
The Role of Melatonin in Pregnancy and the Health Benefits for the Newborn
Abstract
1. Introduction
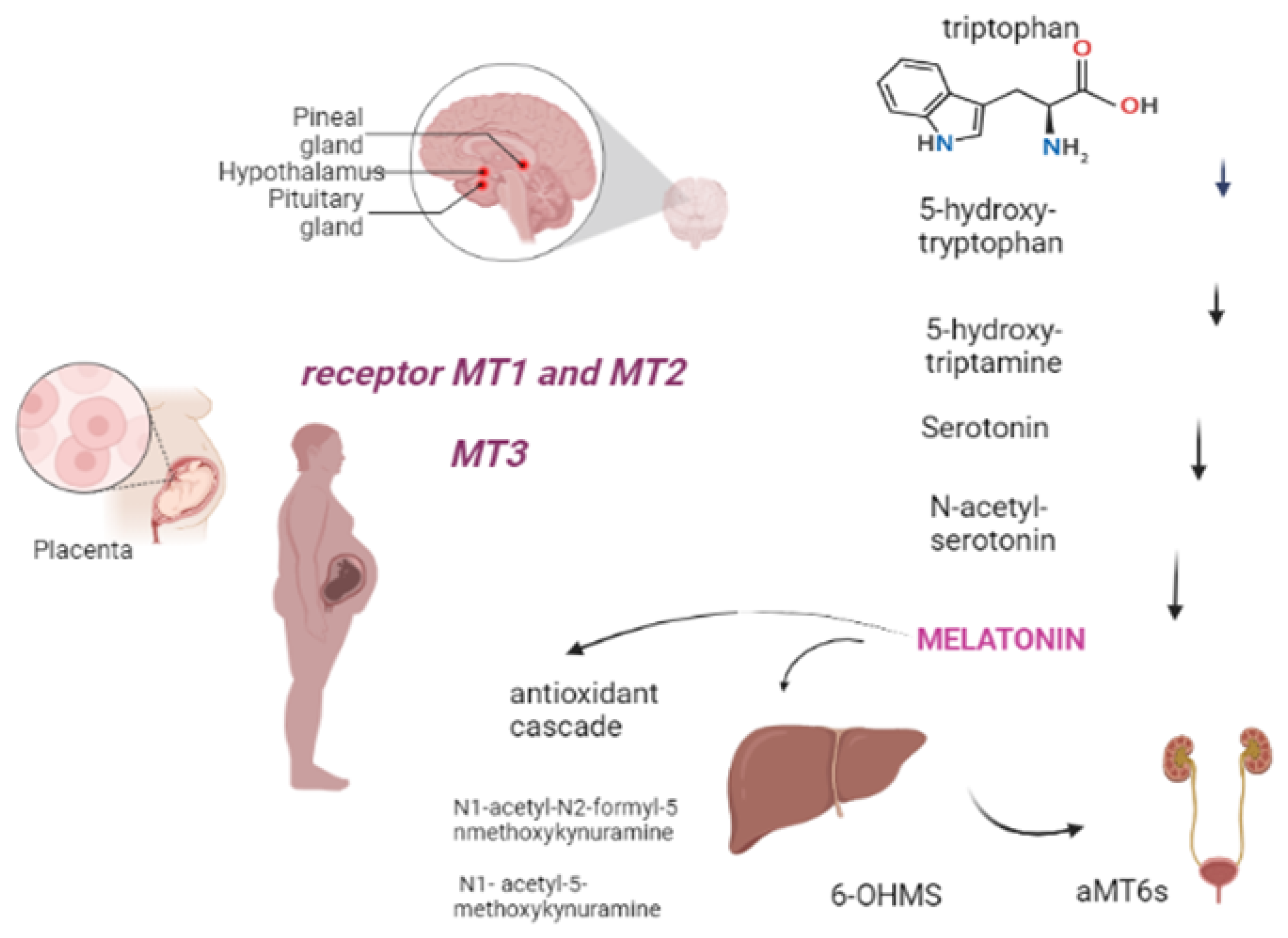
2. Metabolism
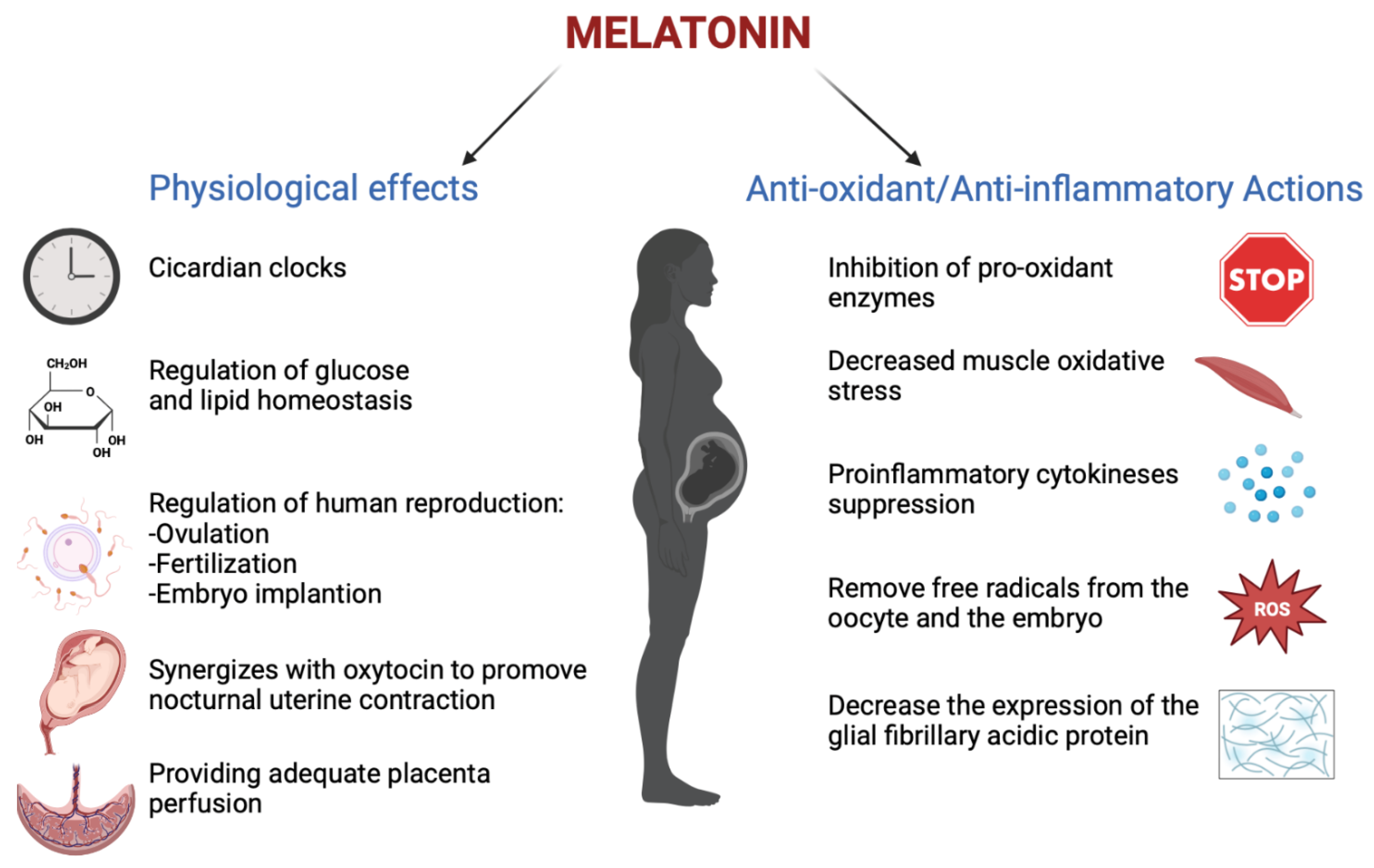
3. Melatonin in Pregnancy
4. Melatonin and Placenta
5. Melatonin in High-Risk Pregnancy
5.1. Melatonin and Gestational Diabetes
5.2. Melatonin and Preeclampsia
5.3. Melatonin and Intrauterine Growth Restriction (IUGR)
6. Melatonin and Labor
7. Melatonin and Preterm Birth
8. Melatonin in Breastfeeding
9. Melatonin in Newborn
10. Hypoxic Ischemic Encephalopathy
11. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Carlomagno, G.; Minini, M.; Tilotta, M.; Unfer, V. From Implantation to Birth: Insight into Molecular Melatonin Functions. Int. J. Mol. Sci. 2018, 19, 2802. [Google Scholar] [CrossRef]
- Xie, F.; Zhang, J.; Zhai, M.; Liu, Y.; Hu, H.; Yu, Z.; Zhang, J.; Lin, S.; Liang, D.; Cao, Y. Melatonin ameliorates ovarian dysfunction by regulating autophagy in PCOS via the PI3K-Akt pathway. Reproduction 2021, 162, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Tamura, H.; Takasaki, A.; Taketani, T.; Tanabe, M.; Kizuka, F.; Lee, L.; Tamura, I.; Maekawa, R.; Asada, H.; Yamagata, Y.; et al. Melatonin as a free radical scavenger in the ovarian follicle. Endocr. J. 2013, 60, 1–13. [Google Scholar] [CrossRef]
- Zare, H.; Shafabakhsh, R.; Reiter, R.J.; Asemi, Z. Melatonin is a potential inhibitor of ovarian cancer: Molecular aspects. J. Ovarian Res. 2019, 12, 26. [Google Scholar] [CrossRef]
- Garcia, J.J.; Lopez-Pingarron, L.; Almeida-Souza, P.; Tres, A.; Escudero, P.; Garcia-Gil, F.A.; Tan, D.X.; Reiter, R.J.; Ramirez, J.M.; Bernal-Perez, M. Protective effects of melatonin in reducing oxidative stress and in preserving the fluidity of biological membranes: A review. J. Pineal Res. 2014, 56, 225–237. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, S.; Nakazawa, K.; Sakai, J.; Kometani, K.; Iwashita, M.; Yoshimura, Y.; Maruyama, T. Melatonin as a local regulator of human placental function. J. Pineal Res. 2005, 39, 261–265. [Google Scholar] [CrossRef]
