

Genomic and Immune Approach in Platinum Refractory HPV-Negative Head and Neck Squamous Cell Carcinoma Patients Treated with Immunotherapy: A Novel Combined Profile
 , , , ,
, , , ,  , , , ,
, , , ,  , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Patient Enrolment
2.2. Samples Collection
2.3. Circulating Soluble Molecules
2.4. Statistical Analysis
2.5. Connectivity Analysis
2.6. Comprehensive Genomic Cancer Profiling
3. Results
3.1. Outcomes
3.2. Early Progressive Disease (Cases 1–5)
3.3. Responders Patients (Cases 6–10)
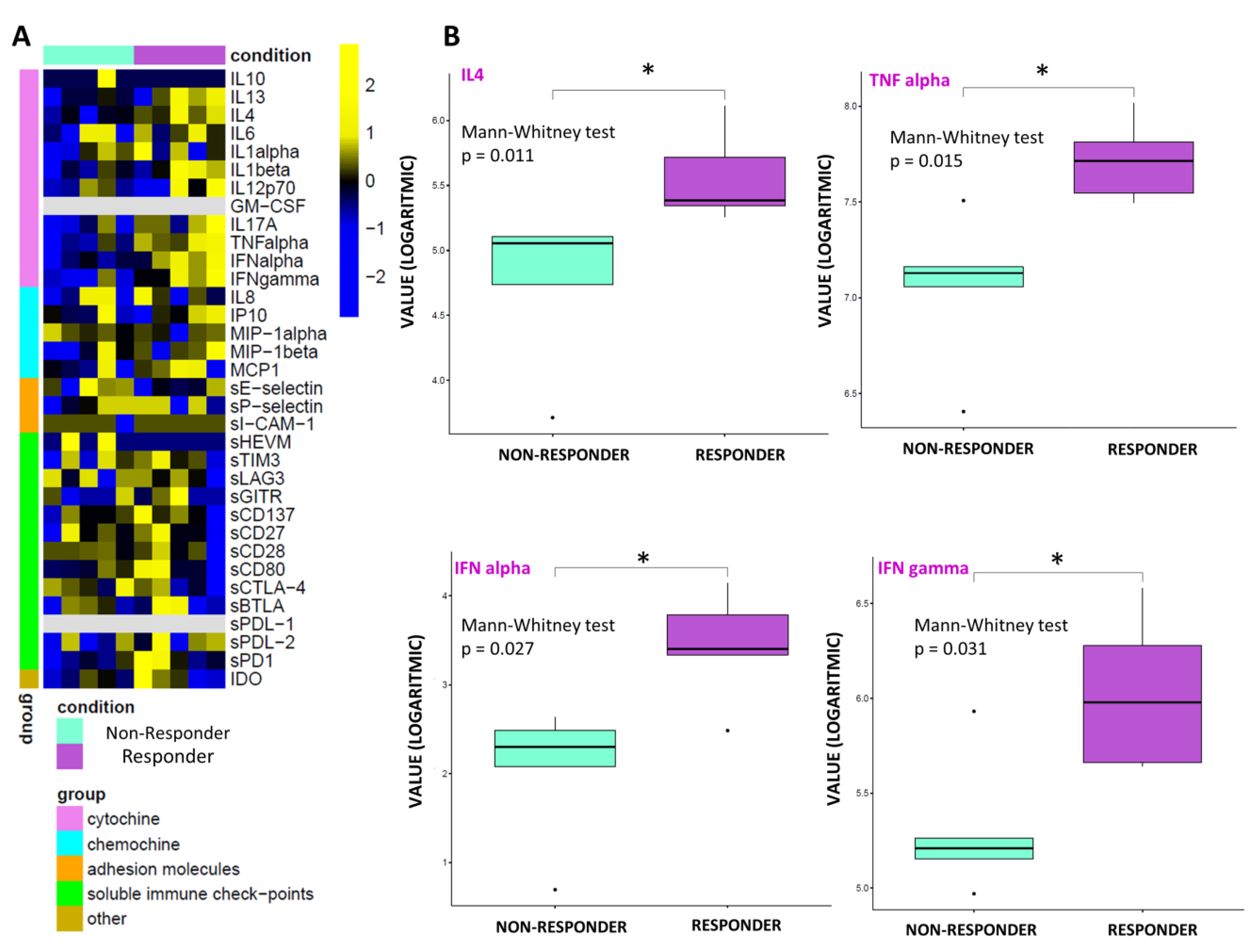
3.4. Statistical and Connectivity Analysis of Circulating Molecules
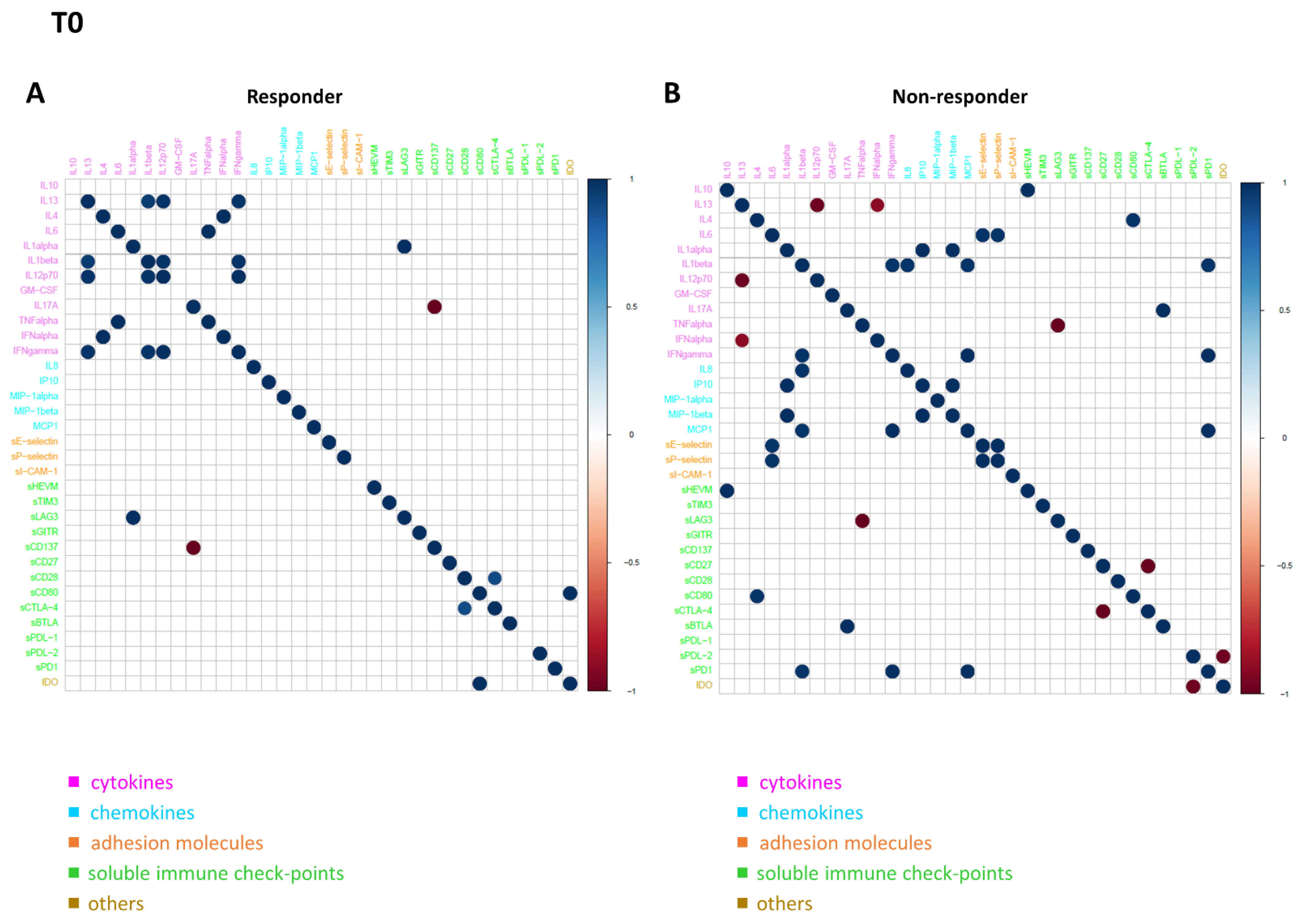
3.5. Connectivity Network Analysis between Circulating Molecules in Responder and Nonresponder Patients
4. Discussion
4.1. Soluble Immune Profile
4.2. Genomic Profile
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Ferris, R.L. Immunology and Immunotherapy of Head and Neck Cancer. J. Clin. Oncol. 2015, 33, 3293–3304. [Google Scholar] [CrossRef]
- Taube, J.M.; Klein, A.; Brahmer, J.R.; Xu, H.; Pan, X.; Kim, J.H.; Chen, L.; Pardoll, D.M.; Topalian, S.L.; Anders, R.A. Association of PD-1, PD-1 Ligands, and Other Features of the Tumor Immune Microenvironment with Response to Anti-PD-1 Therapy. Clin. Cancer Res. 2014, 20, 5064–5074. [Google Scholar] [CrossRef] [PubMed]
- Botticelli, A.; Cirillo, A.; Scagnoli, S.; Cerbelli, B.; Strigari, L.; Cortellini, A.; Pizzuti, L.; Vici, P.; De Galitiis, F.; Di Pietro, F.R.; et al. The Agnostic Role of Site of Metastasis in Predicting Outcomes in Cancer Patients Treated with Immunotherapy. Vaccines 2020, 8, 203. [Google Scholar] [CrossRef] [PubMed]
- Botticelli, A.; Mezi, S.; Pomati, G.; Cerbelli, B.; Di Rocco, C.; Amirhassankhani, S.; Sirgiovanni, G.; Occhipinti, M.; Napoli, V.; Emiliani, A.; et al. The 5-Ws of Immunotherapy in Head and Neck Cancer. Crit. Rev. Oncol. Hematol. 2020, 153, 103041. [Google Scholar] [CrossRef]
- De Felice, F.; Polimeni, A.; Tombolini, V. Human Papillomavirus (HPV) Vaccine and HPV-Related Head and Neck Cancer: What’s next? Oral Oncol. 2019, 93, 125–126. [Google Scholar] [CrossRef]
- Botticelli, A.; Cirillo, A.; Strigari, L.; Valentini, F.; Cerbelli, B.; Scagnoli, S.; Cerbelli, E.; Zizzari, I.G.; Rocca, C.D.; D’Amati, G.; et al. Anti–PD-1 and Anti–PD-L1 in Head and Neck Cancer: A Network Meta-Analysis. Front. Immunol. 2021, 12, 705096. [Google Scholar] [CrossRef] [PubMed]
- Ferris, R.L.; Blumenschein, G.; Fayette, J.; Guigay, J.; Colevas, A.D.; Licitra, L.; Harrington, K.J.; Kasper, S.; Vokes, E.E.; Even, C.; et al. Nivolumab vs Investigator’s Choice in Recurrent or Metastatic Squamous Cell Carcinoma of the Head and Neck: 2-Year Long-Term Survival Update of CheckMate 141 with Analyses by Tumor PD-L1 Expression. Oral Oncol. 2018, 81, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Cohen, E.E.W.; Soulières, D.; Le Tourneau, C.; Dinis, J.; Licitra, L.; Ahn, M.-J.; Soria, A.; Machiels, J.-P.; Mach, N.; Mehra, R.; et al. Pembrolizumab versus Methotrexate, Docetaxel, or Cetuximab for Recurrent or Metastatic Head-and-Neck Squamous Cell Carcinoma (KEYNOTE-040): A Randomised, Open-Label, Phase 3 Study. Lancet 2019, 393, 156–167. [Google Scholar] [CrossRef]
- Burtness, B.; Harrington, K.J.; Greil, R.; Soulières, D.; Tahara, M.; de Castro, G.; Psyrri, A.; Basté, N.; Neupane, P.; Bratland, Å.; et al. Pembrolizumab Alone or with Chemotherapy versus Cetuximab with Chemotherapy for Recurrent or Metastatic Squamous Cell Carcinoma of the Head and Neck (KEYNOTE-048): A Randomised, Open-Label, Phase 3 Study. Lancet 2019, 394, 1915–1928. [Google Scholar] [CrossRef]
- Moller, G.; Moller, E. The Concept of Immunological Surveillance against Neoplasia. Immunol. Rev. 1976, 28, 3–17. [Google Scholar] [CrossRef]
