
Aspirin and Infection: A Narrative Review

 ,
,  ,
,  , , , , , , ,
, , , , , , ,
Abstract
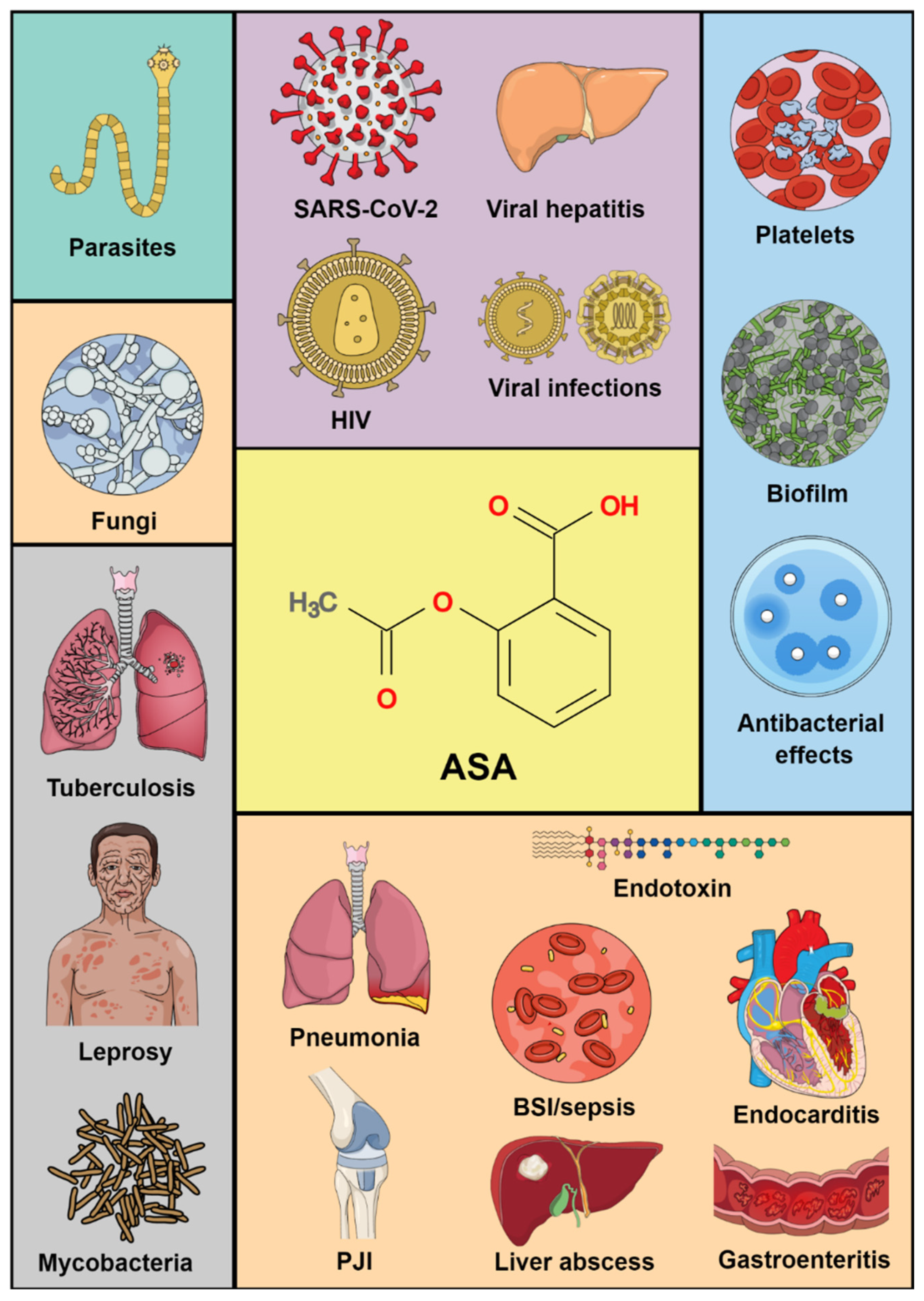
1. Introduction
2. Methods
3. Results
3.1. Aspirin, Platelets and Infections
3.2. Aspirin and Antibiofilm Effects
3.3. Aspirin and Bacterial Infections
3.3.1. Antibacterial General Effects
3.3.2. Aspirin and Endotoxin (Gram-Negative Lipopolysaccharide)
3.3.3. Aspirin and Bloodstream Infections/Sepsis
3.3.4. Aspirin and Endocarditis
3.3.5. Aspirin and Gastroenteritis/Liver Abscess
3.3.6. Aspirin and Prosthetic Joint Infections
3.3.7. Aspirin and Pneumonia
3.3.8. Aspirin and Mycobacterial Diseases
3.4. Aspirin and Fungal Infections
3.5. Aspirin and Parasitic Infections
3.6. Aspirin and Viral Infection
3.6.1. Aspirin and Human Immunodeficiency Virus Infection
3.6.2. Aspirin and SARS-CoV-2 Infection
3.6.3. Aspirin and Viral Hepatitis
3.6.4. Aspirin and Other Viral Infections
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Desborough, M.J.R.; Keeling, D.M. The Aspirin Story—From Willow to Wonder Drug. Br. J. Haematol. 2017, 177, 674–683. [Google Scholar] [CrossRef] [PubMed]
- Fuster, V.; Sweeny, J.M. Aspirin: A Historical and Contemporary Therapeutic Overview. Circulation 2011, 123, 768–778. [Google Scholar] [CrossRef] [PubMed]
- Piria, R. Sur la Salicine et les Produits de Sa Decomposition. Mémoire lu à l’Académie des Sciences de l’Institut de France le 26 Nov. 1838; Crochard: Paris, France, 1838. [Google Scholar]
- Thun, M.J.; Jacobs, E.J.; Patrono, C. The Role of Aspirin in Cancer Prevention. Nat. Rev. Clin. Oncol. 2012, 9, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Hannachi, N.; Baudoin, J.-P.; Prasanth, A.; Habib, G.; Camoin-Jau, L. The Distinct Effects of Aspirin on Platelet Aggregation Induced by Infectious Bacteria. Platelets 2020, 31, 1028–1038. [Google Scholar] [CrossRef]
- Chabert, A.; Damien, P.; Verhoeven, P.O.; Grattard, F.; Berthelot, P.; Zeni, F.; Panicot-Dubois, L.; Robert, S.; Dignat-George, F.; Eyraud, M.-A.; et al. Acetylsalicylic Acid Differentially Limits the Activation and Expression of Cell Death Markers in Human Platelets Exposed to Staphylococcus Aureus Strains. Sci. Rep. 2017, 7, 5610. [Google Scholar] [CrossRef]
- Anavekar, N.S.; Tleyjeh, I.M.; Anavekar, N.S.; Mirzoyev, Z.; Steckelberg, J.M.; Haddad, C.; Khandaker, M.H.; Wilson, W.R.; Chandrasekaran, K.; Baddour, L.M. Impact of Prior Antiplatelet Therapy on Risk of Embolism in Infective Endocarditis. Clin. Infect. Dis. 2007, 44, 1180–1186. [Google Scholar] [CrossRef]
- Chan, K.-L.; Dumesnil, J.G.; Cujec, B.; Sanfilippo, A.J.; Jue, J.; Turek, M.A.; Robinson, T.I.; Moher, D. Investigators of the Multicenter Aspirin Study in Infective Endocarditis. A Randomized Trial of Aspirin on the Risk of Embolic Events in Patients with Infective Endocarditis. J. Am. Coll. Cardiol. 2003, 42, 775–780. [Google Scholar] [CrossRef]
- Hannachi, N.; Habib, G.; Camoin-Jau, L. Aspirin Effect on—Platelet Interactions during Infectious Endocarditis. Front. Med. 2019, 6, 217. [Google Scholar] [CrossRef]
- Arman, M.; Krauel, K.; Tilley, D.O.; Weber, C.; Cox, D.; Greinacher, A.; Kerrigan, S.W.; Watson, S.P. Amplification of Bacteria-Induced Platelet Activation Is Triggered by FcγRIIA, Integrin αIIbβ3, and Platelet Factor 4. Blood 2014, 123, 3166–3174. [Google Scholar] [CrossRef]
- Zhang, G.; Han, J.; Welch, E.J.; Ye, R.D.; Voyno-Yasenetskaya, T.A.; Malik, A.B.; Du, X.; Li, Z. Lipopolysaccharide Stimulates Platelet Secretion and Potentiates Platelet Aggregation via TLR4/MyD88 and the cGMP-Dependent Protein Kinase Pathway. J. Immunol. 2009, 182, 7997–8004. [Google Scholar] [CrossRef]
- Ward, J.R.; Bingle, L.; Judge, H.M.; Brown, S.B.; Storey, R.F.; Whyte, M.K.B.; Dower, S.K.; Buttle, D.J.; Sabroe, I. Agonists of Toll-like Receptor (TLR)2 and TLR4 Are Unable to Modulate Platelet Activation by Adenosine Diphosphate and Platelet Activating Factor. Thromb. Haemost. 2005, 94, 831–838. [Google Scholar] [CrossRef] [PubMed]
- Waller, A.K.; Sage, T.; Kumar, C.; Carr, T.; Gibbins, J.M.; Clarke, S.R. Staphylococcus aureus Lipoteichoic Acid Inhibits Platelet Activation and Thrombus Formation via the Paf Receptor. J. Infect. Dis. 2013, 208, 2046–2057. [Google Scholar] [CrossRef] [PubMed]
- Svensson, L.; Baumgarten, M.; Mörgelin, M.; Shannon, O. Platelet Activation by Streptococcus pyogenes Leads to Entrapment in Platelet Aggregates, from Which Bacteria Subsequently Escape. Infect. Immun. 2014, 82, 4307–4314. [Google Scholar] [CrossRef] [PubMed]
- Johansson, D.; Shannon, O.; Rasmussen, M. Platelet and Neutrophil Responses to Gram Positive Pathogens in Patients with Bacteremic Infection. PLoS ONE 2011, 6, e26928. [Google Scholar] [CrossRef] [PubMed]
- Bayer, A.S.; Sullam, P.M.; Ramos, M.; Li, C.; Cheung, A.L.; Yeaman, M.R. Staphylococcus aureus Induces Platelet Aggregation via a Fibrinogen-Dependent Mechanism Which Is Independent of Principal Platelet Glycoprotein IIb/IIIa Fibrinogen-Binding Domains. Infect. Immun. 1995, 63, 3634–3641. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zhang, W.; Chen, M.; Zhang, J.; Zhang, M.; Dai, K. Aspirin Induces Platelet Apoptosis. Platelets 2013, 24, 637–642. [Google Scholar] [CrossRef]
- Nayak, M.K.; Dash, A.; Singh, N.; Dash, D. Aspirin Delimits Platelet Life Span by Proteasomal Inhibition. PLoS ONE 2014, 9, e105049. [Google Scholar] [CrossRef]
- Alem, M.A.S.; Douglas, L.J. Effects of Aspirin and Other Nonsteroidal Anti-Inflammatory Drugs on Biofilms and Planktonic Cells of Candida Albicans. Antimicrob. Agents Chemother. 2004, 48, 41–47. [Google Scholar] [CrossRef]
- Chan, A.K.Y.; Tsang, Y.C.; Chu, C.H.; Tsang, C.S.P. Aspirin as an Antifungal-Lock Agent in Inhibition of Candidal Biofilm Formation in Surgical Catheters. Infect. Drug Resist. 2021, 14, 1427–1433. [Google Scholar] [CrossRef]
