Rodionova, T.M.; Ihnatieva, O.M.; Bobrytska, V.Y.; Kushnir, O.V.; Matsenko, L.L.; Kushch, V.S.; Bulavina, O.V.; et al. Study of disinfectants bactericidal properties against Campylobacter spp. at sub-zero temperatures. Ukranina J. Eclogy 2021, 11, 391–398. [Google Scholar]
- Sibanda, N.; McKenna, A.; Richmond, A.; Ricke, S.C.; Callaway, T.; Stratakos, A.C.; Gundogdu, O.; Corcionivoschi, N. A Review of the Effect of Management Practices on Campylobacter Prevalence in Poultry Farms. Front. Microbiol. 2018, 9, 2002. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA). ECDC (European Food Safety Authority and European Centre for Disease Prevention and Control) The European Union One Health 2018 Zoonoses Report. EFSA J. 2021, 19, e06406. [Google Scholar]
- Lamas, A.; Miranda, J.; Regal, P.; Vazquez, B.; Franco, C.M.; Cepeda, A. A comprehensive review of non-enterica subspecies of Salmonella enterica. Microbiol. Res. 2018, 206, 60–73. [Google Scholar] [CrossRef]
- Obe, T.; Nannapaneni, R.; Schilling, W.; Zhang, L.; Kiess, A. Antimicrobial tolerance, biofilm formation, and molecular characterization of Salmonella isolates from poultry processing equipment. J. Appl. Poult. Res. 2021, 30, 100195. [Google Scholar] [CrossRef]
- Strantzali, D.; Kostoglou, D.; Perikleous, A.; Zestas, M.; Ornithopoulou, S.; Dubois-Brissonnet, F.; Giaouris, E. Comparative assessment of the disinfection effectiveness of thymol and benzalkonium chloride against adapted and non-adapted to thymol biofilm cells of a Salmonella typhimurium epidemic phage type DT193 strain. Food Control 2021, 129, 108239. [Google Scholar] [CrossRef]
- Nascente, E.D.P.; Rauecker, N.; Teles, A.V.; Divino, L.D.D.A.; Chagas, S.R.; Elias, L.M.; Matos, M.P.C.; Andrade, M.A.; Pascoal, L.M. Inactivation of Multidrug-Resistant Salmonella Heidelberg Isolated from Pigs by Ozonated Water under Different Conditions. Ozone: Sci. Eng. 2021, 1–9. [Google Scholar] [CrossRef]
- De Melo, R.T.; dos Reis Cardoso, T.; Peres PA, B.M.; Braz, R.F.; Monteiro, G.P.; Rossi, D.A. Salmonella enterica Serovar Minnesota Biofilms, Susceptibility to Biocides, and Molecular Characterization. Pathogens 2021, 10, 581. [Google Scholar] [CrossRef] [PubMed]
- Algburi, A.; Asghar, A.; Huang, Q.; Mustfa, W.; Javed, H.U.; Zehm, S.; Chikindas, M.L. Black cardamom essential oil prevents Escherichia coli O157:H7 and Salmonella typhimurium JSG 1748 biofilm formation through inhibition of quorum sensing. J. Food Sci. Technol. 2021, 58, 3183–3191. [Google Scholar]

| Biocide | Strain | Concentration | Mechanism/Notes | Gene/Protein | Ref. |
|---|---|---|---|---|---|
| Benzalkonium chloride | L. monocytogenes SLCC2540 | MIC 4 µg/mL; MBC 11 µg/mL | Increased abundance of viable but nonculturable cells. | May induce mutations in efflux pump systems, which are responsible for multidrug resistance | [115] |
| L. monocytogenes ST6 | Greater than 10 μg /mL | n.i. | BC tolerance due to the presence of pLMST6 plasmid (EmrC gene responsible for the multidrug efflux pump protein) | [116] | |
| L. monocytogenes isolated from South Africa food chain (e.g., ST1, ST2, ST121, ST204, and ST321) | n.i. | n.i. | ↑bcrABC cassette ↑ermC ↑mdrL ↑Ide ↑SSI-1 ↑ SSI-2 | [117] | |
| BP1-BP5BP6-BP11 | L. monocytogenes isolated strains 2075, 2081, and 2355 from the mushroom production environment. Strains 3050, 3051, 3101, 3102, and 3104 isolated from the production environment after steam cookout | 1.7-log and 6-log reductions; the highest reduction, 6-log—BP6 and BP11 MICs ranged between 1 and 50% | n.i. | n.i. | [118] |