- Tenore, G.; Nuvoli, A.; Mohsen, A.; Cassoni, A.; Battisti, A.; Terenzi, V.; Della Monaca, M.; Raponi, I.; Brauner, E.; De Felice, F.; et al. Tobacco, Alcohol and Family History of Cancer as Risk Factors of Oral Squamous Cell Carcinoma: Case-Control Retrospective Study. Appl. Sci. 2020, 10, 3896. [Google Scholar] [CrossRef]
- Bai, R.; Chen, N.; Li, L.; Du, N.; Bai, L.; Lv, Z.; Tian, H.; Cui, J. Mechanisms of Cancer Resistance to Immunotherapy. Front. Oncol. 2020, 10, 1290. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, S.; Bhattacharya-Chatterjee, M.; O’Malley, B.W.; Chatterjee, S.K. Inhibition of NK Cell Activity through TGF-Β1 by Down-Regulation of NKG2D in a Murine Model of Head and Neck Cancer. J. Immunol. 2005, 175, 5541–5550. [Google Scholar] [CrossRef] [PubMed]
- Ferris, R.L.; Whiteside, T.L.; Ferrone, S. Immune Escape Associated with Functional Defects in Antigen-Processing Machinery in Head and Neck Cancer. Clin. Cancer Res. 2006, 12, 3890–3895. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, T.K.; Dworacki, G.; Tsukihiro, T.; Meidenbauer, N.; Gooding, W.; Johnson, J.T.; Whiteside, T.L. Spontaneous Apoptosis of Circulating T Lymphocytes in Patients with Head and Neck Cancer and Its Clinical Importance. Clin. Cancer Res. 2002, 8, 2553–2562. [Google Scholar]
- Shang, B.; Liu, Y.; Jiang, S.; Liu, Y. Prognostic Value of Tumor-Infiltrating FoxP3+ Regulatory T Cells in Cancers: A Systematic Review and Meta-Analysis. Sci. Rep. 2015, 5, 15179. [Google Scholar] [CrossRef]
- Badoual, C.; Hans, S.; Rodriguez, J.; Peyrard, S.; Klein, C.; Agueznay, N.E.H.; Mosseri, V.; Laccourreye, O.; Bruneval, P.; Fridman, W.H.; et al. Prognostic Value of Tumor-Infiltrating CD4+ T-Cell Subpopulations in Head and Neck Cancers. Clin. Cancer Res. 2006, 12, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Weber, F. Lymphocytes and Thyroid Cancer: More to It than Meets the Eye? Endocr.-Relat. Cancer 2014, 21, C1–C5. [Google Scholar] [CrossRef]
- Guigay, J.; Fayette, J.; Mesia, R.; Saada-Bouzid, E.; Lafond, C.; Geoffrois, L.; Martin, L.; Capitain, O.; Cupissol, D.; Castanie, H.; et al. TPExtreme Randomized Trial: Quality of Life (QoL) and Survival According to Second-Line Treatments in Patients with Recurrent/Metastatic Head and Neck Squamous Cell Carcinoma (R/M HNSCC). J. Clin. Oncol. 2020, 38, 6507. [Google Scholar] [CrossRef]
- Botticelli, A.; Mezi, S.; Pomati, G.; Sciattella, P.; Cerbelli, B.; Roberto, M.; Mammone, G.; Cirillo, A.; Cassano, A.; Di Dio, C.; et al. The Impact of Locoregional Treatment on Response to Nivolumab in Advanced Platinum Refractory Head and Neck Cancer: The Need Trial. Vaccines 2020, 8, 191. [Google Scholar] [CrossRef]
- Botticelli, A.; Cirillo, A.; Pomati, G.; Cerbelli, B.; Scagnoli, S.; Roberto, M.; Gelibter, A.; Mammone, G.; Calandrella, M.L.; Cerbelli, E.; et al. The Role of Opioids in Cancer Response to Immunotherapy. J. Transl. Med. 2021, 19, 119. [Google Scholar] [CrossRef]
- Thommen, D.S.; Schreiner, J.; Müller, P.; Herzig, P.; Roller, A.; Belousov, A.; Umana, P.; Pisa, P.; Klein, C.; Bacac, M.; et al. Progression of Lung Cancer Is Associated with Increased Dysfunction of T Cells Defined by Coexpression of Multiple Inhibitory Receptors. Cancer Immunol. Res. 2015, 3, 1344–1355. [Google Scholar] [CrossRef]
- Hanna, G.J.; Lizotte, P.; Cavanaugh, M.; Kuo, F.C.; Shivdasani, P.; Frieden, A.; Chau, N.G.; Schoenfeld, J.D.; Lorch, J.H.; Uppaluri, R.; et al. Frameshift Events Predict Anti–PD-1/L1 Response in Head and Neck Cancer. JCI Insight 2018, 3, e98811. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, Q.; Shi, B.; Xu, P.; Hu, Z.; Bai, L.; Zhang, X. Development of a Sandwich ELISA for Evaluating Soluble PD-L1 (CD274) in Human Sera of Different Ages as Well as Supernatants of PD-L1+ Cell Lines. Cytokine 2011, 56, 231–238. [Google Scholar] [CrossRef]
