




Next-Generation Sequencing Comparative Analysis of DNA Mutations between Blood-Derived Extracellular Vesicles and Matched Cancer Tissue in Patients with Grade 4 Glioblastoma

 , , , ,
, , , ,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
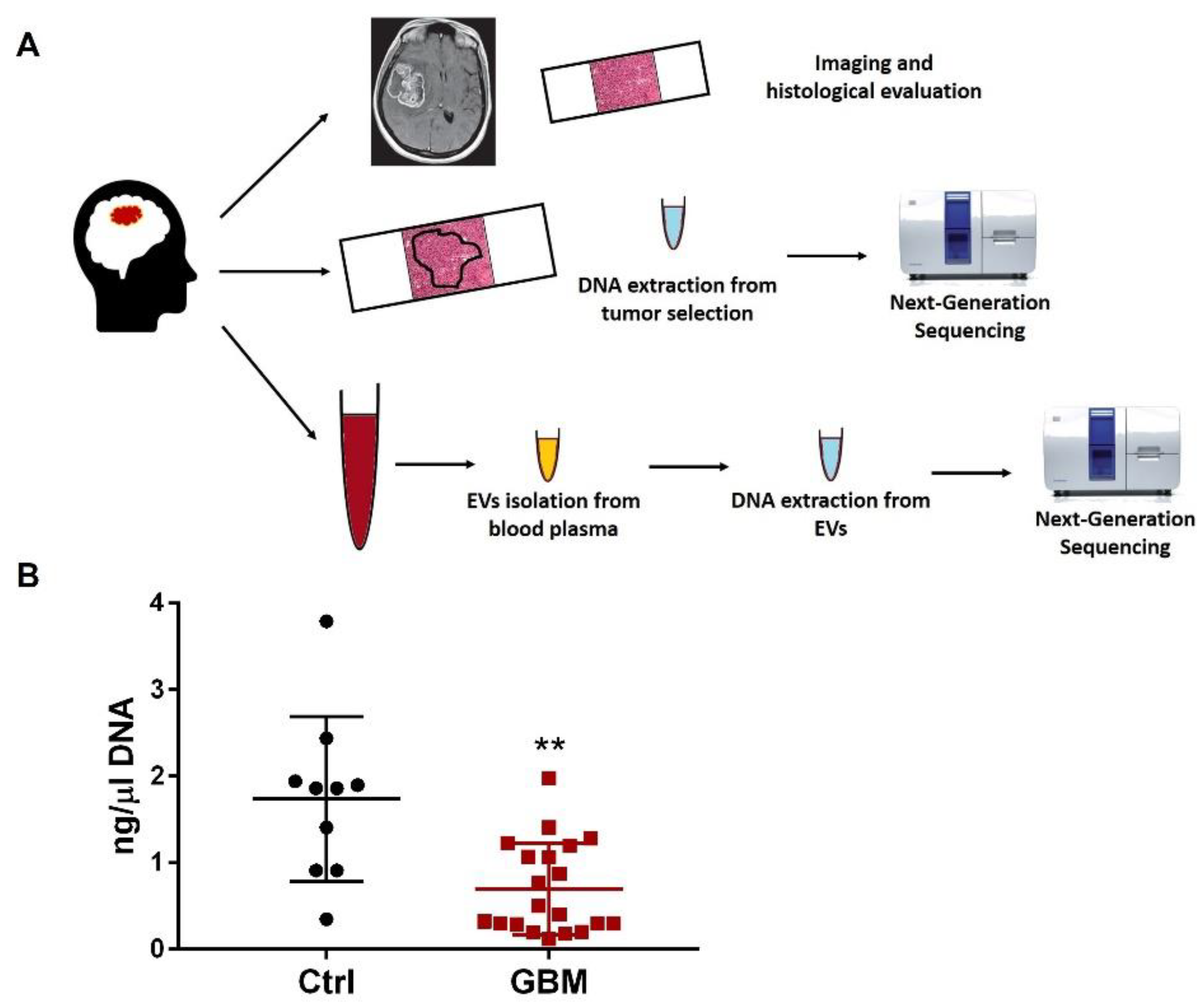
2.1. Study Design
2.2. Inclusion and Exclusion Criteria
2.3. Radiologic Assessment
2.4. Tissue Sampling and Histopathologic Analysis
2.5. Tissue Processing and DNA Extraction
2.6. Plasma Separation, EV Isolation and DNA Extraction
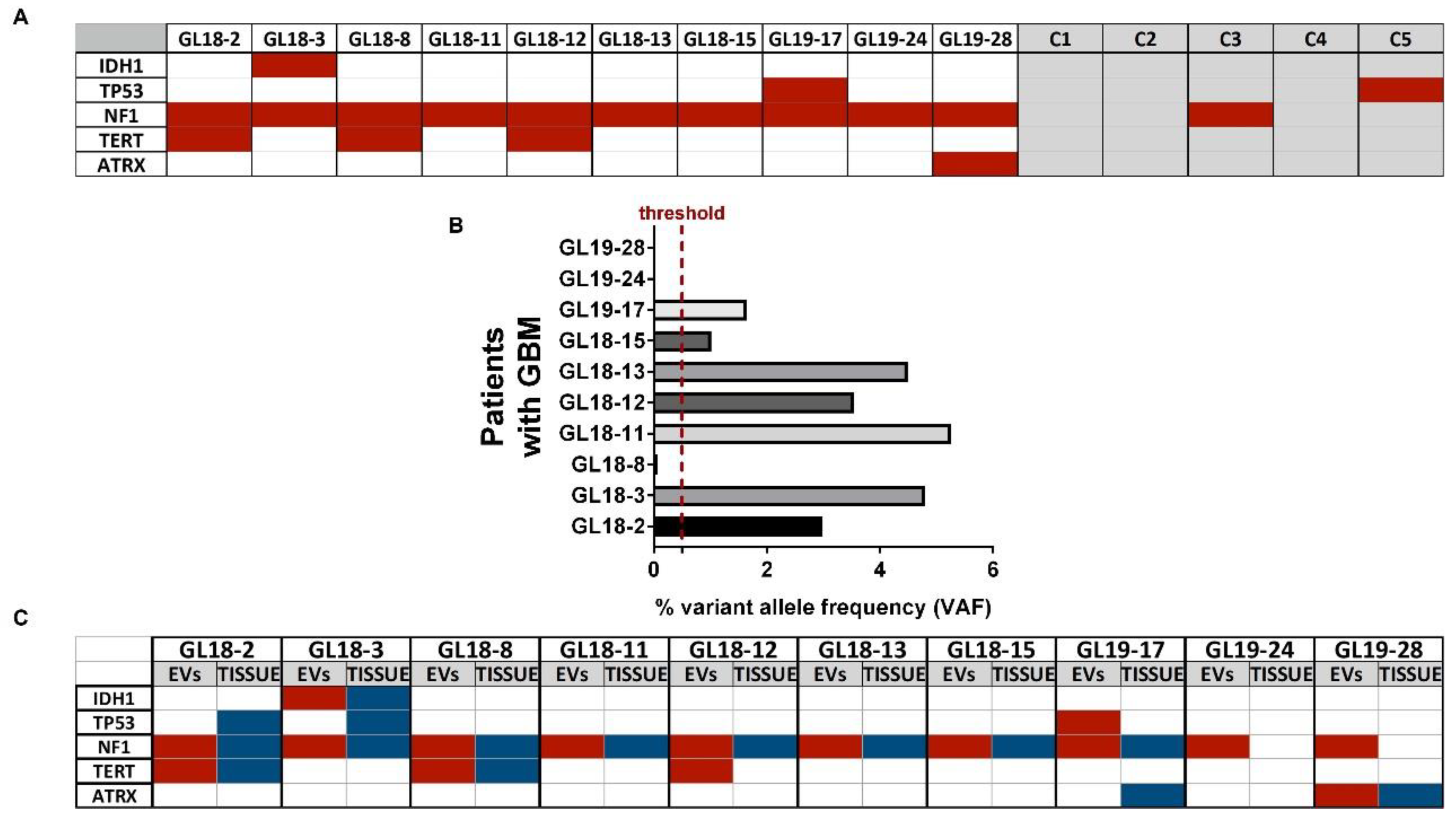
2.7. Next-Generation Sequencing Analysis
2.8. Statistical Analysis
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stupp, R.; Hegi, M.E.; Mason, W.P.; van den Bent, M.J.; Taphoorn, M.J.; Janzer, R.C.; Ludwin, S.K.; Allgeier, A.; Fisher, B.; Belanger, K.; et al. Effects of radiotherapy with concomitant and adjuvant temozolomide versus radiotherapy alone on survival in glioblastoma in a randomised phase III study: 5-year analysis of the EORTC-NCIC trial. Lancet Oncol. 2009, 10, 459–466. [Google Scholar] [CrossRef]
- Chinot, O.L.; Wick, W.; Mason, W.; Henriksson, R.; Saran, F.; Nishikawa, R.; Carpentier, A.F.; Hoang-Xuan, K.; Kavan, P.; Cernea, D.; et al. Bevacizumab plus radiotherapy-temozolomide for newly diagnosed glioblastoma. N. Engl. J. Med. 2014, 370, 709–722. [Google Scholar] [CrossRef] [Green Version]
- Stupp, R.; Mason, W.P.; van den Bent, M.J.; Weller, M.; Fisher, B.; Taphoorn, M.J.; Belanger, K.; Brandes, A.A.; Marosi, C.; Bogdahn, U.; et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N. Engl. J. Med. 2005, 352, 987–996. [Google Scholar] [CrossRef] [Green Version]
- Louis, D.N.; Perry, A.; Reifenberger, G.; von Deimling, A.; Figarella-Branger, D.; Cavenee, W.K.; Ohgaki, H.; Wiestler, O.D.; Kleihues, P.; Ellison, D.W. The 2016 World Health Organization Classification of Tumors of the Central Nervous System: A summary. Acta Neuropathol. 2016, 131, 803–820. [Google Scholar] [CrossRef] [Green Version]
- Rosa, P.; Scibetta, S.; Pepe, G.; Mangino, G.; Capocci, L.; Moons, S.J.; Boltje, T.J.; Fazi, F.; Petrozza, V.; Di Pardo, A.; et al. Polysialic Acid Sustains the Hypoxia-Induced Migration and Undifferentiated State of Human Glioblastoma Cells. Int. J. Mol. Sci. 2022, 23, 9563. [Google Scholar] [CrossRef]
- Louis, D.N.; Perry, A.; Wesseling, P.; Brat, D.J.; Cree, I.A.; Figarella-Branger, D.; Hawkins, C.; Ng, H.K.; Pfister, S.M.; Reifenberger, G.; et al. The 2021 WHO Classification of Tumors of the Central Nervous System: A summary. Neuro. Oncol. 2021, 23, 1231–1251. [Google Scholar] [CrossRef] [PubMed]