- Morrissey, M.J.; Duntley, S.P.; Anch, A.M.; Nonneman, R. Active sleep and its role in the prevention of apoptosis in the developing brain. Med. Hypotheses 2004, 62, 876–879. [Google Scholar] [CrossRef] [PubMed]
- Sharkey, J.T.; Puttaramu, R.; Word, R.A.; Olcese, J. Melatonin synergizesn with oxytocin to enhance contractility of human myometrial smoothn muscle cells. J. Clin. Endocrinol. Metab. 2009, 94, 421–427. [Google Scholar] [CrossRef] [PubMed]
- Swarnamani, K.; Davies-Tuck, M.; Wallace, E.; Mol, B.W.; Mockler, J. A double-blind randomised placebo-controlled trial of melatonin as an adjuvant agent in induction of labour (MILO): A study protocol. BMJ Open 2020, 10, e032480. [Google Scholar] [CrossRef]
- Slominski, R.M.; Reiter, R.J.; Schlabritz-Loutsevitch, N.; Ostrom, R.S.; Slominski, A.T. Melatonin membrane receptors in peripheral tissues: Distribution and functions. Mol. Cell. Endocrinol. 2012, 351, 152–166. [Google Scholar] [CrossRef]
- Back, K.; Tan, D.X.; Reiter, R.J. Melatonin biosynthesis in plants: Multiple pathways catalyze tryptophan to melatonin in the cytoplasm or chloroplasts. J. Pineal Res. 2016, 61, 426–437. [Google Scholar] [CrossRef]
- Hardeland, R.; Pandi-Perumal, S.R.; Cardinali, D.P. Melatonin. Int. J. Biochem. Cell Biol. 2006, 38, 313–316. [Google Scholar] [CrossRef]
- Ekmekcioglu, C. Melatonin receptors in humans: Biological role and clinical relevance. Biomed. Pharmacother. 2006, 60, 97–108. [Google Scholar] [CrossRef]
- Hardeland, R.; Tan, D.X.; Reiter, R.J. Kynuramines, metabolites of melatonin and other indoles: The resurrection of an almost forgotten class of biogenic amines. J. Pineal Res. 2009, 47, 109–126. [Google Scholar] [CrossRef]
- Chuffa, L.G.; Lupi Júnior, L.A.; Seiva, F.R.; Martinez, M.; Domeniconi, R.F.; Pinheiro, P.F.; Dos Santos, L.D.; Martinez, F.E. Quantitative proteomic profiling reveals that diverse metabolic pathways are influenced by melatonin in an in vivo model of ovarian carcinoma. J. Proteome Res. 2016, 15, 3872–3882. [Google Scholar] [CrossRef]