- Briukhovetska, D.; Dörr, J.; Endres, S.; Libby, P.; Dinarello, C.A.; Kobold, S. Interleukins in Cancer: From Biology to Therapy. Nat. Rev. Cancer 2021, 21, 481–499. [Google Scholar] [CrossRef] [PubMed]
- Nagarsheth, N.; Wicha, M.S.; Zou, W. Chemokines in the Cancer Microenvironment and Their Relevance in Cancer Immunotherapy. Nat. Rev. Immunol. 2017, 17, 559–572. [Google Scholar] [CrossRef] [PubMed]
- Guerrouahen, B.S.; Maccalli, C.; Cugno, C.; Rutella, S.; Akporiaye, E.T. Reverting Immune Suppression to Enhance Cancer Immunotherapy. Front. Oncol. 2019, 9, 1554. [Google Scholar] [CrossRef] [PubMed]
- Daassi, D.; Mahoney, K.M.; Freeman, G.J. The Importance of Exosomal PDL1 in Tumour Immune Evasion. Nat. Rev. Immunol. 2020, 20, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Zizzari, I.G.; Di Filippo, A.; Scirocchi, F.; Di Pietro, F.R.; Rahimi, H.; Ugolini, A.; Scagnoli, S.; Vernocchi, P.; Del Chierico, F.; Putignani, L.; et al. Soluble Immune Checkpoints, Gut Metabolites and Performance Status as Parameters of Response to Nivolumab Treatment in NSCLC Patients. J. Pers. Med. 2020, 10, 208. [Google Scholar] [CrossRef]
- Ugurel, S.; Schadendorf, D.; Horny, K.; Sucker, A.; Schramm, S.; Utikal, J.; Pföhler, C.; Herbst, R.; Schilling, B.; Blank, C.; et al. Elevated Baseline Serum PD-1 or PD-L1 Predicts Poor Outcome of PD-1 Inhibition Therapy in Metastatic Melanoma. Ann. Oncol. 2020, 31, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhang, J.; Tu, H.; Liang, D.; Chang, D.W.; Ye, Y.; Wu, X. Soluble Immune Checkpoint-Related Proteins as Predictors of Tumor Recurrence, Survival, and T Cell Phenotypes in Clear Cell Renal Cell Carcinoma Patients. J. Immunother. Cancer 2019, 7, 334. [Google Scholar] [CrossRef]
- Le, D.T.; Uram, J.N.; Wang, H.; Bartlett, B.R.; Kemberling, H.; Eyring, A.D.; Skora, A.D.; Luber, B.S.; Azad, N.S.; Laheru, D.; et al. PD-1 Blockade in Tumors with Mismatch-Repair Deficiency. N. Engl. J. Med. 2015, 372, 2509–2520. [Google Scholar] [CrossRef] [PubMed]
- De Schutter, H.; Spaepen, M.; Van Opstal, S.; Vander Poorten, V.; Verbeken, E.; Nuyts, S. The Prevalence of Microsatellite Instability in Head and Neck Squamous Cell Carcinoma. J. Cancer Res. Clin. Oncol. 2009, 135, 485–490. [Google Scholar] [CrossRef] [PubMed]
- Le, D.T.; Durham, J.N.; Smith, K.N.; Wang, H.; Bartlett, B.R.; Aulakh, L.K.; Lu, S.; Kemberling, H.; Wilt, C.; Luber, B.S.; et al. Mismatch Repair Deficiency Predicts Response of Solid Tumors to PD-1 Blockade. Science 2017, 357, 409–413. [Google Scholar] [CrossRef]
- Lawrence, M.S.; Stojanov, P.; Polak, P.; Kryukov, G.V.; Cibulskis, K.; Sivachenko, A.; Carter, S.L.; Stewart, C.; Mermel, C.H.; Roberts, S.A.; et al. Mutational Heterogeneity in Cancer and the Search for New Cancer-Associated Genes. Nature 2013, 499, 214–218. [Google Scholar] [CrossRef]
- Skoulidis, F.; Goldberg, M.E.; Greenawalt, D.M.; Hellmann, M.D.; Awad, M.M.; Gainor, J.F.; Schrock, A.B.; Hartmaier, R.J.; Trabucco, S.E.; Gay, L.; et al. STK11/LKB1 Mutations and PD-1 Inhibitor Resistance in KRAS -Mutant Lung Adenocarcinoma. Cancer Discov. 2018, 8, 822–835. [Google Scholar] [CrossRef]
- Chung, C.H.; Parker, J.S.; Karaca, G.; Wu, J.; Funkhouser, W.K.; Moore, D.; Butterfoss, D.; Xiang, D.; Zanation, A.; Yin, X.; et al. Molecular Classification of Head and Neck Squamous Cell Carcinomas Using Patterns of Gene Expression. Cancer Cell 2004, 5, 489–500. [Google Scholar] [CrossRef]
- Walter, V.; Yin, X.; Wilkerson, M.D.; Cabanski, C.R.; Zhao, N.; Du, Y.; Ang, M.K.; Hayward, M.C.; Salazar, A.H.; Hoadley, K.A.; et al. Molecular Subtypes in Head and Neck Cancer Exhibit Distinct Patterns of Chromosomal Gain and Loss of Canonical Cancer Genes. PLoS ONE 2013, 8, e56823. [Google Scholar] [CrossRef]