- Mangino, G.; Iuliano, M.; Carlomagno, S.; Bernardini, N.; Rosa, P.; Chiantore, M.V.; Skroza, N.; Calogero, A.; Potenza, C.; Romeo, G. Interleukin-17A affects extracellular vesicles release and cargo in human keratinocytes. Exp. Dermatol. 2019, 28, 1066–1073. [Google Scholar] [CrossRef] [PubMed]
- Bordin, A.; Chirivì, M.; Pagano, F.; Milan, M.; Iuliano, M.; Scaccia, E.; Fortunato, O.; Mangino, G.; Dhori, X.; de Marinis, E.; et al. Human platelet lysate derived extracellular vesicles enhance angiogenesis through miR-126. Cell Prolif. 2022, e13312. [Google Scholar] [CrossRef] [PubMed]
- Llorente, A.; Skotland, T.; Sylvanne, T.; Kauhanen, D.; Rog, T.; Orlowski, A.; Vattulainen, I.; Ekroos, K.; Sandvig, K. Molecular lipidomics of exosomes released by PC-3 prostate cancer cells. Biochim. Biophys. Acta 2013, 1831, 1302–1309. [Google Scholar] [CrossRef] [PubMed]
- Hallal, S.; Azimi, A.; Wei, H.; Ho, N.; Lee, M.Y.T.; Sim, H.W.; Sy, J.; Shivalingam, B.; Buckland, M.E.; Alexander-Kaufman, K.L. A Comprehensive Proteomic SWATH-MS Workflow for Profiling Blood Extracellular Vesicles: A New Avenue for Glioma Tumour Surveillance. Int. J. Mol. Sci. 2020, 21, 4754. [Google Scholar] [CrossRef] [PubMed]
- Thakur, B.K.; Zhang, H.; Becker, A.; Matei, I.; Huang, Y.; Costa-Silva, B.; Zheng, Y.; Hoshino, A.; Brazier, H.; Xiang, J.; et al. Double-stranded DNA in exosomes: A novel biomarker in cancer detection. Cell Res. 2014, 24, 766–769. [Google Scholar] [CrossRef] [Green Version]
- Giusti, I.; di Francesco, M.; Dolo, V. Extracellular Vesicles in Glioblastoma: Role in Biological Processes and in Therapeutic Applications. Curr. Cancer Drug Targets 2017, 17, 221–235. [Google Scholar] [CrossRef] [PubMed]
- Chistiakov, D.A.; Chekhonin, V.P. Extracellular vesicles shed by glioma cells: Pathogenic role and clinical value. Tumour Biol. 2014, 35, 8425–8438. [Google Scholar] [CrossRef] [PubMed]
- Van der Vos, K.E.; Balaj, L.; Skog, J.; Breakefield, X.O. Brain tumor microvesicles: Insights into intercellular communication in the nervous system. Cell Mol. Neurobiol. 2011, 31, 949–959. [Google Scholar] [CrossRef] [Green Version]
- Shiras, A.; Mondal, A. Extrachromosomal DNA: Redefining the pathogenesis of glioblastoma. Biochim. Biophys. Acta Rev. Cancer 2021, 1876, 188551. [Google Scholar] [CrossRef]