- Dubocovich, M.L.; Markowska, M. Functional MT1 and MT2 melatonin receptors in mammals. Endocrine 2005, 27, 101–110. [Google Scholar] [CrossRef]
- Kim, T.K.; Kleszczynski, K.; Janjetovic, Z.; Sweatman, T.; Lin, Z.; Li, W.; Reiter, R.J.; Fischer, T.W.; Slominski, A.T. Metabolism of melatonin and biological activity of intermediates of melatoninergic pathway in human skin cells. FASEB J. 2013, 27, 2742–2755. [Google Scholar] [CrossRef]
- Slominski, A.T.; Zmijewski, M.A.; Semak, I.; Kim, T.K.; Janjetovic, Z.; Slominski, R.M.; Zmijewski, J.W. Melatonin, mitochondria, and the skin. Cell Mol. Life Sci. 2017, 74, 3913–3925. [Google Scholar] [CrossRef] [PubMed]
- Man, G.C.W.; Zhang, T.; Chen, X.; Wang, J.; Wu, F.; Liu, Y.; Wang, C.C.; Cheong, Y.; Li, T.C. The regulations and role of circadian clock and melatonin in uterine receptivity and pregnancy-An immunological perspective. Am. J. Reprod. Immunol. 2017, 78, e12715. [Google Scholar] [CrossRef] [PubMed]
- Kivela, A. Serum melatonin during human pregnancy. Acta Endocrinol. 1991, 124, 233–237. [Google Scholar] [CrossRef]
- Claustrat, B.; Brun, J.; Chazot, G. The basic physiology and pathophysiology of melatonin. Sleep Med. Rev. 2005, 9, 11–24. [Google Scholar] [CrossRef]
- Silver, A.C.; Arjona, A.; Walker, W.E.; Fikrig, E. The circadian clock controls toll-like-receptor 9-mediated innate and adaptive immunity. Immunity 2012, 36, 251–261. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.; Tamura, H.; Kashida, S.; Takayama, H.; Yagamata, Y.; Karube, A.; Sugino, N.; Kato, H. Changes of serum melatonin level and its relationship to the feto-placental unit during pregnancy. J. Pineal Res. 2001, 30, 29–33. [Google Scholar] [CrossRef] [PubMed]
- NIH 2017. What Is a High-Risk Pregnancy? Available online: https://www.nichd.nih.gov/health/topics/pregnancy/conditioninfo/high-risk (accessed on 31 January 2017).