- Botticelli, A.; Mezi, S.; Pomati, G.; Cerbelli, B.; Cerbelli, E.; Roberto, M.; Giusti, R.; Cortellini, A.; Lionetto, L.; Scagnoli, S.; et al. Tryptophan Catabolism as Immune Mechanism of Primary Resistance to Anti-PD-1. Front. Immunol. 2020, 11, 1243. [Google Scholar] [CrossRef]
- Schrock, A.B.; Frampton, G.M.; Herndon, D.; Greenbowe, J.R.; Wang, K.; Lipson, D.; Yelensky, R.; Chalmers, Z.R.; Chmielecki, J.; Elvin, J.A.; et al. Comprehensive Genomic Profiling Identifies Frequent Drug-Sensitive EGFR Exon 19 Deletions in NSCLC Not Identified by Prior Molecular Testing. Clin. Cancer Res. 2016, 22, 3281–3285. [Google Scholar] [CrossRef] [PubMed]
- Hirshfield, K.M.; Tolkunov, D.; Zhong, H.; Ali, S.M.; Stein, M.N.; Murphy, S.; Vig, H.; Vazquez, A.; Glod, J.; Moss, R.A.; et al. Clinical Actionability of Comprehensive Genomic Profiling for Management of Rare or Refractory Cancers. Oncologist 2016, 21, 1315–1325. [Google Scholar] [CrossRef] [PubMed]
- Rozenblum, A.B.; Ilouze, M.; Dudnik, E.; Dvir, A.; Soussan-Gutman, L.; Geva, S.; Peled, N. Clinical Impact of Hybrid Capture–Based Next-Generation Sequencing on Changes in Treatment Decisions in Lung Cancer. J. Thorac. Oncol. 2017, 12, 258–268. [Google Scholar] [CrossRef] [PubMed]
- Frampton, G.M.; Fichtenholtz, A.; Otto, G.A.; Wang, K.; Downing, S.R.; He, J.; Schnall-Levin, M.; White, J.; Sanford, E.M.; An, P.; et al. Development and Validation of a Clinical Cancer Genomic Profiling Test Based on Massively Parallel DNA Sequencing. Nat. Biotechnol. 2013, 31, 1023–1031. [Google Scholar] [CrossRef] [PubMed]
- Piasecka, B.; Duffy, D.; Urrutia, A.; Quach, H.; Patin, E.; Posseme, C.; Bergstedt, J.; Charbit, B.; Rouilly, V.; MacPherson, C.R.; et al. Distinctive Roles of Age, Sex, and Genetics in Shaping Transcriptional Variation of Human Immune Responses to Microbial Challenges. Proc. Natl. Acad. Sci. USA 2018, 115, E488–E497. [Google Scholar] [CrossRef] [PubMed]
- Patin, E.; Hasan, M.; Bergstedt, J.; Rouilly, V.; Libri, V.; Urrutia, A.; Alanio, C.; Scepanovic, P.; Hammer, C.; Jönsson, F.; et al. Natural Variation in the Parameters of Innate Immune Cells Is Preferentially Driven by Genetic Factors. Nat. Immunol. 2018, 19, 302–314. [Google Scholar] [CrossRef]
- Wynn, T.A. IL-13 Effector Functions. Annu. Rev. Immunol. 2003, 21, 425–456. [Google Scholar] [CrossRef]
- Brevi, A.; Cogrossi, L.L.; Grazia, G.; Masciovecchio, D.; Impellizzieri, D.; Lacanfora, L.; Grioni, M.; Bellone, M. Much More Than IL-17A: Cytokines of the IL-17 Family Between Microbiota and Cancer. Front. Immunol. 2020, 11, 565470. [Google Scholar] [CrossRef]
- Gough, P.; Myles, I.A. Tumor Necrosis Factor Receptors: Pleiotropic Signaling Complexes and Their Differential Effects. Front. Immunol. 2020, 11, 585880. [Google Scholar] [CrossRef]
- Kelly-Welch, A.; Hanson, E.M.; Keegan, A.D. Interleukin-4 (IL-4) Pathway. Sci. STKE 2005, 2005, 293. [Google Scholar] [CrossRef]
- Ghiasi, H.; Cai, S.; Slanina, S.M.; Perng, G.; Nesburn, A.B.; Wechsler, S.L. The Role of Interleukin (IL)-2 and IL-4 in Herpes Simplex Virus Type 1 Ocular Replication and Eye Disease. J. Infect. Dis. 1999, 179, 1086–1093. [Google Scholar] [CrossRef] [PubMed]
- Hirata, A.; Hashimoto, H.; Shibasaki, C.; Narumi, K.; Aoki, K. Intratumoral IFN-α Gene Delivery Reduces Tumor-Infiltrating Regulatory T Cells through the Downregulation of Tumor CCL17 Expression. Cancer Gene Ther. 2019, 26, 334–343. [Google Scholar] [CrossRef] [PubMed]
- Schroder, K.; Hertzog, P.J.; Ravasi, T.; Hume, D.A. Interferon-γ: An Overview of Signals, Mechanisms and Functions. J. Leukoc. Biol. 2004, 75, 163–189. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, T.N.; Thommen, D.S. Tertiary Lymphoid Structures in Cancer. Science 2022, 375, eabf9419. [Google Scholar] [CrossRef]
- Wirsing, A.M.; Ervik, I.K.; Seppola, M.; Uhlin-Hansen, L.; Steigen, S.E.; Hadler-Olsen, E. Presence of High-Endothelial Venules Correlates with a Favorable Immune Microenvironment in Oral Squamous Cell Carcinoma. Mod. Pathol. 2018, 31, 910–922. [Google Scholar] [CrossRef]