| Triclosan | L. monocytogenesL. welshimeri | MIC from 0.0015 to 0.006% | n.i. | n.i. | [119] |
| Cadmium chloride | L. monocytogenes isolates from food-processing environments, Brazil | Ranged from 20 to 200 µg.mL−1 | n.i. | Presence of the efflux pumps MdrL and Lde, cadmium chloride resistance | [120] |
| PAA Chlorhexidine | Listeria monocytogenes BZ001 isolated from a single food-processing environment | MIC 0.075% for PAA and 0.000390625% for chlorhexidine | n.i. | n.i. | [109] |
| Biocide | Strain | Concentration | Mechanism/Notes | Gene/Protein | Ref. |
|---|---|---|---|---|---|
| Triclosan Chlorhexidine Benzalkonium chloride | 96 C. jejuni strains isolated from poultry industry | MIC50 = 32 and MIC90 = 32 (μg/mL); MIC50 = 0.5 and MIC90 = 1; MIC50 = 1 and MIC90 = 2 | n.i. | 99% of strains showed triclosan resistance; 32% chlorhexidine resistance; 100% susceptible to benzalkonium chloride | [127] |
| Chlorine | C. jejuni from poultry carcasses | MIC: 8 ppm for 97.5%; MBC: 128 ppm | Dose-dependent mechanisms, can be resuscitated after chlorine treatment. | n.i. | [129] |
| Antimicrobial mixtures based on sodium chloride, sodium hydroxide, citrus, oregano, and grape seed extracts | C. coli isolates (NC1, NC2, and NC3) and cecum strain (RC018) | MIC: 0.25–1% | n.i. | ↓ inhibit T6SS (hcp) | [20] |
| PinA | C. jejuni (81-176 and F38011) | MIC: 256 µg/mL | Membrane disruption, pore formation, increases membrane permeability. | n.i. | [130] |
| Disinfectant based on glutaraldehyde, formaldehyde, and antifreeze | C. fetus subsp. venerealis and C. fetus subsp. Fetus | MIC: 0.5% | n.i. | n.i. | [131] |
| Biocide | Strain | Concentration | Mechanism/Notes | Gene/Protein | Refs. |
|---|---|---|---|---|---|
| Chlorine QACs | S. enterica isolated from processing environments | Chlorine—500–1000 ppm QACs—3–25 ppm | n.i. | [135] | |
| Thymol | S. enterica serovar Typhimurium epidemic phage type DT193 strain | 1250 ppm total eradication | n.i. | [136] | |
| Ozonated water | S. Heidelberg multidrug-resistant isolated from pigs (farms) | MIC 4.4 mg/L | Induces changes in bacterial DNA after 10 min exposure. | n.i. | [137] |
| Sodium hypochlorite PAA Chlorhexidine | S. enterica serovar Minnesota strains isolated from a poultry slaughter, Brazilia | 1% SH (100%), 0.8% PAA, and 1% chlorhexidine decrease bacterial counts by 3.63 and 2.96 CFU/mL | n.i. | Highly virulent due to factors: avrA, agfA, lpfA, sodC, and luxS genes | [138] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Butucel, E.; Balta, I.; Ahmadi, M.; Dumitrescu, G.; Morariu, F.; Pet, I.; Stef, L.; Corcionivoschi, N. Biocides as Biomedicines against Foodborne Pathogenic Bacteria. Biomedicines 2022, 10, 379. https://doi.org/10.3390/biomedicines10020379
Butucel E, Balta I, Ahmadi M, Dumitrescu G, Morariu F, Pet I, Stef L, Corcionivoschi N. Biocides as Biomedicines against Foodborne Pathogenic Bacteria. Biomedicines. 2022; 10(2):379. https://doi.org/10.3390/biomedicines10020379
Chicago/Turabian StyleButucel, Eugenia, Igori Balta, Mirela Ahmadi, Gabi Dumitrescu, Florica Morariu, Ioan Pet, Lavinia Stef, and Nicolae Corcionivoschi. 2022. "Biocides as Biomedicines against Foodborne Pathogenic Bacteria" Biomedicines 10, no. 2: 379. https://doi.org/10.3390/biomedicines10020379
APA StyleButucel, E., Balta, I., Ahmadi, M., Dumitrescu, G., Morariu, F., Pet, I., Stef, L., & Corcionivoschi, N. (2022). Biocides as Biomedicines against Foodborne Pathogenic Bacteria. Biomedicines, 10(2), 379. https://doi.org/10.3390/biomedicines10020379