- Abidi, S.H.; Ahmed, K.; Kazmi, S.U. The Antibiofilm Activity of Acetylsalicylic Acid, Mefenamic Acid, Acetaminophen against Biofilms Formed by P. aeruginosa and S. epidermidis. J. Pak. Med. Assoc. 2019, 69, 1493–1495. [Google Scholar] [CrossRef]
- Feng, W.; Yang, J.; Ma, Y.; Xi, Z.; Ren, Q.; Wang, S.; Ning, H. Aspirin and Verapamil Increase the Sensitivity of Candida albicans to Caspofungin under Planktonic and Biofilm Conditions. J. Glob. Antimicrob. Resist. 2021, 24, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Liao, Y.; Cong, L.; Lu, X.; Yang, R. In Vitro Interactions between Non-Steroidal Anti-Inflammatory Drugs and Antifungal Agents against Planktonic and Biofilm Forms of Trichosporon asahii. PLoS ONE 2016, 11, e0157047. [Google Scholar] [CrossRef] [PubMed]
- O’Grady, N.P.; Alexander, M.; Burns, L.A.; Dellinger, E.P.; Garland, J.; Heard, S.O.; Lipsett, P.A.; Masur, H.; Mermel, L.A.; Pearson, M.L.; et al. Guidelines for the Prevention of Intravascular Catheter-Related Infections. Am. J. Infect. Control 2011, 39, S1–S34. [Google Scholar] [CrossRef] [PubMed]
- Stepanović, S.; Vuković, D.; Jesić, M.; Ranin, L. Influence of Acetylsalicylic Acid (aspirin) on Biofilm Production by Candida Species. J. Chemother. 2004, 16, 134–138. [Google Scholar] [CrossRef]
- Muller, E.; Al-Attar, J.; Wolff, A.G.; Farber, B.F. Mechanism of Salicylate-Mediated Inhibition of Biofilm in Staphylococcus epidermidis. J. Infect. Dis. 1998, 177, 501–503. [Google Scholar] [CrossRef][Green Version]
- El-Mowafy, S.A.; Abd El Galil, K.H.; El-Messery, S.M.; Shaaban, M.I. Aspirin Is an Efficient Inhibitor of Quorum Sensing, Virulence and Toxins in Pseudomonas Aeruginosa. Microb. Pathog. 2014, 74, 25–32. [Google Scholar] [CrossRef]
- Dotto, C.; Lombarte Serrat, A.; Cattelan, N.; Barbagelata, M.S.; Yantorno, O.M.; Sordelli, D.O.; Ehling-Schulz, M.; Grunert, T.; Buzzola, F.R. The Active Component of Aspirin, Salicylic Acid, Promotes Biofilm Formation in a PIA-Dependent Manner. Front. Microbiol. 2017, 8, 4. [Google Scholar] [CrossRef]
- Jung, C.-J.; Yeh, C.-Y.; Shun, C.-T.; Hsu, R.-B.; Cheng, H.-W.; Lin, C.-S.; Chia, J.-S. Platelets Enhance Biofilm Formation and Resistance of Endocarditis-Inducing Streptococci on the Injured Heart Valve. J. Infect. Dis. 2012, 205, 1066–1075. [Google Scholar] [CrossRef]
- Wei, Y.P.; Chien, J.C.; Hsiang, W.H.; Yang, S.W.; Chen, C.Y. Aspirin Administration Might Accelerate the Subsidence of Periprosthetic Joint Infection. Sci. Rep. 2020, 10, 15967. [Google Scholar] [CrossRef]
- Bartzatt, R.; Cirillo, S.L.G.; Cirillo, J.D.; Donigan, L. Bifunctional Constructs of Aspirin and Ibuprofen (non-Steroidal Anti-Inflammatory Drugs; NSAIDs) That Express Antibacterial and Alkylation Activities. Biotechnol. Appl. Biochem. 2003, 37, 273–282. [Google Scholar] [CrossRef]
- Jones, D.L. Studies of Antimicrobial Action of Acetylsalicylic Acid. J. Am. Osteopath. Assoc. 1970, 69, 1020–1021. [Google Scholar] [PubMed]
- Jones, D.L. Antimicrobial Action of Acetylsalicylic Acid. II. Further Studies. J. Am. Osteopath. Assoc. 1971, 70, 1116–1118. [Google Scholar] [PubMed]
- Hockertz, S.; Paulini, I.; Rogalla, K.; Schettler, T. Influence of Acetylsalicylic Acid on a Listeria Monocytogenes Infection. Agents Actions 1993, 40, 119–123. [Google Scholar] [CrossRef]
- Yang, C.-Y.; Lin, Y.-P. Direct Antistaphylococcal Effects of High-Dose Aspirin in Hemodialysis Patients? Am. J. Kidney Dis. 2007, 50, 342. [Google Scholar] [CrossRef] [PubMed]
- Sedlacek, M.; Gemery, J.M.; Cheung, A.L.; Bayer, A.S.; Remillard, B.D. Aspirin Treatment Is Associated with a Significantly Decreased Risk of Staphylococcus aureus Bacteremia in Hemodialysis Patients with Tunneled Catheters. Am. J. Kidney Dis. 2007, 49, 401–408. [Google Scholar] [CrossRef]
- Chung, J.-G.; Tan, T.-W.; Tsai, H.-Y.; Hsieh, W.-T.; Chen, G.-W.; Lai, J.-M. Effects of Aspirin on Arylamine N-Acetyltransferase Activity in Klebsiella pneumoniae. Drug Chem. Toxicol. 1998, 21, 507–520. [Google Scholar] [CrossRef]
- Tiran, A.; Gruber, H.-J.; Graier, W.F.; Wagner, A.H.; Van Leeuwen, E.B.M.; Tiran, B. Aspirin Inhibits Chlamydia Pneumoniae-Induced Nuclear Factor-Kappa B Activation, Cytokine Expression, and Bacterial Development in Human Endothelial Cells. Arterioscler. Thromb. Vasc. Biol. 2002, 22, 1075–1080. [Google Scholar] [CrossRef]
- Yoneda, H.; Miura, K.; Matsushima, H.; Sugi, K.; Murakami, T.; Ouchi, K.; Yamashita, K.; Itoh, H.; Nakazawa, T.; Suzuki, M.; et al. Aspirin Inhibits Chlamydia pneumoniae-Induced NF-Kappa B Activation, Cyclo-Oxygenase-2 Expression and Prostaglandin E2 Synthesis and Attenuates Chlamydial Growth. J. Med. Microbiol. 2003, 52, 409–415. [Google Scholar] [CrossRef]
- Zimmermann, P.; Curtis, N. The Effect of Aspirin on Antibiotic Susceptibility. Expert Opin. Ther. Targets 2018, 22, 967–972. [Google Scholar] [CrossRef]
- Malla, C.F.; Mireles, N.A.; Ramírez, A.S.; Poveda, J.B.; Tavío, M.M. Aspirin, Sodium Benzoate and Sodium Salicylate Reverse Resistance to Colistin in Enterobacteriaceae and Pseudomonas aeruginosa. J. Antimicrob. Chemother. 2020, 75, 3568–3575. [Google Scholar] [CrossRef]
- Ngaini, Z.; Mortadza, N.A. Synthesis of Halogenated Azo-Aspirin Analogues from Natural Product Derivatives as the Potential Antibacterial Agents. Nat. Prod. Res. 2019, 33, 3507–3514. [Google Scholar] [CrossRef] [PubMed]
- Fajstavr, D.; Neznalová, K.; Kasálková, N.S.; Rimpelová, S.; Kubičíková, K.; Švorčík, V.; Slepička, P. Nanostructured Polystyrene Doped with Acetylsalicylic Acid and Its Antibacterial Properties. Materials 2020, 13, 3609. [Google Scholar] [CrossRef] [PubMed]
- Murphy, V.S.; Greenway, C.V. Aspirin and Pulmonary Lesions in Endotoxin Shock. Am. Heart J. 1972, 84, 581–582. [Google Scholar] [CrossRef]