- Sautès-Fridman, C.; Petitprez, F.; Calderaro, J.; Fridman, W.H. Tertiary Lymphoid Structures in the Era of Cancer Immunotherapy. Nat. Rev. Cancer 2019, 19, 307–325. [Google Scholar] [CrossRef]
- Leung, A.M.; Lee, A.F.; Ozao-Choy, J.; Ramos, R.I.; Hamid, O.; O’Day, S.J.; Shin-Sim, M.; Morton, D.L.; Faries, M.B.; Sieling, P.A.; et al. Clinical Benefit from Ipilimumab Therapy in Melanoma Patients May Be Associated with Serum CTLA4 Levels. Front. Oncol. 2014, 4, 110. [Google Scholar] [CrossRef]
- Zizzari, I.G.; Napoletano, C.; Di Filippo, A.; Botticelli, A.; Gelibter, A.; Calabrò, F.; Rossi, E.; Schinzari, G.; Urbano, F.; Pomati, G.; et al. Exploratory Pilot Study of Circulating Biomarkers in Metastatic Renal Cell Carcinoma. Cancers 2020, 12, 2620. [Google Scholar] [CrossRef]
- Lee, M.; Samstein, R.M.; Valero, C.; Chan, T.A.; Morris, L.G.T. Tumor Mutational Burden as a Predictive Biomarker for Checkpoint Inhibitor Immunotherapy. Hum. Vaccines Immunother. 2020, 16, 112–115. [Google Scholar] [CrossRef]
- Elmusrati, A.; Wang, J.; Wang, C.-Y. Tumor Microenvironment and Immune Evasion in Head and Neck Squamous Cell Carcinoma. Int. J. Oral Sci. 2021, 13, 24. [Google Scholar] [CrossRef]
- Chalmers, Z.R.; Connelly, C.F.; Fabrizio, D.; Gay, L.; Ali, S.M.; Ennis, R.; Schrock, A.; Campbell, B.; Shlien, A.; Chmielecki, J.; et al. Analysis of 100,000 Human Cancer Genomes Reveals the Landscape of Tumor Mutational Burden. Genome Med. 2017, 9, 34. [Google Scholar] [CrossRef] [PubMed]
- The Cancer Genome Atlas Network. Comprehensive Genomic Characterization of Head and Neck Squamous Cell Carcinomas. Nature 2015, 517, 576–582. [Google Scholar] [CrossRef] [PubMed]
- Samstein, R.M.; Lee, C.-H.; Shoushtari, A.N.; Hellmann, M.D.; Shen, R.; Janjigian, Y.Y.; Barron, D.A.; Zehir, A.; Jordan, E.J.; Omuro, A.; et al. Tumor Mutational Load Predicts Survival after Immunotherapy across Multiple Cancer Types. Nat. Genet. 2019, 51, 202–206. [Google Scholar] [CrossRef] [PubMed]
- Seiwert, T.Y.; Zuo, Z.; Keck, M.K.; Khattri, A.; Pedamallu, C.S.; Stricker, T.; Brown, C.; Pugh, T.J.; Stojanov, P.; Cho, J.; et al. Integrative and Comparative Genomic Analysis of HPV-Positive and HPV-Negative Head and Neck Squamous Cell Carcinomas. Clin. Cancer Res. 2015, 21, 632–641. [Google Scholar] [CrossRef]
- Zhu, G.; Pan, C.; Bei, J.-X.; Li, B.; Liang, C.; Xu, Y.; Fu, X. Mutant P53 in Cancer Progression and Targeted Therapies. Front. Oncol. 2020, 10, 595187. [Google Scholar] [CrossRef]
- Ganci, F.; Sacconi, A.; Bossel Ben-Moshe, N.; Manciocco, V.; Sperduti, I.; Strigari, L.; Covello, R.; Benevolo, M.; Pescarmona, E.; Domany, E.; et al. Expression of TP53 Mutation-Associated MicroRNAs Predicts Clinical Outcome in Head and Neck Squamous Cell Carcinoma Patients. Ann. Oncol. 2013, 24, 3082–3088. [Google Scholar] [CrossRef]
- Shi, Q.; Xiao, K.; Wei, W.; Zhang, B.-Y.; Chen, C.; Xu, Y.; Chen, L.-N.; Song, Y.-T.; Ma, X.; Zhang, N.-S.; et al. Associations of TP53 Mutations, Codon 72 Polymorphism and Human Papillomavirus in Head and Neck Squamous Cell Carcinoma Patients. Oncol. Rep. 2013, 30, 2811–2819. [Google Scholar] [CrossRef][Green Version]
- Zeng, X.-T.; Luo, W.; Geng, P.-L.; Guo, Y.; Niu, Y.-M.; Leng, W.-D. Association between the TP53 Codon 72 Polymorphism and Risk of Oral Squamous Cell Carcinoma in Asians: A Meta-Analysis. BMC Cancer 2014, 14, 469. [Google Scholar] [CrossRef]
- Zhang, Y.; Lin, A.; Li, Y.; Ding, W.; Meng, H.; Luo, P.; Zhang, J. Age and Mutations as Predictors of the Response to Immunotherapy in Head and Neck Squamous Cell Cancer. Front. Cell Dev. Biol. 2020, 8, 608969. [Google Scholar] [CrossRef]
- Blagih, J.; Buck, M.D.; Vousden, K.H. P53, Cancer and the Immune Response. J. Cell Sci. 2020, 133, jcs237453. [Google Scholar] [CrossRef]