- Figueroa, J.M.; Skog, J.; Akers, J.; Li, H.; Komotar, R.; Jensen, R.; Ringel, F.; Yang, I.; Kalkanis, S.; Thompson, R.; et al. Detection of wild-type EGFR amplification and EGFRvIII mutation in CSF-derived extracellular vesicles of glioblastoma patients. Neuro Oncol. 2017, 19, 1494–1502. [Google Scholar] [CrossRef] [PubMed]
- Heitzer, E.; Haque, I.S.; Roberts, C.E.S.; Speicher, M.R. Current and future perspectives of liquid biopsies in genomics-driven oncology. Nat. Rev. Genet. 2019, 20, 71–88. [Google Scholar] [CrossRef]
- Andre-Gregoire, G.; Bidere, N.; Gavard, J. Temozolomide affects Extracellular Vesicles Released by Glioblastoma Cells. Biochimie 2018, 155, 11–15. [Google Scholar] [CrossRef]
- De Vrij, J.; Maas, S.L.; Kwappenberg, K.M.; Schnoor, R.; Kleijn, A.; Dekker, L.; Luider, T.M.; de Witte, L.D.; Litjens, M.; van Strien, M.E.; et al. Glioblastoma-derived extracellular vesicles modify the phenotype of monocytic cells. Int. J. Cancer 2015, 137, 1630–1642. [Google Scholar] [CrossRef]
- Naryzhny, S.; Volnitskiy, A.; Kopylov, A.; Zorina, E.; Kamyshinsky, R.; Bairamukov, V.; Garaeva, L.; Shlikht, A.; Shtam, T. Proteome of Glioblastoma-Derived Exosomes as a Source of Biomarkers. Biomedicines 2020, 8, 216. [Google Scholar] [CrossRef] [PubMed]
- Bagley, S.J.; Nabavizadeh, S.A.; Mays, J.J.; Till, J.E.; Ware, J.B.; Levy, S.; Sarchiapone, W.; Hussain, J.; Prior, T.; Guiry, S.; et al. Clinical Utility of Plasma Cell-Free DNA in Adult Patients with Newly Diagnosed Glioblastoma: A Pilot Prospective Study. Clin. Cancer Res. 2020, 26, 397–407. [Google Scholar] [CrossRef]
- Fraser, K.; Jo, A.; Giedt, J.; Vinegoni, C.; Yang, K.S.; Peruzzi, P.; Chiocca, E.A.; Breakefield, X.O.; Lee, H.; Weissleder, R. Characterization of single microvesicles in plasma from glioblastoma patients. Neuro Oncol. 2019, 21, 606–615. [Google Scholar] [CrossRef] [PubMed]
- Caruso, G.; Pacini, L.; Iossa, A.; di Cristofano, C.; Bastianelli, D.; Silecchia, G.; Mele, M.; Petrozza, V.; Calogero, A.; de Falco, E. A rare case of omental extra-gastrointestinal stromal tumor showing two coexisting mutations on exon 14 of the PDGFRA gene. Gastroenterol. Rep. 2021, 9, 377–379. [Google Scholar] [CrossRef]
- De Falco, E.; Pacini, L.; Bastianelli, D.; Spinelli, G.P.; Spoto, C.; Veltri, E.; Calogero, A. Concomitant Mutations G12D and G13D on the Exon 2 of the KRAS Gene: Two Cases of Women with Colon Adenocarcinoma. Diagnostics 2021, 11, 659. [Google Scholar] [CrossRef] [PubMed]