- Halushka, P.V.; Wise, W.C.; Cook, J.A. Studies on the Beneficial Effects of Aspirin in Endotoxic Shock. Relationship to Inhibition of Arachidonic Acid Metabolism. Am. J. Med. 1983, 74, 91–96. [Google Scholar] [CrossRef]
- Feldhoff, C.M.; Gresele, P.; Pieters, G.; Vermylen, J. Acetylsalicylic Acid, BM 13.177 and Picotamide Improve the Survival of Endotoxin-Infused Rabbits. Thromb. Res. 1988, 52, 487–492. [Google Scholar] [CrossRef]
- Rao, P.S.; Cavanagh, D.; Gaston, L.W. Endotoxic Shock in the Primate: Effects of Aspirin and Dipyridamole Administration. Am. J. Obstet. Gynecol. 1981, 140, 914–922. [Google Scholar] [CrossRef]
- Jilma, B.; Blann, A.; Pernerstorfer, T.; Stohlawetz, P.; Eichler, H.G.; Vondrovec, B.; Amiral, J.; Richter, V.; Wagner, O.F. Regulation of Adhesion Molecules during Human Endotoxemia. No Acute Effects of Aspirin. Am. J. Respir. Crit. Care Med. 1999, 159, 857–863. [Google Scholar] [CrossRef]
- Derhaschnig, U.; Schweeger-Exeli, I.; Marsik, C.; Cardona, F.; Minuz, P.; Jilma, B. Effects of Aspirin and NO-Aspirin (NCX 4016) on Platelet Function and Coagulation in Human Endotoxemia. Platelets 2010, 21, 320–328. [Google Scholar] [CrossRef]
- Hamid, U.; Krasnodembskaya, A.; Fitzgerald, M.; Shyamsundar, M.; Kissenpfennig, A.; Scott, C.; Lefrancais, E.; Looney, M.R.; Verghis, R.; Scott, J.; et al. Aspirin Reduces Lipopolysaccharide-Induced Pulmonary Inflammation in Human Models of ARDS. Thorax 2017, 72, 971–980. [Google Scholar] [CrossRef]
- Kiers, D.; van der Heijden, W.A.; van Ede, L.; Gerretsen, J.; de Mast, Q.; van der Ven, A.J.; El Messaoudi, S.; Rongen, G.A.; Gomes, M.; Kox, M.; et al. A Randomised Trial on the Effect of Anti-Platelet Therapy on the Systemic Inflammatory Response in Human Endotoxaemia. Thromb. Haemost. 2017, 117, 1798–1807. [Google Scholar] [CrossRef]
- Leijte, G.P.; Kiers, D.; van der Heijden, W.; Jansen, A.; Gerretsen, J.; Boerrigter, V.; Netea, M.G.; Kox, M.; Pickkers, P. Treatment with Acetylsalicylic Acid Reverses Endotoxin Tolerance in Humans In Vivo: A Randomized Placebo-Controlled Study. Crit. Care Med. 2019, 47, 508–516. [Google Scholar] [CrossRef] [PubMed]
- Dalldorf, F.G.; Jennette, J.C.; Upton, G.W.; Tullar, J.C. Experimental Meningococcal Septicemia. Effect of Aspirin Therapy. Arch. Pathol. Lab. Med. 1978, 102, 515–517. [Google Scholar] [PubMed]
- Carestia, A.; Davis, R.P.; Grosjean, H.; Lau, M.W.; Jenne, C.N. Acetylsalicylic Acid Inhibits Intravascular Coagulation during Staphylococcus Aureus-Induced Sepsis in Mice. Blood 2020, 135, 1281–1286. [Google Scholar] [CrossRef] [PubMed]
- Osthoff, M.; Sidler, J.A.; Lakatos, B.; Frei, R.; Dangel, M.; Weisser, M.; Battegay, M.; Widmer, A.F. Low-Dose Acetylsalicylic Acid Treatment and Impact on Short-Term Mortality in Staphylococcus aureus Bloodstream Infection: A Propensity Score-Matched Cohort Study. Crit. Care Med. 2016, 44, 773–781. [Google Scholar] [CrossRef]
- Trauer, J.; Muhi, S.; McBryde, E.S.; Al Harbi, S.A.; Arabi, Y.M.; Boyle, A.J.; Cartin-Ceba, R.; Chen, W.; Chen, Y.-T.; Falcone, M.; et al. Quantifying the Effects of Prior Acetyl-Salicylic Acid on Sepsis-Related Deaths: An Individual Patient Data Meta-Analysis Using Propensity Matching. Crit. Care Med. 2017, 45, 1871–1879. [Google Scholar] [CrossRef]
- Hsu, J.; Donnelly, J.P.; Chaudhary, N.S.; Moore, J.X.; Safford, M.M.; Kim, J.; Wang, H.E. Aspirin Use and Long-Term Rates of Sepsis: A Population-Based Cohort Study. PLoS ONE 2018, 13, e0194829. [Google Scholar] [CrossRef]
- Eisen, D.P.; Leder, K.; Woods, R.L.; Lockery, J.E.; McGuinness, S.L.; Wolfe, R.; Pilcher, D.; Moore, E.M.; Shastry, A.; Nelson, M.R.; et al. Effect of Aspirin on Deaths Associated with Sepsis in Healthy Older People (ANTISEPSIS): A Randomised, Double-Blind, Placebo-Controlled Primary Prevention Trial. Lancet Respir. Med. 2021, 9, 186–195. [Google Scholar] [CrossRef]
- Levison, M.E.; Carrizosa, J.; Tanphaichitra, D.; Schick, P.K.; Rubin, W. Effect of Aspirin on Thrombogenesis and on Production of Experimental Aortic Valvular Streptococcus Viridans Endocarditis in Rabbits. Blood 1977, 49, 645–650. [Google Scholar] [CrossRef]
- Nicolau, D.P.; Freeman, C.D.; Nightingale, C.H.; Quintiliani, R.; Coe, C.J.; Maderazo, E.G.; Cooper, B.W. Reduction of Bacterial Titers by Low-Dose Aspirin in Experimental Aortic Valve Endocarditis. Infect. Immun. 1993, 61, 1593–1595. [Google Scholar] [CrossRef]
- Taha, T.H.; Durrant, S.S.; Mazeika, P.K.; Nihoyannopoulos, P.; Oakley, C.M. Aspirin to Prevent Growth of Vegetations and Cerebral Emboli in Infective Endocarditis. J. Intern. Med. 1992, 231, 543–546. [Google Scholar] [CrossRef]
- Nicolau, D.P.; Marangos, M.N.; Nightingale, C.H.; Quintiliani, R. Influence of Aspirin on Development and Treatment of Experimental Staphylococcus aureus Endocarditis. Antimicrob. Agents Chemother. 1995, 39, 1748–1751. [Google Scholar] [CrossRef] [PubMed]
- Kupferwasser, L.I.; Yeaman, M.R.; Shapiro, S.M.; Nast, C.C.; Sullam, P.M.; Filler, S.G.; Bayer, A.S. Acetylsalicylic Acid Reduces Vegetation Bacterial Density, Hematogenous Bacterial Dissemination, and Frequency of Embolic Events in Experimental Staphylococcus aureus Endocarditis through Antiplatelet and Antibacterial Effects. Circulation 1999, 99, 2791–2797. [Google Scholar] [CrossRef] [PubMed]
- Eisen, D.P.; McBryde, E.S. An Association Between Aspirin Use in Human Cases of Infective Endocarditis and Reduced Systemic Embolism Is Shown in Meta-Analysis of Observational Studies. J. Infect. Dis. 2015, 212, 673–674. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Cahill, T.J.; Prendergast, B.D. Reply: Aspirin for the Prevention of Infective Endocarditis? J. Am. Coll. Cardiol. 2017, 70, 1105. [Google Scholar] [CrossRef] [PubMed]