- Gazzeri, S.; Gouyer, V.; Vour’ch, C.; Brambilla, C.; Brambilla, E. Mechanisms of P16INK4A Inactivation in Non Small-Cell Lung Cancers. Oncogene 1998, 16, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Sharpless, N.E. INK4a/ARF: A Multifunctional Tumor Suppressor Locus. Mutat. Res. Fundam. Mol. Mech. Mutagenesis 2005, 576, 22–38. [Google Scholar] [CrossRef] [PubMed]
- Chung, C.H.; Guthrie, V.B.; Masica, D.L.; Tokheim, C.; Kang, H.; Richmon, J.; Agrawal, N.; Fakhry, C.; Quon, H.; Subramaniam, R.M.; et al. Genomic Alterations in Head and Neck Squamous Cell Carcinoma Determined by Cancer Gene-Targeted Sequencing. Ann. Oncol. 2015, 26, 1216–1223. [Google Scholar] [CrossRef] [PubMed]
- Chau, N.G.; Li, Y.Y.; Jo, V.Y.; Rabinowits, G.; Lorch, J.H.; Tishler, R.B.; Margalit, D.N.; Schoenfeld, J.D.; Annino, D.J.; Goguen, L.A.; et al. Incorporation of Next-Generation Sequencing into Routine Clinical Care to Direct Treatment of Head and Neck Squamous Cell Carcinoma. Clin. Cancer Res. 2016, 22, 2939–2949. [Google Scholar] [CrossRef]
- Agrawal, N.; Frederick, M.J.; Pickering, C.R.; Bettegowda, C.; Chang, K.; Li, R.J.; Fakhry, C.; Xie, T.-X.; Zhang, J.; Wang, J.; et al. Exome Sequencing of Head and Neck Squamous Cell Carcinoma Reveals Inactivating Mutations in NOTCH1. Science 2011, 333, 1154–1157. [Google Scholar] [CrossRef] [PubMed]
- Stransky, N.; Egloff, A.M.; Tward, A.D.; Kostic, A.D.; Cibulskis, K.; Sivachenko, A.; Kryukov, G.V.; Lawrence, M.S.; Sougnez, C.; McKenna, A.; et al. The Mutational Landscape of Head and Neck Squamous Cell Carcinoma. Science 2011, 333, 1157–1160. [Google Scholar] [CrossRef]
- Kelliher, M.A.; Roderick, J.E. NOTCH Signaling in T-Cell-Mediated Anti-Tumor Immunity and T-Cell-Based Immunotherapies. Front. Immunol. 2018, 9, 1718. [Google Scholar] [CrossRef]
- Janghorban, M.; Xin, L.; Rosen, J.M.; Zhang, X.H.-F. Notch Signaling as a Regulator of the Tumor Immune Response: To Target or Not To Target? Front. Immunol. 2018, 9, 1649. [Google Scholar] [CrossRef]
- Shah, P.A.; Huang, C.; Li, Q.; Kazi, S.A.; Byers, L.A.; Wang, J.; Johnson, F.M.; Frederick, M.J. NOTCH1 Signaling in Head and Neck Squamous Cell Carcinoma. Cells 2020, 9, 2677. [Google Scholar] [CrossRef]
- Zhao, J.; Liang, Y.; Song, F.; Xu, S.; Nian, L.; Zhou, X.; Wang, S. TSG Attenuates LPC-Induced Endothelial Cells Inflammatory Damage through Notch Signaling Inhibition: TSG Attenuates LPC-Induced EC Damage Via Notch Inhibition. IUBMB Life 2016, 68, 37–50. [Google Scholar] [CrossRef]
- Fukusumi, T.; Guo, T.W.; Sakai, A.; Ando, M.; Ren, S.; Haft, S.; Liu, C.; Amornphimoltham, P.; Gutkind, J.S.; Califano, J.A. The NOTCH4-HEY1 Pathway Induces Epithelial–Mesenchymal Transition in Head and Neck Squamous Cell Carcinoma. Clin. Cancer Res. 2018, 24, 619–633. [Google Scholar] [CrossRef] [PubMed]
- Bao, B.; Wang, Z.; Ali, S.; Kong, D.; Li, Y.; Ahmad, A.; Banerjee, S.; Azmi, A.S.; Miele, L.; Sarkar, F.H. Retraction Notice to “Notch-1 Induces Epithelial-Mesenchymal Transition Consistent with Cancer Stem Cell Phenotype in Pancreatic Cancer Cells”. Cancer Lett. 2018, 423, 153. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Hong, X.; Song, Z.; Xu, Y.; Li, C.; Wang, G.; Zhang, Y.; Zhao, X.; Zhao, Z.; Zhao, J.; et al. Identification of Deleterious NOTCH Mutation as Novel Predictor to Efficacious Immunotherapy in NSCLC. Clin. Cancer Res. 2020, 26, 3649–3661. [Google Scholar] [CrossRef] [PubMed]
- Suda, T.; Hama, T.; Kondo, S.; Yuza, Y.; Yoshikawa, M.; Urashima, M.; Kato, T.; Moriyama, H. Copy Number Amplification of the PIK3CA Gene Is Associated with Poor Prognosis in Non-Lymph Node Metastatic Head and Neck Squamous Cell Carcinoma. BMC Cancer 2012, 12, 416. [Google Scholar] [CrossRef] [PubMed]