- Lanoix, D.; Guérin, P.; Vaillancourt, C. Placental melatonin production and melatonin receptor expression are altered in preeclampsia: New insights into the role of this hormone in pregnancy. J. Pineal Res. 2012, 53, 417–425. [Google Scholar] [CrossRef]
- Ejaz, H.; Figaro, J.K.; Woolner, A.M.F.; Thottakam, B.M.V.; Galley, H.F. Maternal Serum Melatonin Increases During Pregnancy and Falls Immediately After Delivery Implicating the Placenta as a Major Source of Melatonin. Front. Endocrinol. 2021, 11, 623038. [Google Scholar] [CrossRef]
- Reiter, R.J.; Tan, D.X.; Korkmaz, A.; Rosales-Corral, S.A. Melatonin and stable circadian rhythms optimize maternal, placental and fetal physiology. Hum. Reprod. Update 2014, 20, 293–307. [Google Scholar] [CrossRef] [PubMed]
- Spaight, C.; Gross, J.; Horsch, A.; Puder, J.J. Gestational Diabetes Mellitus. Endocr. Dev. 2016, 31, 163–178. [Google Scholar] [CrossRef]
- Liu, S.; Guo, Y.; Yuan, Q.; Pan, Y.; Wang, L.; Liu, Q.; Wang, F.; Wang, J.; Hao, A. Melatonin prevents neural tube defects in the offspring of diabetic pregnancy. J. Pineal Res. 2015, 59, 508–517. [Google Scholar] [CrossRef]
- Firneisz, G.; Rosta, K.; Al-Aissa, Z.; Hadarits, O.; Harreiter, J.; Nádasdi, Á.; Bancher-Todesca, D.; Németh, L.; Igaz, P.; Rigó, J., Jr.; et al. The MTNR1B rs10830963 Variant in Interaction with Pre-Pregnancy BMI is a Pharmacogenetic Marker for the Initiation of Antenatal Insulin Therapy in Gestational Diabetes Mellitus. Int. J. Mol. Sci. 2018, 19, 3734. [Google Scholar] [CrossRef] [PubMed]
- Nisa, H.; Qi, K.H.T.; Leng, J.; Zhou, T.; Liu, H.; Li, W.; Wang, L.; Li, N.; Hu, G.; Qi, L. The Circadian Rhythm-Related MTNR1B Genotype, Gestational Weight Gain, and Postpartum Glycemic Changes. J. Clin. Endocrinol. Metab. 2018, 103, 2284–2290. [Google Scholar] [CrossRef]
- Zhang, Y.; Sun, C.M.; Hu, X.Q.; Zhao, Y. Relationship between melatonin receptor 1B and insulin receptor substrate 1 polymorphisms with gestational diabetes mellitus: A systematic review and meta-analysis. Sci. Rep. 2014, 4, 6113. [Google Scholar] [CrossRef]
- Huang, B.; Wang, Y.; Qin, L.; Wei, Q.; Liu, N.; Jiang, M.; Yu, H.-P.; Yu, X.-Y. A functional polymorphism rs10830963 in melatonin receptor 1B associated with the risk of gestational. Biosci. Rep. 2019, 39, BSR20190744. [Google Scholar] [CrossRef]
- Liao, S.; Liu, Y.; Tan, Y.; Gan, L.; Mei, J.; Song, W.; Chi, S.; Dong, X.; Chen, X.; Deng, S. Association of genetic variants of melatonin receptor 1B with gestational plasma glucose level and risk of glucose intolerance in pregnant Chinese women. PLoS ONE 2012, 7, e40113. [Google Scholar] [CrossRef]
- Liu, C.; Wu, Y.; Li, H.; Qi, Q.; Langenberg, C.; Loos, R.J.; Lin, X. MTNR1B rs10830963 is associated with fasting plasma glucose, HbA1C and impaired beta-cell function in Chinese Hans from Shanghai. BMC Med. Genet. 2010, 11, 59. [Google Scholar] [CrossRef]
- Laste, G.; da Silva, A.A.; Gheno, B.R.; et Rychcik, P.M. Relationship between melatonin and high-risk pregnancy: A review of investigations published between the years 2010 and 2020. Chronobiol. Int. 2021, 38, 168–181. [Google Scholar] [CrossRef]
- Mol, B.W.J.; Roberts, C.T.; Thangaratinam, S.; Magee, L.A.; de Groot, C.J.M.; Hofmeyr, G.J. Pre-eclampsia. Lancet 2016, 387, 999–1011. [Google Scholar] [CrossRef]
- Ma’ayeh, M.; Costantine, M.M. Prevention of preeclampsia. Semin. Fetal Neonatal Med. 2020, 25, 101123. [Google Scholar] [CrossRef]
- Phipps, E.; Prasanna, D.; Brima, W.; Jim, B. Preeclampsia: Updates in Pathogenesis, Definitions, and Guidelines. Clin. J. Am. Soc. Nephrol. 2016, 11, 1102–1113. [Google Scholar] [CrossRef] [PubMed]
- Sagrillo-Fagundes, L.; Assunção Salustiano, E.M.; Ruano, R.; Markus, R.P.; Vaillancourt, C. Melatonin modulates autophagy and inflammation protecting human placental trophoblast from hypoxia/reoxygenation. J. Pineal Res. 2018, 65, e12520. [Google Scholar] [CrossRef] [PubMed]
- Cheung, R.T.F. The utility of melatonin in reducing cerebral damage resulting from ischemia and reperfusion. J. Pineal Res. 2003, 34, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Genario, R.; Morello, E.; Bueno, A.A.; Santos, H.O. The usefulness of melatonin in the field of obstetrics and gynecology. Pharmacol. Res. 2019, 147, 104337. [Google Scholar] [CrossRef] [PubMed]