- Madias, J.E. Readers’ Comments: Aspirin And/or Other Antiplatelet Agents for the Prevention of Infective Endocarditis. Am. J. Cardiol. 2020, 125, 1450–1451. [Google Scholar] [CrossRef]
- Farris, R.K.; Tapper, E.J.; Powell, D.W.; Morris, S.M. Effect of Aspirin on Normal and Cholera Toxin-Stimulated Intestinal Electrolyte Transport. J. Clin. Investig. 1976, 57, 916–924. [Google Scholar] [CrossRef]
- Powell, D.W.; Tapper, E.J.; Morris, S.M. Aspirin-Stimulated Intestinal Electrolyte Transport in Rabbit Ileum in Vitro. Gastroenterology 1979, 76, 1429–1437. [Google Scholar] [CrossRef]
- Finck, A.D.; Katz, R.L. Prevention of Cholera-Induced Intestinal Secretion in the Cat by Aspirin. Nature 1972, 238, 273–274. [Google Scholar] [CrossRef]
- Nalin, D.R. Letter: Failure of Aspirin to Reverse Intestinal Secretion after Cholera Toxin in Dogs. Lancet 1976, 2, 576. [Google Scholar] [CrossRef]
- Burke, V.; Gracey, M.; Suharyono, W.; Sunoto, S. Reduction by Aspirin of Intestinal Fluid-Loss in Acute Childhood Gastroenteritis. Lancet 1980, 1, 1329–1330. [Google Scholar] [CrossRef]
- Gracey, M.; Phadke, M.A.; Burke, V.; Raut, S.K.; Singh, B. Aspirin in Acute Gastroenteritis: A Clinical and Microbiological Study. J. Pediatr. Gastroenterol. Nutr. 1984, 3, 692–695. [Google Scholar] [CrossRef] [PubMed]
- Castor, B.; Thorén, A.; Barkenius, G. Failure of Aspirin in Symptomatic Treatment of Acute Diarrhoea. J. Diarrhoeal Dis. Res. 1991, 9, 29–32. [Google Scholar] [PubMed]
- Islam, A.; Bardhan, P.K.; Islam, M.R.; Rahman, M. A Randomized Double Blind Trial of Aspirin versus Placebo in Cholera and Non-Cholera Diarrhoea. Trop. Geogr. Med. 1986, 38, 221–225. [Google Scholar] [PubMed]
- Domenico, P.; Schwartz, S.; Cunha, B.A. Reduction of Capsular Polysaccharide Production in Klebsiella pneumoniae by Sodium Salicylate. Infect. Immun. 1989, 57, 3778–3782. [Google Scholar] [CrossRef]
- Lee, C.-H.; Su, L.-H.; Liu, J.-W.; Chang, C.-C.; Chen, R.-F.; Yang, K.-D. Aspirin Enhances Opsonophagocytosis and Is Associated to a Lower Risk for Klebsiella pneumoniae Invasive Syndrome. BMC Infect. Dis. 2014, 14, 47. [Google Scholar] [CrossRef]
- Liu, J.-S.; Lee, C.-H.; Chuah, S.-K.; Tai, W.-C.; Chang, C.-C.; Chen, F.-J. Aspirin Use Is Associated with Reduced Risk for Recurrence of Pyogenic Liver Abscess: A Propensity Score Analysis. Sci. Rep. 2019, 9, 11511. [Google Scholar] [CrossRef]
- Tai, C.-H.; Hsu, C.-N.; Yang, S.-C.; Wu, C.-K.; Liang, C.-M.; Tai, W.-C.; Chuah, S.-K.; Lee, C.-H. The Impact of Aspirin on Klebsiella pneumoniae Liver Abscess in Diabetic Patients. Sci. Rep. 2020, 10, 21329. [Google Scholar] [CrossRef]
- Huang, R.; Buckley, P.S.; Scott, B.; Parvizi, J.; Purtill, J.J. Administration of Aspirin as a Prophylaxis Agent against Venous Thromboembolism Results in Lower Incidence of Periprosthetic Joint Infection. J. Arthroplast. 2015, 30, 39–41. [Google Scholar] [CrossRef]
- Jiang, Y.; Wang, S.-N.; Wu, H.-T.; Qin, H.-J.; Ren, M.-L.; Lin, J.-C.; Yu, B. Aspirin Alleviates Orthopedic Implant-associated Infection. Int. J. Mol. Med. 2019, 44, 1281–1288. [Google Scholar] [CrossRef]
- Eyre, P.; Nymeyer, D.H.; McCraw, B.M.; Deline, T.R. Protection by Acetylsalicylic Acid and Other Agents in Experimental Acute Interstitial Pneumonia of Calves. Vet. Rec. 1976, 98, 64–66. [Google Scholar] [CrossRef]
- Light, R.B. Indomethacin and Acetylsalicylic Acid Reduce Intrapulmonary Shunt in Experimental Pneumococcal Pneumonia. Am. Rev. Respir. Dis. 1986, 134, 520–525. [Google Scholar] [PubMed]
- Ferrer, M.; Torres, A.; Baer, R.; Hernández, C.; Roca, J.; Rodriguez-Roisin, R. Effect of Acetylsalicylic Acid on Pulmonary Gas Exchange in Patients with Severe Pneumonia. Chest 1997, 111, 1094–1100. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Anthony, D.; Yatmaz, S.; Wijburg, O.; Satzke, C.; Levy, B.; Vlahos, R.; Bozinovski, S. Aspirin-Triggered Resolvin D1 Reduces Pneumococcal Lung Infection and Inflammation in a Viral and Bacterial Coinfection Pneumonia Model. Clin. Sci. 2017, 131, 2347–2362. [Google Scholar] [CrossRef] [PubMed]
- Abdulnour, R.E.; Sham, H.P.; Douda, D.N.; Colas, R.A.; Dalli, J.; Bai, Y.; Ai, X.; Serhan, C.N.; Levy, B.D. Aspirin-Triggered Resolvin D1 Is Produced during Self-Resolving Gram-Negative Bacterial Pneumonia and Regulates Host Immune Responses for the Resolution of Lung Inflammation. Mucosal Immunol. 2016, 9, 1278–1287. [Google Scholar] [CrossRef] [PubMed]
- Esposito, A.L. Aspirin Impairs Antibacterial Mechanisms in Experimental Pneumococcal Pneumonia. Am. Rev. Respir. Dis. 1984, 130, 857–862. [Google Scholar] [PubMed]
- Kolditz, M.; Welte, T. Preventing Cardiovascular Events after Pneumonia with Aspirin: One Step Forward, but Still Many to Go. Eur. Respir. J. 2021, 57, 2003778. [Google Scholar] [CrossRef] [PubMed]
- Falcone, M.; Russo, A.; Cangemi, R.; Farcomeni, A.; Calvieri, C.; Barillà, F.; Scarpellini, M.G.; Bertazzoni, G.; Palange, P.; Taliani, G.; et al. Lower Mortality Rate in Elderly Patients with Community-Onset Pneumonia on Treatment with Aspirin. J. Am. Heart Assoc. 2015, 4, e001595. [Google Scholar] [CrossRef]
- Oz, F.; Gul, S.; Kaya, M.G.; Yazici, M.; Bulut, I.; Elitok, A.; Ersin, G.; Abakay, O.; Akkoyun, C.D.; Oncul, A.; et al. Does Aspirin Use Prevent Acute Coronary Syndrome in Patients with Pneumonia: Multicenter Prospective Randomized Trial. Coron. Artery Dis. 2013, 24, 231–237. [Google Scholar] [CrossRef]
- Hamilton, F.; Arnold, D.; Henley, W.; Payne, R.A. Aspirin Reduces Cardiovascular Events in Patients with Pneumonia: A Prior Event Rate Ratio Analysis in a Large Primary Care Database. Eur. Respir. J. 2021, 57, 2002795. [Google Scholar] [CrossRef]
- Chen, Y.-C.; Chen, Y.-Y.; Yeh, H.W.; Yeh, T.-Y.; Huang, J.-Y.; Liao, P.-L.; Yeh, L.-T.; Yang, S.-F.; Chou, M.-C.; Yeh, C.-B. Association between Aspirin Use and Decreased Risk of Pneumonia in Patients with Cardio-Cerebra-Vascular Ischemic Disease: A Population-Based Cohort Study. Front. Public Health 2021, 9, 625834. [Google Scholar] [CrossRef]