- Cenciarini, M.; Valentino, M.; Belia, S.; Sforna, L.; Rosa, P.; Ronchetti, S.; D’Adamo, M.C.; Pessia, M. Dexamethasone in Glioblastoma Multiforme Therapy: Mechanisms and Controversies. Front. Mol. Neurosci. 2019, 12, 65. [Google Scholar] [CrossRef]
- Lee, S.Y. Temozolomide resistance in glioblastoma multiforme. Genes Dis. 2016, 3, 198–210. [Google Scholar] [CrossRef] [Green Version]
- Rosa, P.; Catacuzzeno, L.; Sforna, L.; Mangino, G.; Carlomagno, S.; Mincione, G.; Petrozza, V.; Ragona, G.; Franciolini, F.; Calogero, A. BK channels blockage inhibits hypoxia-induced migration and chemoresistance to cisplatin in human glioblastoma cells. J. Cell. Physiol. 2018, 233, 6866–6877. [Google Scholar] [CrossRef]
- Noch, E.K.; Ramakrishna, R.; Magge, R. Challenges in the Treatment of Glioblastoma: Multisystem Mechanisms of Therapeutic Resistance. World Neurosurg. 2018, 116, 505–517. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, C.A.; Kaye, A.H.; Drummond, K.J.; Widodo, S.S.; Mantamadiotis, T.; Vella, L.J.; Stylli, S.S. Extracellular vesicles and their role in glioblastoma. Crit. Rev. Clin. Lab. Sci. 2019, 1–26. [Google Scholar] [CrossRef]
- Ho, J.; Chaiswing, L.; St Clair, D.K. Extracellular Vesicles and Cancer Therapy: Insights into the Role of Oxidative Stress. Antioxidants 2022, 11, 1194. [Google Scholar] [CrossRef] [PubMed]
- Miceli, M.; Roma, E.; Rosa, P.; Feroci, M.; Loreto, M.A.; Tofani, D.; Gasperi, T. Synthesis of Benzofuran-2-One Derivatives and Evaluation of Their Antioxidant Capacity by Comparing DPPH Assay and Cyclic Voltammetry. Molecules 2018, 23, 710. [Google Scholar] [CrossRef] [Green Version]
- Salvianti, F.; Gelmini, S.; Costanza, F.; Mancini, I.; Sonnati, G.; Simi, L.; Pazzagli, M.; Pinzani, P. The pre-analytical phase of the liquid biopsy. New Biotechnol. 2020, 55, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Piazza, A.; Rosa, P.; Ricciardi, L.; Mangraviti, A.; Pacini, L.; Calogero, A.; Raco, A.; Miscusi, M. Circulating Exosomal-DNA in Glioma Patients: A Quantitative Study and Histopathological Correlations-A Preliminary Study. Brain Sci. 2022, 12, 500. [Google Scholar] [CrossRef] [PubMed]
- Del Bene, M.; Osti, D.; Faletti, S.; Beznoussenko, G.V.; DiMeco, F.; Pelicci, G. Extracellular vesicles: The key for precision medicine in glioblastoma. Neuro Oncol. 2022, 24, 184–196. [Google Scholar] [CrossRef]
- Sabbagh, Q.; Andre-Gregoire, G.; Guevel, L.; Gavard, J. Vesiclemia: Counting on extracellular vesicles for glioblastoma patients. Oncogene 2020, 39, 6043–6052. [Google Scholar] [CrossRef]