- Lui, V.W.Y.; Hedberg, M.L.; Li, H.; Vangara, B.S.; Pendleton, K.; Zeng, Y.; Lu, Y.; Zhang, Q.; Du, Y.; Gilbert, B.R.; et al. Frequent Mutation of the PI3K Pathway in Head and Neck Cancer Defines Predictive Biomarkers. Cancer Discov. 2013, 3, 761–769. [Google Scholar] [CrossRef] [PubMed]
- Welcsh, P.L. BRCA1 and BRCA2 and the Genetics of Breast and Ovarian Cancer. Hum. Mol. Genet. 2001, 10, 705–713. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Miki, Y. Role of BRCA1 and BRCA2 as Regulators of DNA Repair, Transcription, and Cell Cycle in Response to DNA Damage. Cancer Sci. 2004, 95, 866–871. [Google Scholar] [CrossRef]
- Paul, A. The Breast Cancer Susceptibility Genes (BRCA) in Breast and Ovarian Cancers. Front. Biosci. 2014, 19, 605. [Google Scholar] [CrossRef]
- Venkitaraman, A.R. How Do Mutations Affecting the Breast Cancer Genes BRCA1 and BRCA2 Cause Cancer Susceptibility? DNA Repair 2019, 81, 102668. [Google Scholar] [CrossRef]
- Li, J.; Wang, W.; Zhang, Y.; Cieślik, M.; Guo, J.; Tan, M.; Green, M.D.; Wang, W.; Lin, H.; Li, W.; et al. Epigenetic Driver Mutations in ARID1A Shape Cancer Immune Phenotype and Immunotherapy. J. Clin. Investig. 2020, 130, 2712–2726. [Google Scholar] [CrossRef]
- Tan, Y.S.; Sansanaphongpricha, K.; Xie, Y.; Donnelly, C.R.; Luo, X.; Heath, B.R.; Zhao, X.; Bellile, E.; Hu, H.; Chen, H.; et al. Mitigating SOX2-Potentiated Immune Escape of Head and Neck Squamous Cell Carcinoma with a STING-Inducing Nanosatellite Vaccine. Clin. Cancer Res. 2018, 24, 4242–4255. [Google Scholar] [CrossRef] [PubMed]
- Wuebben, E.L.; Rizzino, A. The Dark Side of SOX2: Cancer—A Comprehensive Overview. Oncotarget 2017, 8, 44917–44943. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | All Patients N 10 |
|---|---|
| Age (years) Median age (range) | 66 (46–76) |
| Gender Male Female | 8 2 |
| Baseline PS 1 0 1 | 2 8 |
| Risk factors Smoking history (SH) Alcohol abuse (AA) | 8 3 |
| Tumor Location Oral cavity Oropharynx Larynx | 4 1 5 |
| Histology Squamous cell carcinoma | 10 |
| Grading 2 3 | 4 6 |
| HPV 2 Positive Negative | 0 10 |
| Only recurrent disease Metastatic site ≥ 1 Previous I line (extreme) | 1 9 10 |
| Patients | Age | Primary Tumor | PS | Best Response | PFS (Months) | OS (Months) | Early Progression | Patient Status 1 | Genomic Signature |
|---|---|---|---|---|---|---|---|---|---|
| Case 1 | 62 | Oral cavity | 1 | PD | 2 | 6 | Yes | DOD | TMB-intermediate 8 mut/MB MSI-stable |
| Case 2 | 65 | Larynx | 1 | PD | 1 | 21 | Yes | ALIVE | TMB 6 muts/MB MSI-stable |
| Case 3 | 76 | Larynx | 1 | PD | 2 | 2 | Yes | DOD | TMB 6 muts/MB MSI-stable |
| Case 4 | 65 | Oral cavity | 1 | PD | 2 | 11 | Yes | DOD | TMB 3 muts/MB MSI-stable |
| Case 5 | 53 | Larynx | 1 | PD | 1 | 3 | Yes | DOD | TMB 4 muts/MB MSI-stable |
| Case 6 | 52 | Oropharynx | 1 | SD | 5 | 6 | No | DOD | TMB 5 muts/MB MSI-stable |
| Case 7 | 63 | Oral cavity | 1 | SD | 6 | 7 | No | DOD | TMB 3 muts/MB MSI-stable |
| Case 8 | 71(F) | Larynx | 0 | PR | 33 | 33 | No | ALIVE | TMB 30 muts/MB MSI-stable |
| Case 9 | 67 | Larynx | 0 | PR | 20 | 39 | No | ALIVE | TMB 15 muts/MB MSI-stable |
| Case 10 | 46(F) | Oral cavity | 1 | PR | 4 | 5 | No | DOD | TMB 1 muts/MB MSI-stable |
| Not Mutated | Case Report | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Mutated | # 1 | # 2 | # 3 | # 4 | # 5 | # 6 | # 7 | # 8 | # 9 | # 10 |
| TMB | 8 Muts/Mb | 6 Muts/Mb | 6 Muts/Mb | 3 Muts/Mb | 4 Muts/Mb | 5 Muts/Mb | 3 Muts/Mb | 30 Muts/Mb | 15 Muts/Mb | 1 Muts/Mb |
| Microsatellite Status | MS-Stable | MS-Stable | MS-Stable | MS-Stable | MS-Stable | MS-Stable | MS-Stable | MS-Stable | MS-Stable | MS-Stable |
| TP53 | ||||||||||
| BRCA1 | ||||||||||
| ARID1A | ||||||||||
| CDKN2A/B | ||||||||||
| BCL2L1 | ||||||||||
| NFE2L2 | ||||||||||
| NOTCH1 | ||||||||||
| CCND1 | ||||||||||
| ERBB2 | ||||||||||
| CDK6 | ||||||||||
| PIK3CA | ||||||||||
| SOX2 | ||||||||||