- Falcone, M.; Russo, A.; Farcomeni, A.; Pieralli, F.; Vannucchi, V.; Vullo, V.; Violi, F.; Venditti, M. Septic Shock from Community-Onset Pneumonia: Is There a Role for Aspirin plus Macrolides Combination? Intensive Care Med. 2016, 42, 301–302. [Google Scholar] [CrossRef] [PubMed]
- Falcone, M.; Russo, A.; Shindo, Y.; Farcomeni, A.; Pieralli, F.; Cangemi, R.; Liu, J.; Xia, J.; Okumura, J.; Sano, M.; et al. A Hypothesis-Generating Study of the Combination of Aspirin plus Macrolides in Patients with Severe Community-Acquired Pneumonia. Antimicrob. Agents Chemother. 2019, 63, e01556-18. [Google Scholar] [CrossRef] [PubMed]
- Misra, U.K.; Kalita, J.; Nair, P.P. Role of Aspirin in Tuberculous Meningitis: A Randomized Open Label Placebo Controlled Trial. J. Neurol. Sci. 2010, 293, 12–17. [Google Scholar] [CrossRef]
- Misra, U.K.; Kalita, J.; Sagar, B.; Bhoi, S.K. Does Adjunctive Corticosteroid and Aspirin Therapy Improve the Outcome of Tuberculous Meningitis? Neurol. India 2018, 66, 1672–1677. [Google Scholar] [PubMed]
- Rizvi, I.; Garg, R.K.; Malhotra, H.S.; Kumar, N.; Uniyal, R. Role of Aspirin in Tuberculous Meningitis: A Systematic Review and Meta-Analysis. Neurol. India 2019, 67, 993–1002. [Google Scholar] [PubMed]
- Kroesen, V.M.; Rodríguez-Martínez, P.; García, E.; Rosales, Y.; Díaz, J.; Martín-Céspedes, M.; Tapia, G.; Sarrias, M.R.; Cardona, P.-J.; Vilaplana, C. A Beneficial Effect of Low-Dose Aspirin in a Murine Model of Active Tuberculosis. Front. Immunol. 2018, 9, 798. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Du, Z.; Ni, M.; Wang, Z.; Liang, M.; Sheng, H.; Zhang, A.; Yang, J. Aspirin Enhances the Clinical Efficacy of Anti-Tuberculosis Therapy in Pulmonary Tuberculosis in Patients with Type 2 Diabetes Mellitus. Infect. Dis. 2020, 52, 721–729. [Google Scholar] [CrossRef]
- Klenerman, P. Prostaglandins and Leprosy. A Role for Aspirin? Lepr. Rev. 1989, 60, 51–58. [Google Scholar] [CrossRef]
- Garg, S.K.; Kumar, B.; Shukla, V.K.; Bakaya, V.; Lal, R.; Kaur, S. Pharmacokinetics of Aspirin and Chloramphenicol in Normal and Leprotic Patients before and after Dapsone Therapy. Int. J. Clin. Pharmacol. Ther. Toxicol. 1988, 26, 204–205. [Google Scholar]
- Kar, H.K.; Roy, R.G. Comparison of Colchicine and Aspirin in the Treatment of Type 2 Lepra Reaction. Lepr. Rev. 1988, 59, 201–203. [Google Scholar] [CrossRef]
- Sharma, P.K.; Singh, D.K. A Traditional NSAID Aspirin along with Clofazimine in Erythema Nodosum Leprosum Reaction: Study of Six Cases. Indian J. Lepr. 2016, 87, 249–253. [Google Scholar] [PubMed]
- Kock, J.L.F.; Sebolai, O.M.; Pohl, C.H.; van Wyk, P.W.J.; Lodolo, E.J. Oxylipin Studies Expose Aspirin as Antifungal. FEMS Yeast Res. 2007, 7, 1207–1217. [Google Scholar] [CrossRef] [PubMed]
- Sebolai, O.M.; Pohl, C.H.; Botes, P.J.; van Wyk, P.W.J.; Mzizi, R.; Swart, C.W.; Kock, J.L.F. Distribution of 3-Hydroxy Oxylipins and Acetylsalicylic Acid Sensitivity in Cryptococcus Species. Can. J. Microbiol. 2008, 54, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Leeuw, N.J.; Swart, C.W.; Ncango, D.M.; Pohl, C.H.; Sebolai, O.M.; Strauss, C.J.; Botes, P.J.; van Wyk, P.W.J.; Nigam, S.; Kock, J.L.F. Acetylsalicylic Acid as Antifungal in Eremothecium and Other Yeasts. Antonie Van Leeuwenhoek 2007, 91, 393–405. [Google Scholar] [CrossRef]
- Ogundeji, A.O.; Mjokane, N.; Folorunso, O.S.; Pohl, C.H.; Nyaga, M.M.; Sebolai, O.M. The Repurposing of Acetylsalicylic Acid as a Photosensitiser to Inactivate the Growth of Cryptococcal Cells. Pharmaceuticals 2021, 14, 404. [Google Scholar] [CrossRef]
- Trofa, D.; Agovino, M.; Stehr, F.; Schäfer, W.; Rykunov, D.; Fiser, A.; Hamari, Z.; Nosanchuk, J.D.; Gácser, A. Acetylsalicylic Acid (aspirin) Reduces Damage to Reconstituted Human Tissues Infected with Candida Species by Inhibiting Extracellular Fungal Lipases. Microbes Infect. 2009, 11, 1131–1139. [Google Scholar] [CrossRef][Green Version]
- Pereira, P.A.T.; Bini, D.; Bovo, F.; Faccioli, L.H.; Monteiro, M.C. Neutrophils Influx and Proinflammatory Cytokines Inhibition by Sodium Salicylate, Unlike Aspirin, in Candida albicans-Induced Peritonitis Model. Folia Microbiol. 2016, 61, 337–346. [Google Scholar] [CrossRef]
- Pérez-Molina, J.A.; Molina, I. Chagas Disease. Lancet 2018, 391, 82–94. [Google Scholar] [CrossRef]
- Shikanai-Yasuda, M.A.; Carvalho, N.B. Oral Transmission of Chagas Disease. Clin. Infect. Dis. 2012, 54, 845–852. [Google Scholar] [CrossRef]
- Cossentini, L.A.; Da Silva, R.V.; Yamada-Ogatta, S.F.; Yamauchi, L.M.; De Almeida Araújo, E.J.; Pinge-Filho, P. Aspirin Treatment Exacerbates Oral Infections by Trypanosoma cruzi. Exp. Parasitol. 2016, 164, 64–70. [Google Scholar] [CrossRef]
- Silvero-Isidre, A.; Morínigo-Guayuán, S.; Meza-Ojeda, A.; Mongelós-Cardozo, M.; Centurión-Wenninger, C.; Figueredo-Thiel, S.; Sanchez, D.F.; Acosta, N. Protective Effect of Aspirin Treatment on Mouse Behavior in the Acute Phase of Experimental Infection with Trypanosoma cruzi. Parasitol. Res. 2018, 117, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Oda, J.Y.; Belém, M.O.; Carlos, T.M.; Gouveia, R.; Luchetti, B.F.C.; Moreira, N.M.; Massocatto, C.L.; Araújo, S.M.; Sant Ana, D.M.G.; Buttow, N.C.; et al. Myenteric Neuroprotective Role of Aspirin in Acute and Chronic Experimental Infections with Trypanosoma cruzi. Neurogastroenterol. Motil. 2017, 29, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Souza, N.D.D.E.; Belin, B.S.; Massocatto, C.L.; Araújo, S.M.D.E.; Sant’Ana, D.M.G.; Araújo, E.J.A.; Filho, P.; Nihei, O.K.; Moreira, N.M. Effect of Acetylsalicylic Acid on Total Myenteric Neurons in Mice Experimentally Infected with Trypanosoma cruzi. An. Acad. Bras. Cienc. 2019, 91, e20180389. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Machado, F.S.; Huang, H.; Oz, H.S.; Jelicks, L.A.; Prado, C.M.; Koba, W.; Fine, E.J.; Zhao, D.; Factor, S.M.; et al. Aspirin Treatment of Mice Infected with Trypanosoma cruzi and Implications for the Pathogenesis of Chagas Disease. PLoS ONE 2011, 6, e16959. [Google Scholar] [CrossRef] [PubMed]