- Maire, C.L.; Fuh, M.M.; Kaulich, K.; Fita, K.D.; Stevic, I.; Heiland, D.H.; Welsh, J.A.; Jones, J.C.; Gorgens, A.; Ricklefs, T.; et al. Genome-wide methylation profiling of glioblastoma cell-derived extracellular vesicle DNA allows tumor classification. Neuro Oncol. 2021, 23, 1087–1099. [Google Scholar] [CrossRef]
- Santiago-Dieppa, D.R.; Steinberg, J.; Gonda, D.; Cheung, V.J.; Carter, B.S.; Chen, C.C. Extracellular vesicles as a platform for ‘liquid biopsy’ in glioblastoma patients. Expert Rev. Mol. Diagn. 2014, 14, 819–825. [Google Scholar] [CrossRef] [Green Version]
- Fernando, M.R.; Jiang, C.; Krzyzanowski, G.D.; Ryan, W.L. New evidence that a large proportion of human blood plasma cell-free DNA is localized in exosomes. PLoS ONE 2017, 12, e0183915. [Google Scholar] [CrossRef] [Green Version]
- Torrisi, F.; Alberghina, C.; D’Aprile, S.; Pavone, A.M.; Longhitano, L.; Giallongo, S.; Tibullo, D.; di Rosa, M.; Zappala, A.; Cammarata, F.P.; et al. The Hallmarks of Glioblastoma: Heterogeneity, Intercellular Crosstalk and Molecular Signature of Invasiveness and Progression. Biomedicines 2022, 10, 806. [Google Scholar] [CrossRef]
- Patel, A.P.; Tirosh, I.; Trombetta, J.J.; Shalek, A.K.; Gillespie, S.M.; Wakimoto, H.; Cahill, D.P.; Nahed, B.V.; Curry, W.T.; Martuza, R.L.; et al. Single-cell RNA-seq highlights intratumoral heterogeneity in primary glioblastoma. Science 2014, 344, 1396–1401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Romero, N.; Carrion-Navarro, J.; Esteban-Rubio, S.; Lazaro-Ibanez, E.; Peris-Celda, M.; Alonso, M.M.; Guzman-De-Villoria, J.; Fernandez-Carballal, C.; de Mendivil, A.O.; Garcia-Duque, S.; et al. DNA sequences within glioma-derived extracellular vesicles can cross the intact blood-brain barrier and be detected in peripheral blood of patients. Oncotarget 2017, 8, 1416–1428. [Google Scholar] [CrossRef] [Green Version]
- Degli Esposti, C.; Iadarola, B.; Maestri, S.; Beltrami, C.; Lavezzari, D.; Morini, M.; De Marco, P.; Erminio, G.; Garaventa, A.; Zara, F.; et al. Exosomes from Plasma of Neuroblastoma Patients Contain Doublestranded DNA Reflecting the Mutational Status of Parental Tumor Cells. Int. J. Mol. Sci. 2021, 22, 3667. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Parsons, D.W.; Jin, G.; McLendon, R.; Rasheed, B.A.; Yuan, W.; Kos, I.; Batinic-Haberle, I.; Jones, S.; Riggins, G.J.; et al. IDH1 and IDH2 mutations in gliomas. N. Engl. J. Med. 2009, 360, 765–773. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, X.; Yan, X.; Sun, M.; Fan, Y.; Huang, Y. Significance of TERT and ATRX mutations in glioma. Oncol. Lett. 2019, 17, 95–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marques, C.; Unterkircher, T.; Kroon, P.; Oldrini, B.; Izzo, A.; Dramaretska, Y.; Ferrarese, R.; Kling, E.; Schnell, O.; Nelander, S.; et al. NF1 regulates mesenchymal glioblastoma plasticity and aggressiveness through the AP-1 transcription factor FOSL1. eLife 2021, 10, e64846. [Google Scholar] [CrossRef] [PubMed]
- Reynes, G.; Vila, V.; Fleitas, T.; Reganon, E.; Font de Mora, J.; Jorda, M.; Martinez-Sales, V. Circulating endothelial cells and procoagulant microparticles in patients with glioblastoma: Prognostic value. PLoS ONE 2013, 8, e69034. [Google Scholar] [CrossRef] [PubMed]
- Zeng, A.; Wei, Z.; Rabinovsky, R.; Jun, H.J.; El Fatimy, R.; Deforzh, E.; Arora, R.; Yao, Y.; Yao, S.; Yan, W.; et al. Glioblastoma-Derived Extracellular Vesicles Facilitate Transformation of Astrocytes via Reprogramming Oncogenic Metabolism. iScience 2020, 23, 101420. [Google Scholar] [CrossRef] [PubMed]
- Shao, H.; Chung, J.; Balaj, L.; Charest, A.; Bigner, D.D.; Carter, B.S.; Hochberg, F.H.; Breakefield, X.O.; Weissleder, R.; Lee, H. Protein typing of circulating microvesicles allows real-time monitoring of glioblastoma therapy. Nat. Med. 2012, 18, 1835–1840. [Google Scholar] [CrossRef] [PubMed]
- Tirro, E.; Massimino, M.; Broggi, G.; Romano, C.; Minasi, S.; Gianno, F.; Antonelli, M.; Motta, G.; Certo, F.; Altieri, R.; et al. A Custom DNA-Based NGS Panel for the Molecular Characterization of Patients With Diffuse Gliomas: Diagnostic and Therapeutic Applications. Front. Oncol. 2022, 12, 861078. [Google Scholar] [CrossRef] [PubMed]
- Broggi, G.; Piombino, E.; Altieri, R.; Romano, C.; Certo, F.; Barbagallo, G.M.V.; Vigneri, P.; Condorelli, D.; Colarossi, L.; Colarossi, C.; et al. Glioblastoma, IDH-Wild Type With FGFR3-TACC3 Fusion: When Morphology May Reliably Predict the Molecular Profile of a Tumor. A Case Report and Literature Review. Front. Neurol. 2022, 13, 823015. [Google Scholar] [CrossRef] [PubMed]
- Barger, C.J.; Suwala, A.K.; Soczek, K.M.; Wang, A.S.; Kim, M.Y.; Hong, C.; Doudna, J.A.; Chang, S.M.; Phillips, J.J.; Solomon, D.A.; et al. Conserved features of TERT promoter duplications reveal an activation mechanism that mimics hotspot mutations in cancer. Nat. Commun. 2022, 13, 5430. [Google Scholar] [CrossRef] [PubMed]