| EPHB4 | ||||||||||
| FGF19 | ||||||||||
| FGF3 | ||||||||||
| FGF4 | ||||||||||
| PRKCI | ||||||||||
| TERC | ||||||||||
| NOTCH2 | ||||||||||
| RB1 | ||||||||||
| SMAD4 | ||||||||||
| TERT | ||||||||||
| FGFR1 | ||||||||||
| CD22 | ||||||||||
| NSD3 | ||||||||||
| PRKCI | ||||||||||
| AKT2 | ||||||||||
| MTAP | ||||||||||
| SPEN | ||||||||||
| KDM6A | ||||||||||
| DNMT3A | ||||||||||
| CUL3 | ||||||||||
| PIK3R1 | ||||||||||
| BCORL1 | ||||||||||
| PTEN | ||||||||||
| FGF14 | ||||||||||
| NSD3 | ||||||||||
| PARK2 | ||||||||||
| ZNF703 | ||||||||||
| TET2 | ||||||||||
| PBRM1 | ||||||||||
| PIK3C2G | ||||||||||
| Soluble Molecules | Class of Molecules | Cell Source | Ligands | Main Function | Type of Action | References |
|---|---|---|---|---|---|---|
| IL13 | Cytokyne | T CD4 Cells CD8 cells NK B-cells Monocytes Eosinophils Mast cells | IL13Rα1 IL13Rα2 |
| proinflammatory | [47] |
| IL17A | Cytokine | Lymphocytes TCD4 Th17 | IL17Rα |
| proinflammatory | [48] |
| TNF alpha | Cytokine | Macrophages NK T cells | TNFR1 TNFR2 |
| Immune-activation/ pro-inflammatory | [49] |
| IL4 | Cytokine | T cells Mast cells | IL4-Rα |
| Pro-inflammatory | [50,51] |
| INF alpha | Cytokine | DC Macrophages NK cells Macrophages Endothelial cells Fibroblasts | INFαR1/2 |
| Pro-inflammatory immune-activation | [52] |
| INF gamma | Cytokine | Lymphocytes T (th1) CD8 and NK | INFγR1/2 |
| immunoactivating/ possible immunosuppressive activity) | [53] |
| Gene Alteration | Pathways | Mechanism Effect on Tumor Cells | Effect on Immune System and Response to Immunotherapy |
|---|---|---|---|
| TP53 | P53 |
|
|
| NOTCH1/2 | NOTCH pathway |
| Resistance Immune escape |
| CDKN2A/B | Rb/INK4/ARF |
| Conflicting evidence regarding the association between CDKN2A genomic alterations and response to ICIs |
| PIK3CA | PI3K |
|
|
| BRCA1/2 | DDR pathway |
|
|
| ARID1A | Encodes for switch/sucrose nonfermenting (SWI/SNF) chromatin remodeling complexes |
| Immune evasion |
| SOX2 | PI3K-AKT pathway |
| Immune evasion |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mezi, S.; Pomati, G.; Zizzari, I.G.; Di Filippo, A.; Cerbelli, B.; Cirillo, A.; Fiscon, G.; Amirhassankhani, S.; Valentini, V.; De Vincentiis, M.; et al. Genomic and Immune Approach in Platinum Refractory HPV-Negative Head and Neck Squamous Cell Carcinoma Patients Treated with Immunotherapy: A Novel Combined Profile. Biomedicines 2022, 10, 2732. https://doi.org/10.3390/biomedicines10112732
Mezi S, Pomati G, Zizzari IG, Di Filippo A, Cerbelli B, Cirillo A, Fiscon G, Amirhassankhani S, Valentini V, De Vincentiis M, et al. Genomic and Immune Approach in Platinum Refractory HPV-Negative Head and Neck Squamous Cell Carcinoma Patients Treated with Immunotherapy: A Novel Combined Profile. Biomedicines. 2022; 10(11):2732. https://doi.org/10.3390/biomedicines10112732
Chicago/Turabian StyleMezi, Silvia, Giulia Pomati, Ilaria Grazia Zizzari, Alessandra Di Filippo, Bruna Cerbelli, Alessio Cirillo, Giulia Fiscon, Sasan Amirhassankhani, Valentino Valentini, Marco De Vincentiis, and et al. 2022. "Genomic and Immune Approach in Platinum Refractory HPV-Negative Head and Neck Squamous Cell Carcinoma Patients Treated with Immunotherapy: A Novel Combined Profile" Biomedicines 10, no. 11: 2732. https://doi.org/10.3390/biomedicines10112732
APA StyleMezi, S., Pomati, G., Zizzari, I. G., Di Filippo, A., Cerbelli, B., Cirillo, A., Fiscon, G., Amirhassankhani, S., Valentini, V., De Vincentiis, M., Corsi, A., Di Gioia, C., Tombolini, V., Della Rocca, C., Polimeni, A., Nuti, M., Marchetti, P., & Botticelli, A. (2022). Genomic and Immune Approach in Platinum Refractory HPV-Negative Head and Neck Squamous Cell Carcinoma Patients Treated with Immunotherapy: A Novel Combined Profile. Biomedicines, 10(11), 2732. https://doi.org/10.3390/biomedicines10112732