- McKinney, D.; Boyd, H.; Langager, A.; Oswald, M.; Pfister, A.; Warshak, C.R. The impact of fetal growth restriction on latency in the setting of expectant management of preeclampsia. Am. J. Obstet. Gynecol. 2016, 214, 395.e1-7. [Google Scholar] [CrossRef]
- Berbets, A.M.; Barbe, A.M.; Andriiets, O.A.; Andriiets, A.V.; Yuzko, O.M. Melatonin Levels Decrease in the Umbilical Cord in Case of Intrauterine Growth Restriction. J. Med. Life 2020, 13, 548–553. [Google Scholar] [CrossRef] [PubMed]
- Grobman, W.A.; Rice, M.M.; Reddy, U.M.; Tita, A.T.N.; Silver, R.M.; Mallett, G.; Hill, K.; Thom, E.A.; El-Sayed, Y.Y.; Perez-Delboy, A.; et al. Labor induction versus expectant management in low-risk nulliparous women. N. Engl. J. Med. 2018, 379, 513–523. [Google Scholar] [CrossRef]
- Walker, K.F.; Bugg, G.J.; Macpherson, M.; McCormick, C.; Grace, N.; Wildsmith, C.; Bradshaw, L.; Smith, G.C.S.; Thornton, J.G. Randomized trial of labor induction in women 35 years of age or older. N. Engl. J. Med. 2016, 374, 813–822. [Google Scholar] [CrossRef]
- Hammad, I.A.; Chauhan, S.P.; Magann, E.F.; Abuhamad, A.Z. Peripartum complications with cesarean delivery: A review of Maternal-Fetal medicine units network publications. J. Matern. Fetal Neonatal Med. 2014, 27, 463–474. [Google Scholar] [CrossRef]
- Clark, E.A.S.; Silver, R.M. Long-term maternal morbidity associated with repeat cesarean delivery. Am. J. Obstet. Gynecol. 2011, 205, S2–S10. [Google Scholar] [CrossRef] [PubMed]
- Ramachandrappa, A.; Jain, L. Elective cesarean section: Its impact on neonatal respiratory outcome. Clin. Perinatol. 2008, 35, 373–393. [Google Scholar] [CrossRef]
- Sagrillo-Fagundes, L.; Soliman, A.; Vaillancourt, C. Maternal and placental melatonin: Actions and implication for successful pregnancies. Minerva Ginecol 2014, 66, 251–266. [Google Scholar]
- Olcese, J.; Lozier, S.; Paradise, C. Melatonin and the circadian timing of human parturition. Reprod. Sci. 2013, 20, 168–174. [Google Scholar] [CrossRef] [PubMed]
- Rahman, S.A.; Bibbo, C.; Olcese, J.; Czeisler, C.A.; Robinson, J.N.; Klerman, E.B. Relationship between endogenous melatonin concentrations and uterine contractions in late third trimester of human pregnancy. J. Pineal Res. 2019, 66, e12566. [Google Scholar] [CrossRef] [PubMed]
- Olcese, J.; Beesley, S. Clinical significance of melatonin receptors in the human myometrium. Fertil. Steril. 2014, 102, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.H.; Park, J.H.; Lee, Y.L.; Kang, I.J.; Kim, D.W.; Hwang, I.K.; Lee, C.-H.; Yan, B.C.; Kim, Y.-M.; Lee, T.-K.; et al. Melatonin improves vascular cognitive impairment induced by ischemic stroke by remyelination via activation of ERK1/2 signaling and restoration of glutamatergic synapses in the gerbil hippocampus. Biomed. Pharmacother. 2018, 108, 687–697. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Wang, Z.; Pan, S.; Zhang, H.; Fang, M.; Jiang, H.; Zhang, H.; Gao, Z.; Xu, K.; Li, Z.; et al. Melatonin protects against blood-brain barrier damage by inhibiting the TLR4/ NF-κB signaling pathway after LPS treatment in neonatal rats. Oncotarget 2017, 8, 31638–31654. Available online: https://pubmed.ncbi.nlm.nih.gov/28404943 (accessed on 1 October 2022). [CrossRef]
- Villapol, S.; Fau, S.; Renolleau, S.; Biran, V.; Charriaut-Marlangue, C.; Baud, O. Melatonin promotes myelination by decreasing white matter inflammation after neonatal stroke. Pediatr. Res. 2011, 69, 51–55. Available online: https://pubmed.ncbi.nlm.nih.gov/20856166 (accessed on 1 October 2022). [CrossRef]
- Sánchez-Hidalgo, M.; Guerrero, J.M.; Villegas, I.; Packham, G.; de la Lastra, C.A. Melatonin, a natural programmed cell death inducer in cancer. Curr. Med. Chem. 2012, 19, 3805–3821. Available online: https://pubmed.ncbi.nlm.nih.gov/22612707 (accessed on 1 October 2022). [CrossRef]
- Biran, V.; Decobert, F.; Bednarek, N.; Boizeau, P.; Benoist, J.F.; Claustrat, B.; Barré, J.; Colella, M.; Frérot, A.; Garnotel, R.; et al. Melatonin Levels in Preterm and Term Infants and Their Mothers. Int. J. Mol. Sci. 2019, 20, 2077. [Google Scholar] [CrossRef]