- Molina-Berríos, A.; Campos-Estrada, C.; Lapier, M.; Duaso, J.; Kemmerling, U.; Galanti, N.; Ferreira, J.; Morello, A.; López-Muñoz, R.; Maya, J.D. Protection of Vascular Endothelium by Aspirin in a Murine Model of Chronic Chagas’ Disease. Parasitol. Res. 2013, 112, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Ricci, M.F.; Béla, S.R.; Moraes, M.M.; Bahia, M.T.; Mazzeti, A.L.; Oliveira, A.C.S.; Andrade, L.O.; Radí, R.; Piacenza, L.; Arantes, R.M.E. Neuronal Parasitism, Early Myenteric Neurons Depopulation and Continuous Axonal Networking Damage as Underlying Mechanisms of the Experimental Intestinal Chagas’ Disease. Front. Cell. Infect. Microbiol. 2020, 10, 583899. [Google Scholar] [CrossRef]
- Massocatto, C.L.; Martins Moreira, N.; Muniz, E.; Marques de Araújo, S.; Pinge-Filho, P.; Rossi, R.M.; de Almeida Araújo, E.J.; de Mello Gonçales Sant’Ana, D. Treatment with Low Doses of Aspirin during Chronic Phase of Experimental Chagas’ Disease Increases Oesophageal Nitrergic Neuronal Subpopulation in Mice. Int. J. Exp. Pathol. 2017, 98, 356–362. [Google Scholar] [CrossRef]
- Massocatto, C.L.; Moreira, N.M.; Muniz, E.; Pinge-Filho, P.; Rossi, R.M.; de Araújo, E.J.A.; de Sant’Ana, D.M.G. Aspirin Prevents Atrophy of Esophageal Nitrergic Myenteric Neurons in a Mouse Model of Chronic Chagas Disease. Dis. Esophagus 2017, 30, 1–8. [Google Scholar] [CrossRef]
- López-Muñoz, R.; Faúndez, M.; Klein, S.; Escanilla, S.; Torres, G.; Lee-Liu, D.; Ferreira, J.; Kemmerling, U.; Orellana, M.; Morello, A.; et al. Trypanosoma cruzi: In Vitro Effect of Aspirin with Nifurtimox and Benznidazole. Exp. Parasitol. 2010, 124, 167–171. [Google Scholar] [CrossRef]
- Pereira, R.S.; Malvezi, A.D.; Lovo-Martins, M.I.; Lucchetti, B.F.C.; Santos, J.P.; Tavares, E.R.; Verri, W.A., Jr.; de Almeida Araújo, E.J.; Yamauchi, L.M.; Yamada-Ogatta, S.F.; et al. Combination Therapy Using Benznidazole and Aspirin during the Acute Phase of Experimental Chagas Disease Prevents Cardiovascular Dysfunction and Decreases Typical Cardiac Lesions in the Chronic Phase. Antimicrob. Agents Chemother. 2020, 64, e000069-20. [Google Scholar] [CrossRef]
- Hemmer, C.J.; Kern, P.; Holst, F.G.; Nawroth, P.P.; Dietrich, M. Neither Heparin nor Acetylsalicylic Acid Influence the Clinical Course in Human Plasmodium Falciparum Malaria: A Prospective Randomized Study. Am. J. Trop. Med. Hyg. 1991, 45, 608–612. [Google Scholar] [CrossRef] [PubMed]
- Gryseels, B.; Polman, K.; Clerinx, J.; Kestens, L. Human Schistosomiasis. Lancet 2006, 368, 1106–1118. [Google Scholar] [CrossRef]
- Feitosa, K.A.; Zaia, M.G.; Rodrigues, V.; Castro, C.A.; de Correia, R.O.; Pinto, F.G.; Rossi, K.N.Z.P.; Avó, L.R.S.; Afonso, A.; Anibal, F.F. Associated with Acetylsalicylic Acid and Their Relation to the Hepatic Fibrosis in Infected Mice. Front. Pharmacol. 2017, 8, 1000. [Google Scholar] [CrossRef] [PubMed]
- Kaewpitoon, N.; Kaewpitoon, S.-J.; Pengsaa, P. Opisthorchiasis in Thailand: Review and Current Status. World J. Gastroenterol. 2008, 14, 2297–2302. [Google Scholar] [CrossRef] [PubMed]
- Sudsarn, P.; Boonmars, T.; Ruangjirachuporn, W.; Namwat, N.; Loilome, W.; Sriraj, P.; Aukkanimart, R.; Nadchanan, W.; Jiraporn, S. Combination of Praziquantel and Aspirin Minimizes Liver Pathology of Hamster Opisthorchis viverrini Infection Associated Cholangiocarcinoma. Pathol. Oncol. Res. 2016, 22, 57–65. [Google Scholar] [CrossRef]
- Currier, J.S.; Lundgren, J.D.; Carr, A.; Klein, D.; Sabin, C.A.; Sax, P.E.; Schouten, J.T.; Smieja, M. Working Group 2 Epidemiological Evidence for Cardiovascular Disease in HIV-Infected Patients and Relationship to Highly Active Antiretroviral Therapy. Circulation 2008, 118, e29–e35. [Google Scholar] [CrossRef]
- Ryom, L.; Cotter, A.; De Miguel, R.; Béguelin, C.; Podlekareva, D.; Arribas, J.R.; Marzolini, C.; Mallon, P.; Rauch, A.; Kirk, O.; et al. 2019 Update of the European AIDS Clinical Society Guidelines for Treatment of People Living with HIV Version 10.0. HIV Med. 2020, 21, 617–624. [Google Scholar] [CrossRef]
- Pereira, C.F.; Paridaen, J.T.M.L.; Rutten, K.; Huigen, M.C.D.G.; van de Bovenkamp, M.; Middel, J.; Beerens, N.; Berkhout, B.; Schuurman, R.; Marnett, L.J.; et al. Aspirin-like Molecules That Inhibit Human Immunodeficiency Virus 1 Replication. Antivir. Res. 2003, 58, 253–263. [Google Scholar] [CrossRef]
- O’Brien, M.; Montenont, E.; Hu, L.; Nardi, M.A.; Valdes, V.; Merolla, M.; Gettenberg, G.; Cavanagh, K.; Aberg, J.A.; Bhardwaj, N.; et al. Aspirin Attenuates Platelet Activation and Immune Activation in HIV-1-Infected Subjects on Antiretroviral Therapy: A Pilot Study. J. Acquir. Immune Defic. Syndr. 2013, 63, 280–288. [Google Scholar] [CrossRef]
- O’Brien, M.P.; Hunt, P.W.; Kitch, D.W.; Klingman, K.; Stein, J.H.; Funderburg, N.T.; Berger, J.S.; Tebas, P.; Clagett, B.; Moisi, D.; et al. A Randomized Placebo Controlled Trial of Aspirin Effects on Immune Activation in Chronically Human Immunodeficiency Virus-Infected Adults on Virologically Suppressive Antiretroviral Therapy. Open Forum Infect. Dis. 2017, 4, ofw278. [Google Scholar] [CrossRef]