- Killela, P.J.; Reitman, Z.J.; Jiao, Y.; Bettegowda, C.; Agrawal, N.; Diaz, L.A., Jr.; Friedman, A.H.; Friedman, H.; Gallia, G.L.; Giovanella, B.C.; et al. TERT promoter mutations occur frequently in gliomas and a subset of tumors derived from cells with low rates of self-renewal. Proc. Natl. Acad. Sci. USA 2013, 110, 6021–6026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olympios, N.; Gilard, V.; Marguet, F.; Clatot, F.; Di Fiore, F.; Fontanilles, M. TERT Promoter Alterations in Glioblastoma: A Systematic Review. Cancers 2021, 13, 1147. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Characteristics | Patients (n = 26) |
|---|---|
| Median age, years | 60.1 (±16) |
| Gender (%) | |
| Male | 57.7 |
| Female | 42.3 |
| Tumor location (%) | |
| Frontal lobe | 30.8 |
| Right temporal lobe | 11.5 |
| Left temporal lobe | 19.2 |
| Temporo-parietal lobe | 3.9 |
| Temporo-insular | 3.9 |
| Occipital | 11.5 |
| Frontoparietal lobe | 7.7 |
| Parietal | 11.5 |
| Histology (WHO 2016) (%) | |
| IV | 77.0 |
| III | 11.5 |
| II | 11.5 |
| I | 0 |
| Surgical resection (%) | |
| Gross total | 46.2 |
| Near total | 42.3 |
| Partial | 0 |
| Biopsy | 11.5 |
| Median tumor volume, cm3 (±SD) | |
| Total Flair | 63.4 (±56.3) |
| Non-enhancing (t1 Post-contrast) | 6.6 (±16.8) |
| Enhancing (t1 Post-contrast) | 16.1 (±18.7) |
| Enhancing + Non-enhancing (t1 Post- | 23.4 (±30.6) |
| contrast) | |
| Karnofsky Performance Status (KPS) (%) preoperative | |
| 90\100 | 76.9 |
| 80\70 | 19.2 |
| <60 | 3.9 |
| Karnofsky Performance Status (KPS) (%) preoperative | |
| 90\100 | 84.6 |
| 80\70 | 9.2 |
| <60 | 11.5 |
| Progression (%) | |
| Yes | 80.8 |
| No | 19.2 |
| Free survival, months | 13.23 (±17.9) |
| Survival, months | 15.92 (±16.11) |
| Immunohistochemical Characteristics | Patients (n = 26) |
|---|---|
| GFAP | 23 |
| OLIG2 | 15 |
| ATRX | 2 |
| EGFR | 7 |
| IDH1 | 3 |
| p53 | 19 |
| ki67 (mean % all patients) | 28 |
| mitosis (mean over 10 HPF all patients) | 22.5 |
| Characteristics | Patients (n = 10) |
|---|---|
| Median age, years | 64.6 (±16.06) |
| Gender (%) | |
| Male | 70 |
| Female | 30 |
| Tumor location, (%) | |
| Frontal lobe | 20 |
| Right temporal lobe | 20 |
| Left temporal lobe | 20 |
| Temporo-parietal lobe | 10 |
| Occipital | 20 |
| Frontoparietal lobe | 10 |
| Surgical resection, (%) | |
| Gross total | 30 |
| Near total | 40 |
| Partial | 10 |
| Biopsy | 20 |
| Median tumor volume, cm3 (±SD) | |
| Total Flair | 85.3 (±5.55) |
| Non-enhancing (t1 Post-contrast) | 12.26 (±25.66) |
| Enhancing (t1 Post-contrast) | 17.26 (±25.66) |