- Netzer-Tomkins, H.; Rubin, L.; Ephros, M. Breastfeeding is associ- ated with decreased hospitalization for neonatal fever. Breasfeed. Med. 2016, 11, 218–221. [Google Scholar] [CrossRef]
- Zalewski, B.M.; Patro, B.; Veldhorst, M.; Kouwenhoven, S.; Crespo Escobar, P.; Calvo Lerma, J.; Koletzko, B.; van Goudoever, J.B.; Sza-jewska, H. Nutrition of infants and young children (one to three years) and its effect on later health: A systematic review of cur- rent recommendations (EarlyNutrition project). Crit. Rev. Food Sci. Nutr. 2017, 57, 489–500. [Google Scholar] [CrossRef] [PubMed]
- Anderson, G.; Vaillancourt, C.; Maes, M.; Reiter, R.R. Breastfeeding and melatonin: Implications for improving perinatal health. J. Breastfeed. Biol. 2016, 1, 8–20. [Google Scholar] [CrossRef]
- Dvorak, B. Milk epidermal growth factor and gut protection. J. Pediatr. 2010, 156 (Suppl. 2), S31–S35. [Google Scholar] [CrossRef] [PubMed]
- Sumová, A.; Bendová, Z.; Sládek, M.; El-Hennamy, R.; Matejů, K.; Polidarová, L.; Sosniyenko, S.; Illnerová, H. Circadian molecular clocks tick along ontogenesis. Physiol. Res. 2008, 57 (Suppl. 3), S139–S148. [Google Scholar] [CrossRef] [PubMed]
- Katzer, D.; Pauli, L.; Mueller, A.; Reutter, H.; Reinsberg, J.; Fimmers, R.; Bartmann, P.; Bagci, S. Melatonin concentrations and antioxidative capacity of human breast milk according to gestational age and the time of day. J. Hum. Lact. 2016, 32, NP105–10. [Google Scholar] [CrossRef] [PubMed]
- Dumbell, R.; Matveeva, O.; Oster, H. Circadian clocks, stress, and immunity. Front. Endocrinol. 2016, 7, 37. [Google Scholar] [CrossRef]
- Matamoros, S.; Gras-Leguen, C.; Le Vacon, F.; Potel, G.; de La Coche-tiere, M.F. Development of intestinal microbiota in infants and its impact on health. Trends Microbiol. 2013, 21, 167–173. [Google Scholar] [CrossRef]
- Thibeau, S.; D’Apolito, K.; Minnick, A.F.; Dietrich, M.S.; Kane, B.; Cooley, S.; Groer, M. Relationships of maternal stress with milk immune components in African American mothers of healthy term infants. Breastfeed. Med. 2016, 11, 6–14. [Google Scholar] [CrossRef]
- Cohen Engler, A.; Hadash, A.; Shehadeh, N.; Pillar, G. Breastfeeding may improve nocturnal sleep and reduce infantile colic: Potential role of breast milk melatonin. Eur. J. Pediatr. 2012, 171, 729–732. [Google Scholar] [CrossRef]
- Rochow, N.; Fusch, G.; Choi, A.; Chessell, L.; Elliott, L.; McDonald, K.; Kuiper, E.; Purcha, M.; Turner, S.; Chan, E.; et al. Target fortification of breast milk with fat, protein, and carbohydrates for preterm infants. J. Pediatr. 2013, 163, 1001–1007. [Google Scholar] [CrossRef]
- Chen, Y.C.; Tain, Y.L.; Sheen, J.M.; Huang, L.T. Melatonin utility in neonates and children. J. Formos. Med. Assoc. 2012, 111, 57–66. [Google Scholar] [CrossRef]
- Saugstad, O.D. Oxygen and retinopathy of prematurity. J. Perinatol. 2006, 26 (Suppl. 1), S46–S50; discussion S63–S64. [Google Scholar] [CrossRef] [PubMed]
- McLean, C.; Ferriero, D. Mechanisms of hypoxic-ischemic injury in the term infant. Semin. Perinatol. 2004, 28, 425–432. [Google Scholar] [CrossRef]
- Lai, M.C.; Yang, S.N. Perinatal hypoxic-ischemic encephalopathy. J. Biomed. Biotechnol. 2011, 2011, 609813. [Google Scholar] [CrossRef]
- Rasmussen, D.D.; Marck, B.T.; Boldt, B.M.; Yellon, S.M.; Matsumoto, A.M. Suppression of hypothalamic pro-opiomelanocortin (POMC) gene expression by daily melatonin supplementation in aging rats. J. Pineal Res. 2003, 34, 127–133. [Google Scholar] [CrossRef]
- Muñoz-Hoyos, A.; Bonillo-Perales, A.; Avila-Villegas, R.; González-Ripoll, M.; Uberos, J.; Florido-Navío, J.; Molina-Carballo, A. Melatonin levels during the first week of life and their relation with the antioxidant response in the perinatal period. Neonatology 2007, 92, 209–216. [Google Scholar] [CrossRef]
- Saito, S.; Sakai, M. Th1/Th2 balance in preeclampsia. J. Reprod. Immunol. 2003, 59, 161–173. [Google Scholar] [CrossRef]