- Blanchard, H.C.; Taha, A.Y.; Rapoport, S.I.; Yuan, Z.-X. Low-Dose Aspirin (acetylsalicylate) Prevents Increases in Brain PGE2, 15-Epi-Lipoxin A4 and 8-Isoprostane Concentrations in 9 Month-Old HIV-1 Transgenic Rats, a Model for HIV-1 Associated Neurocognitive Disorders. Prostaglandins Leukot. Essent. Fatty Acids 2015, 96, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Marcus, J.L.; Neugebauer, R.S.; Leyden, W.A.; Chao, C.R.; Xu, L.; Quesenberry, C.P., Jr.; Klein, D.B.; Towner, W.J.; Horberg, M.A.; Silverberg, M.J. Use of Abacavir and Risk of Cardiovascular Disease among HIV-Infected Individuals. J. Acquir. Immune Defic. Syndr. 2016, 71, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Falcinelli, E.; Francisci, D.; Schiaroli, E.; Minuz, P.; Orsini, S.; Malincarne, L.; Sebastiano, M.; Mezzasoma, A.M.; Pasticci, M.B.; Guglielmini, G.; et al. Effect of Aspirin Treatment on Abacavir-Associated Platelet Hyperreactivity in HIV-Infected Patients. Int. J. Cardiol. 2018, 263, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Maggi, P.; De Socio, G.V.; Cicalini, S.; D’Abbraccio, M.; Dettorre, G.; Di Biagio, A.; Martinelli, C.; Nunnari, G.; Rusconi, S.; Sighinolfi, L.; et al. Statins and Aspirin in the Prevention of Cardiovascular Disease among HIV-Positive Patients between Controversies and Unmet Needs: Review of the Literature and Suggestions for a Friendly Use. AIDS Res. Ther. 2019, 16, 11. [Google Scholar] [CrossRef]
- Santos Junior, G.G.D.; Araújo, P.S.R.; Leite, K.M.E.; Godoi, E.T.; Vasconcelos, A.F.; Lacerda, H.R. The Effect of Atorvastatin + Aspirin on the Endothelial Function Differs with Age in Patients with HIV: A Case-Control Study. Arq. Bras. Cardiol. 2021, 117, 365–375. [Google Scholar]
- Weekly Epidemiological Update on COVID-19—30 November 2021. Available online: https://www.who.int/publications/m/item/weekly-epidemiological-update-on-covid-19---30-november-2021 (accessed on 9 December 2021).
- Glatthaar-Saalmüller, B.; Mair, K.H.; Saalmüller, A. Antiviral Activity of Aspirin against RNA Viruses of the Respiratory Tract-an in Vitro Study. Influenza Other Respir. Viruses 2017, 11, 85–92. [Google Scholar] [CrossRef]
- Rizk, J.G.; Lavie, C.J.; Gupta, A. Low-Dose Aspirin for Early COVID-19: Does the Early Bird Catch the Worm? Expert Opin. Investig. Drugs 2021, 30, 785–788. [Google Scholar] [CrossRef]
- Wijaya, I.; Andhika, R.; Huang, I.; Purwiga, A.; Budiman, K.Y. The Effects of Aspirin on the Outcome of COVID-19: A Systematic Review and Meta-Analysis. Clin. Epidemiol. Glob. Health 2021, 12, 100883. [Google Scholar] [CrossRef]
- Martha, J.W.; Pranata, R.; Lim, M.A.; Wibowo, A.; Akbar, M.R. Active Prescription of Low-Dose Aspirin during or prior to Hospitalization and Mortality in COVID-19: A Systematic Review and Meta-Analysis of Adjusted Effect Estimates. Int. J. Infect. Dis. 2021, 108, 6–12. [Google Scholar] [CrossRef]
- Kow, C.S.; Hasan, S.S. Use of Antiplatelet Drugs and the Risk of Mortality in Patients with COVID-19: A Meta-Analysis. J. Thromb. Thrombolysis 2021, 52, 124–129. [Google Scholar] [CrossRef]
- Salah, H.M.; Mehta, J.L. Meta-Analysis of the Effect of Aspirin on Mortality in COVID-19. Am. J. Cardiol. 2021, 142, 158–159. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, R.; Kumar, A. Use of Aspirin in Reduction of Mortality of COVID-19 Patients: A Meta-Analysis. Int. J. Clin. Pract. 2021, 75, e14515. [Google Scholar] [CrossRef] [PubMed]
- Connors, J.M.; Brooks, M.M.; Sciurba, F.C.; Krishnan, J.A.; Bledsoe, J.R.; Kindzelski, A.; Baucom, A.L.; Kirwan, B.-A.; Eng, H.; Martin, D.; et al. Effect of Antithrombotic Therapy on Clinical Outcomes in Outpatients with Clinically Stable Symptomatic COVID-19: The ACTIV-4B Randomized Clinical Trial. JAMA 2021, 326, 1703–1712. [Google Scholar] [CrossRef] [PubMed]
- RECOVERY Collaborative Group. Aspirin in Patients Admitted to Hospital with COVID-19 (RECOVERY): A Randomised, Controlled, Open-Label, Platform Trial. Lancet 2021, 399, 143–151. [Google Scholar] [CrossRef]
- Sayed Ahmed, H.A.; Merrell, E.; Ismail, M.; Joudeh, A.I.; Riley, J.B.; Shawkat, A.; Habeb, H.; Darling, E.; Goweda, R.A.; Shehata, M.H.; et al. Rationales and Uncertainties for Aspirin Use in COVID-19: A Narrative Review. Fam. Med. Community Health 2021, 9, e000741. [Google Scholar] [CrossRef]
- Rivas-Estilla, A.M.; Bryan-Marrugo, O.L.; Trujillo-Murillo, K.; Pérez-Ibave, D.; Charles-Niño, C.; Pedroza-Roldan, C.; Ríos-Ibarra, C.; Ramírez-Valles, E.; Ortiz-López, R.; Islas-Carbajal, M.C.; et al. Cu/Zn Superoxide Dismutase (SOD1) Induction Is Implicated in the Antioxidative and Antiviral Activity of Acetylsalicylic Acid in HCV-Expressing Cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 302, G1264–G1273. [Google Scholar] [CrossRef]
- Yin, P.; Zhang, L. Aspirin Inhibits Hepatitis C Virus Entry by Downregulating Claudin-1. J. Viral Hepat. 2016, 23, 62–64. [Google Scholar] [CrossRef]
- Liao, Y.-H.; Hsu, R.-J.; Wang, T.-H.; Wu, C.-T.; Huang, S.-Y.; Hsu, C.-Y.; Su, Y.-C.; Hsu, W.-L.; Liu, D.-W. Aspirin Decreases Hepatocellular Carcinoma Risk in Hepatitis C Virus Carriers: A Nationwide Cohort Study. BMC Gastroenterol. 2020, 20, 6. [Google Scholar] [CrossRef]
- Li, X.; Wu, S.; Yu, Y. Aspirin Use and the Incidence of Hepatocellular Carcinoma in Patients with Hepatitis B Virus or Hepatitis C Virus Infection: A Meta-Analysis of Cohort Studies. Front. Med. 2021, 7, 968. [Google Scholar] [CrossRef]
- Paik, Y.-H.; Kim, J.K.; Lee, J.I.; Kang, S.H.; Kim, D.Y.; An, S.H.; Lee, S.J.; Lee, D.K.; Han, K.-H.; Chon, C.Y.; et al. Celecoxib Induces Hepatic Stellate Cell Apoptosis through Inhibition of Akt Activation and Suppresses Hepatic Fibrosis in Rats. Gut 2009, 58, 1517–1527. [Google Scholar] [CrossRef]
- Leng, J.; Han, C.; Demetris, A.J.; Michalopoulos, G.K.; Wu, T. Cyclooxygenase-2 Promotes Hepatocellular Carcinoma Cell Growth through Akt Activation: Evidence for Akt Inhibition in Celecoxib-Induced Apoptosis. Hepatology 2003, 38, 756–768. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, H.; Kondo, M.; Nakamori, S.; Nagano, H.; Wakasa, K.-I.; Sugita, Y.; Chang-De, J.; Kobayashi, S.; Damdinsuren, B.; Dono, K.; et al. JTE-522, a Cyclooxygenase-2 Inhibitor, Is an Effective Chemopreventive Agent against Rat Experimental Liver fibrosis1. Gastroenterology 2003, 125, 556–571. [Google Scholar] [CrossRef]