| Enhancing + Non-enhancing (t1 Post- | 31.32 (±34.67) |
| contrast) | |
| Karnofsky Performance Status (KPS), (%) Preoperative | |
| 90\100 | 70 |
| 80\70 | 20 |
| <60 | 10 |
| Karnofsky Performance Status (KPS), (%) Preoperative | |
| 90\100 | 70 |
| 80\70 | 20 |
| <60 | 10 |
| Progression (%) | |
| Yes | 90 |
| No | 10 |
| Free survival, months | 8.2 (±11.37) |
| Survival, months | 10.5 (±8.14) |
| Gene | Nucleotide Change | Amino Acid Change |
|---|---|---|
| NF1 | c.233delA c.1466A>G c.1658A>G c.2027delC c.2568C>G c.3033delA c.3859T>C c.2297T>C | p.Asn78fs p.Tyr489Cys p.His553Arg p.Pro678fs p.Ser856Arg p.Thr1013fs p.Phe1287Leu p.Ile766Thr |
| IDH1 | c.395G>A | p.Arg132His |
| TP53 | c.700T>A c.1146delA c.841G>T | p.Tyr234Asn p.Lys382fs p.Asp281Tyr |
| ATRX | c.1074delA c.2658_2659delGA | p.Lys358fs p.Glu886fs |
| TERT | c.336delC | p.Glu113fs |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosa, P.; De Falco, E.; Pacini, L.; Piazza, A.; Ciracì, P.; Ricciardi, L.; Fiorentino, F.; Trungu, S.; Miscusi, M.; Raco, A.; et al. Next-Generation Sequencing Comparative Analysis of DNA Mutations between Blood-Derived Extracellular Vesicles and Matched Cancer Tissue in Patients with Grade 4 Glioblastoma. Biomedicines 2022, 10, 2590. https://doi.org/10.3390/biomedicines10102590
Rosa P, De Falco E, Pacini L, Piazza A, Ciracì P, Ricciardi L, Fiorentino F, Trungu S, Miscusi M, Raco A, et al. Next-Generation Sequencing Comparative Analysis of DNA Mutations between Blood-Derived Extracellular Vesicles and Matched Cancer Tissue in Patients with Grade 4 Glioblastoma. Biomedicines. 2022; 10(10):2590. https://doi.org/10.3390/biomedicines10102590
Chicago/Turabian StyleRosa, Paolo, Elena De Falco, Luca Pacini, Amedeo Piazza, Paolo Ciracì, Luca Ricciardi, Francesco Fiorentino, Sokol Trungu, Massimo Miscusi, Antonino Raco, and et al. 2022. "Next-Generation Sequencing Comparative Analysis of DNA Mutations between Blood-Derived Extracellular Vesicles and Matched Cancer Tissue in Patients with Grade 4 Glioblastoma" Biomedicines 10, no. 10: 2590. https://doi.org/10.3390/biomedicines10102590
APA StyleRosa, P., De Falco, E., Pacini, L., Piazza, A., Ciracì, P., Ricciardi, L., Fiorentino, F., Trungu, S., Miscusi, M., Raco, A., & Calogero, A. (2022). Next-Generation Sequencing Comparative Analysis of DNA Mutations between Blood-Derived Extracellular Vesicles and Matched Cancer Tissue in Patients with Grade 4 Glioblastoma. Biomedicines, 10(10), 2590. https://doi.org/10.3390/biomedicines10102590