- Stoll, B.J.; Hansen, N.I.; Bell, E.F.; Shankaran, S.; Laptook, A.R.; Walsh, M.C.; Hale, E.C.; Newman, N.S.; Schibler, K.; Carlo, W.A.; et al. Eunice Kennedy Shriver National Institute of Child Health and Human Development Neonatal Research Network. Neonatal outcomes of extremely preterm infants from the NICHD Neonatal Research Network. Pediatrics 2010, 126, 443–456. [Google Scholar] [CrossRef] [PubMed]
- Volpe, J.J. Encephalopathy of prematurity includes neuronal abnormalities. Pediatrics 2005, 116, 221–225, Erratum in Pediatrics 2006, 118, 1807. [Google Scholar] [CrossRef] [PubMed]
- Back, S.A.; Riddle, A.; McClure, M.M. Maturation-dependent vulnerability of perinatal white matter in premature birth. Stroke 2007, 38 (Suppl. 2), 724–730. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, F.F.; Ferriero, D.M. Therapeutics for neonatal brain injury. Pharmacol. Ther. 2008, 120, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.P.; Ramaswamy, V.; Michelson, D.; Barkovich, A.J.; Holshouser, B.; Wycliffe, N.; Glidden, D.V.; Deming, D.; Partridge, J.C.; Wu, Y.W.; et al. Patterns of brain injury in term neonatal encephalopathy. J. Pediatr. 2005, 146, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Bennet, L.; Roelfsema, V.; Pathipati, P.; Quaedackers, J.S.; Gunn, A.J. Relationship between evolving epileptiform activity and delayed loss of mitochondrial activity after asphyxia measured by near-infrared spectroscopy in preterm fetal sheep. J. Physiol. 2006, 572 Pt 1, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Vannucci, S.J.; Hagberg, H. Hypoxia-ischemia in the immature brain. J. Exp. Biol. 2004, 207 Pt 18, 3149–3154. [Google Scholar] [CrossRef]
- Bennet, L.; Tan, S.; Van den Heuij, L.; Derrick, M.; Groenendaal, F.; van Bel, F.; Juul, S.; Back, S.A.; Northington, F.; Robertson, N.J.; et al. Cell therapy for neonatal hypoxia-ischemia and cerebral palsy. Ann. Neurol. 2012, 71, 589–600. [Google Scholar] [CrossRef]
- Merchant, N.M.; Azzopardi, D.V.; Hawwa, A.F.; McElnay, J.C.; Middleton, B.; Arendt, J.; Arichi, T.; Gressens, P.; Edwards, A.D. Pharmacokinetics of melatonin in preterm infants. Br. J. Clin. Pharmacol. 2013, 76, 725–733. Available online: https://pubmed.ncbi.nlm.nih.gov/23432339 (accessed on 1 October 2022). [CrossRef]
- Praninskienė, R.; Dumalakienė, I.; Kemežys, R.; Mauricas, M.; Jučaitė, A. Diurnal melatonin patterns in children: Ready to apply in clinical practice? Pediatr. Neurol. 2012, 46, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Gitto, E.; Marseglia, L.; Manti, S.; D’Angelo, G.; Barberi, I.; Salpietro, C.; Reiter, R.J. Protective role of melatonin in neonatal diseases. Oxid. Med. Cell. Longev. 2013, 2013, 980374. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.W.; Chen, T.Y.; Hung, C.Y.; Tai, S.H.; Huang, S.Y.; Chang, C.C.; Hung, H.Y.; Lee, E.J. Melatonin protects brain against ischemia/reperfusion injury by attenuating endoplasmic reticulum stress. Int. J. Mol. Med. 2018, 42, 182–192. [Google Scholar] [CrossRef]
- Carloni, S.; Proietti, F.; Rocchi, M.; Longini, M.; Marseglia, L.; D’Angelo, G.; Balduini, W.; Gitto, E.; Buonocore, G. Melatonin pharmacokinetics following oral administration in preterm neonates. Molecules 2017, 22, 2115. Available online: https://pubmed.ncbi.nlm.nih.gov/29194416 (accessed on 1 October 2022). [CrossRef]
- Yawno, T.; Mahen, M.; Li, J.; Fahey, M.C.; Jenkin, G.; Miller, S.L. The Beneficial Effects of Melatonin Administration Following Hypoxia-Ischemia in Preterm Fetal Sheep. Front. Cell. Neurosci. 2017, 11, 296. [Google Scholar] [CrossRef]
- Alonso-Alconada, D.; Alvarez, A.; Lacalle, J.; Hilario, E. Histological study of the protective effect of melatonin on neural cells after neonatal hypoxia-ischemia. Histol. Histopathol. 2012, 27, 771–783. [Google Scholar] [CrossRef]
- Robertson, N.J.; Faulkner, S.; Fleiss, B.; Bainbridge, A.; Andorka, C.; Price, D.; Powell, E.; Lecky-Thompson, L.; Thei, L.; Chandrasekaran, M.; et al. Melatonin augments hypothermic neuroprotection in a perinatal asphyxia model. Brain 2013, 136 Pt 1, 90–105. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Authors and Year | Type of Study | Aim of the Study | Outcomes and Conclusion |
|---|---|---|---|
| Tya Vine et al., 2022 | Review | To determinate the influence of melatonin on sleep during pregnancy |
|