- Cheng, J.; Imanishi, H.; Amuro, Y.; Hada, T. NS-398, a Selective Cyclooxygenase 2 Inhibitor, Inhibited Cell Growth and Induced Cell Cycle Arrest in Human Hepatocellular Carcinoma Cell Lines. Int. J. Cancer 2002, 99, 755–761. [Google Scholar] [CrossRef] [PubMed]
- Malehmir, M.; Pfister, D.; Gallage, S.; Szydlowska, M.; Inverso, D.; Kotsiliti, E.; Leone, V.; Peiseler, M.; Surewaard, B.G.J.; Rath, D.; et al. Platelet GPIbα Is a Mediator and Potential Interventional Target for NASH and Subsequent Liver Cancer. Nat. Med. 2019, 25, 641–655. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Chung, G.E.; Lee, J.-H.; Oh, S.; Nam, J.Y.; Chang, Y.; Cho, H.; Ahn, H.; Cho, Y.Y.; Yoo, J.-J.; et al. Antiplatelet Therapy and the Risk of Hepatocellular Carcinoma in Chronic Hepatitis B Patients on Antiviral Treatment. Hepatology 2017, 66, 1556–1569. [Google Scholar] [CrossRef] [PubMed]
- Alexander, S. The Prevention of Recurrent Herpes Simplex with Aspirin. Br. J. Dermatol. 1962, 74, 83–85. [Google Scholar] [CrossRef]
- Speir, E.; Yu, Z.X.; Ferrans, V.J.; Huang, E.S.; Epstein, S.E. Aspirin Attenuates Cytomegalovirus Infectivity and Gene Expression Mediated by Cyclooxygenase-2 in Coronary Artery Smooth Muscle Cells. Circ. Res. 1998, 83, 210–216. [Google Scholar] [CrossRef]
- Zhu, H.; Cong, J.-P.; Yu, D.; Bresnahan, W.A.; Shenk, T.E. Inhibition of Cyclooxygenase 2 Blocks Human Cytomegalovirus Replication. Proc. Natl. Acad. Sci. USA 2002, 99, 3932–3937. [Google Scholar] [CrossRef]
- De Benedittis, G.; Besana, F.; Lorenzetti, A. A New Topical Treatment for Acute Herpetic Neuralgia and Post-Herpetic Neuralgia: The Aspirin/diethyl Ether Mixture. An Open-Label Study plus a Double-Blind Controlled Clinical Trial. Pain 1992, 48, 383–390. [Google Scholar] [CrossRef]
- Bareggi, S.R.; Pirola, R.; De Benedittis, G. Skin and Plasma Levels of Acetylsalicylic Acid: A Comparison between Topical Aspirin/diethyl Ether Mixture and Oral Aspirin in Acute Herpes Zoster and Postherpetic Neuralgia. Eur. J. Clin. Pharmacol. 1998, 54, 231–235. [Google Scholar] [CrossRef]
- Primache, V.; Binda, S.; De Benedittis, G.; Barbi, M. In Vitro Activity of Acetylsalicylic Acid on Replication of Varicella-Zoster Virus. New Microbiol. 1998, 21, 397–401. [Google Scholar] [PubMed]
- Karádi, I.; Kárpáti, S.; Romics, L. Aspirin in the Management of Recurrent Herpes Simplex Virus Infection. Ann. Intern. Med. 1998, 128, 696–697. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.-F.; Wang, H.; Li, Z.-J.; Deng, X.-Y.; Xiang, H.; Tao, Y.-G.; Li, W.; Tang, M.; Cao, Y. Aspirin Induces Lytic Cytotoxicity in Epstein-Barr Virus-Positive Cells. Eur. J. Pharmacol. 2008, 589, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Bitko, V.; Velazquez, A.; Yang, L.; Yang, Y.C.; Barik, S. Transcriptional Induction of Multiple Cytokines by Human Respiratory Syncytial Virus Requires Activation of NF-Kappa B and Is Inhibited by Sodium Salicylate and Aspirin. Virology 1997, 232, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Graham, N.M.; Burrell, C.J.; Douglas, R.M.; Debelle, P.; Davies, L. Adverse Effects of Aspirin, Acetaminophen, and Ibuprofen on Immune Function, Viral Shedding, and Clinical Status in Rhinovirus-Infected Volunteers. J. Infect. Dis. 1990, 162, 1277–1282. [Google Scholar] [CrossRef]
- Hsia, J.; Simon, G.L.; Higgins, N.; Goldstein, A.L.; Hayden, F.G. Immune Modulation by Aspirin during Experimental Rhinovirus Colds. Bull. N. Y. Acad. Med. 1989, 65, 45–56. [Google Scholar]
- Huang, R.T.; Dietsch, E. Anti-Influenza Viral Activity of Aspirin in Cell Culture. N. Engl. J. Med. 1988, 319, 797. [Google Scholar]
- Beutler, A.I.; Chesnut, G.T.; Mattingly, J.C.; Jamieson, B. FPIN’s Clinical Inquiries. Aspirin Use in Children for Fever or Viral Syndromes. Am. Fam. Phys. 2009, 80, 1472–1474. [Google Scholar]
- Floyd, N.; Ferro, A. Mechanisms of aspirin resistance. Pharmacol. Ther. 2014, 141, 69–78. [Google Scholar] [CrossRef]


{kind=link}
| ASA’s Target | Main Results of ASA Exposure | |
|---|---|---|
| Platelets |
| |
| Biofilm |
| |
| Bacteria | General considerations |
|
| Endotoxin |
| |
| BSI/sepsis |
| |
| Endocarditis |
| |
| Gastroenteritis and liver abscess |
| |
| PJI |
| |
| Pneumonia |
| |
| Mycobacterial diseases |
| |
| Fungi |
| |
| Parasites |
| |
| Viruses | HIV |
|
| SARS-CoV-2 |
| |
| Viral hepatitis |
| |
| Others |
| |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Bella, S.; Luzzati, R.; Principe, L.; Zerbato, V.; Meroni, E.; Giuffrè, M.; Crocè, L.S.; Merlo, M.; Perotto, M.; Dolso, E.; et al. Aspirin and Infection: A Narrative Review. Biomedicines 2022, 10, 263. https://doi.org/10.3390/biomedicines10020263
Di Bella S, Luzzati R, Principe L, Zerbato V, Meroni E, Giuffrè M, Crocè LS, Merlo M, Perotto M, Dolso E, et al. Aspirin and Infection: A Narrative Review. Biomedicines. 2022; 10(2):263. https://doi.org/10.3390/biomedicines10020263
Chicago/Turabian StyleDi Bella, Stefano, Roberto Luzzati, Luigi Principe, Verena Zerbato, Elisa Meroni, Mauro Giuffrè, Lory Saveria Crocè, Marco Merlo, Maria Perotto, Elisabetta Dolso, and et al. 2022. "Aspirin and Infection: A Narrative Review" Biomedicines 10, no. 2: 263. https://doi.org/10.3390/biomedicines10020263
APA StyleDi Bella, S., Luzzati, R., Principe, L., Zerbato, V., Meroni, E., Giuffrè, M., Crocè, L. S., Merlo, M., Perotto, M., Dolso, E., Maurel, C., Lovecchio, A., Dal Bo, E., Lagatolla, C., Marini, B., Ippodrino, R., & Sanson, G. (2022). Aspirin and Infection: A Narrative Review. Biomedicines, 10(2), 263. https://doi.org/10.3390/biomedicines10020263