| Ejaz H. et al., 2021 | Case controlled trial | To determine serum levels of melatonin and its major metabolite 6- hydroxymelatonin sulfate (6-OHMS) in normal pregnant women during each trimester of pregnancy, and immediately after delivery |
|
| Garofoli F et al., 2021 | Randomized controlled trial | The planned study will be the first aiming to evaluate the capacity of melatonin to mitigate brain impairment due to premature birth. |
|
| Laste G. et al., 2021 | Review | Investigating the relationship between melatonin and high-risk pregnancy. |
|
| Berbets AM et al., 2020 | Case control | To investigate whether the level of melatonin, cytokines, and PlGF in umbilical blood after birth is different in the case of IUGR compared to normal fetuses. |
|
| Swarnamani K et al., 2020 | A double-blind randomized placebo-controlled trial | to test the hypothesis that addition of melatonin will reduce the need for caesarean section. |
|
| Biran V. et al., 2019 | prospective cohort study | To determine serum levels of melatonin in different GW |
|
| De Almeida Chuffa et al., 2019 | Review | To focus on the main in vivo and in vitro functions of melatonin on uterine physiological processes, such as decidualization and implantation, and also on the feto-maternal tissues, and reviews how exogenous melatonin functions from a mechanistic standpoint to preserve the organ health. |
|
| Genario R. et al., 2019 | Review | To further investigate the evidence available on the effects of melatonin supplementation in animal and human studies, focusing on its potential application to gynecology. |
|
| Ji Yeon Lee MD et al., 2019 | Case controlled trial | To investigate the effects of maternally administered melatonin on preterm birth and perinatal brain injury in a mouse model of maternal inflammation. |
|
| Rahman SA et al., 2019 | Randomized case control | To evaluate the impact of light-induced modulation of melatonin secretion on uterine contractions in women during late third-trimester |
|
| Carlomagno G. et al., 2018 | Review | Positive effects on the outcomes of compromised pregnancies |
|
| Hobson SR et al., 2018 | Original Manuscript | To determine if melatonin could be a useful adjuvant therapy |
|
| Gene Chi Wai et al., 2017 | Review | To provide an overview, from an immunological perspective, on the roles of circadian clock and melatonin in pregnancy. |
|
| Katzer D. et al., 2016 | Case controlled trial | To evaluate the antioxidative capacity in breast milk and its regulation by time of day |
|
| YU WEN LIN. et al., 2018 | Case controlled trial | They evaluated whether melatonin inhibits ER stress in cultured neurons exposed to oxygen and glucose deprivation and in rats subjected to transient focal cerebral ischemia |
|
| Serum melatonin concentration | |
| I TRIMESTER OF PREGNANCY | Up to 611.4 pg/mL |
| II TRIMESTER OF PREGNANCY | Up to 1246.4pg/mL |
| III TRIMESTER OF PREGNANCY | Up to 1372 pg/mL (Ejaz H. et al., 2021) |
| AT DELIVERY | Up to 158 pg/mL (Biran V., 2019) |
| Serum melatonin concentration | |
| NEWBORN AT BIRTH | Up to 184 pg/mL |
| NEWBORN ON 3 DAYS AFTER BIRTH | Up to 75 pg/mL (Biran V., 2019) |
| Milk melatonin concentration | |
| MOTHER MILK | Up to 20 pg/mL (Biran V., 2019) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Verteramo, R.; Pierdomenico, M.; Greco, P.; Milano, C. The Role of Melatonin in Pregnancy and the Health Benefits for the Newborn. Biomedicines 2022, 10, 3252. https://doi.org/10.3390/biomedicines10123252
Verteramo R, Pierdomenico M, Greco P, Milano C. The Role of Melatonin in Pregnancy and the Health Benefits for the Newborn. Biomedicines. 2022; 10(12):3252. https://doi.org/10.3390/biomedicines10123252
Chicago/Turabian StyleVerteramo, Rosita, Matteo Pierdomenico, Pantaleo Greco, and Carmelia Milano. 2022. "The Role of Melatonin in Pregnancy and the Health Benefits for the Newborn" Biomedicines 10, no. 12: 3252. https://doi.org/10.3390/biomedicines10123252
APA StyleVerteramo, R., Pierdomenico, M., Greco, P., & Milano, C. (2022). The Role of Melatonin in Pregnancy and the Health Benefits for the Newborn. Biomedicines, 10(12), 3252. https://doi.org/10.3390/biomedicines10123252