
Role of Extracellular Vesicles in Thyroid Physiology and Diseases: Implications for Diagnosis and Treatment

Abstract
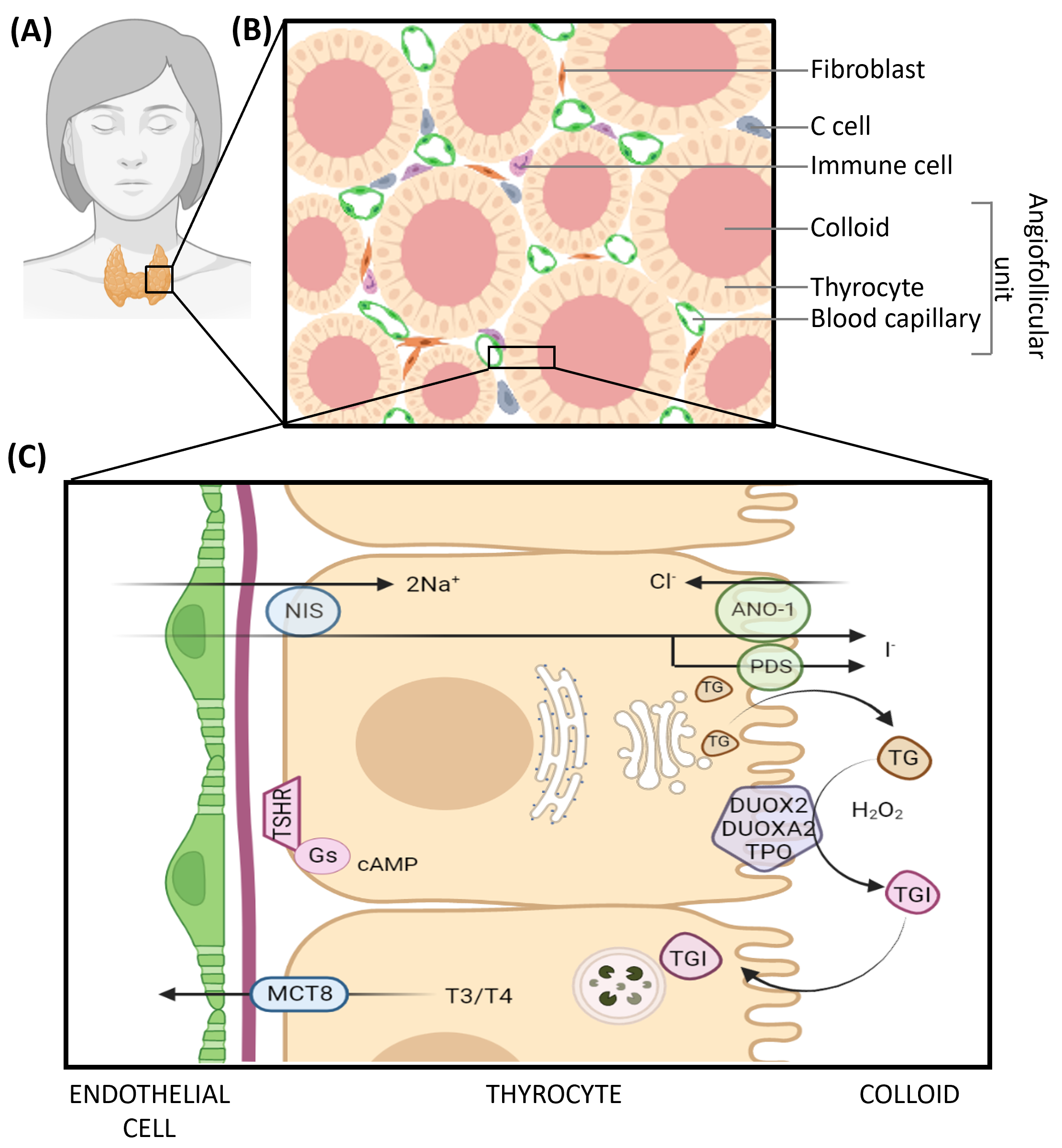
1. Thyroid Physiology and Thyroid Diseases
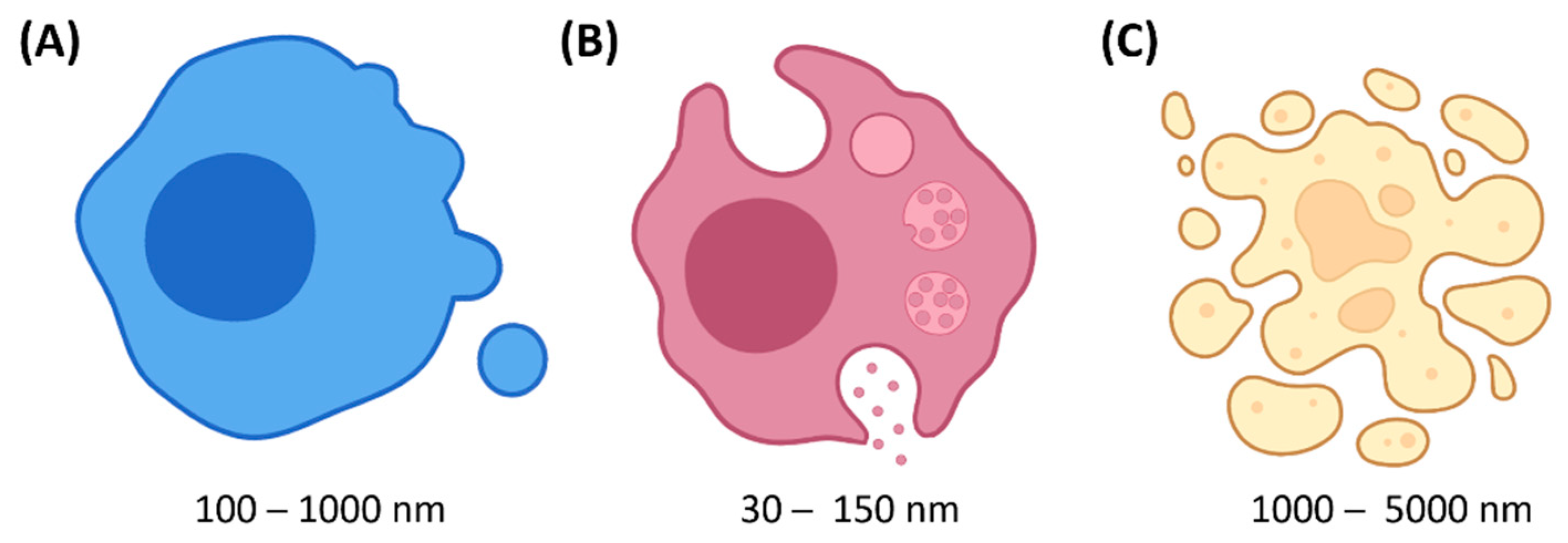
2. Extracellular Vesicles (EVs)
2.1. Definition & Biogenesis
2.2. Function
2.3. Therapeutic and Diagnostic Potentials
3. EVs in Thyroid Physiology
4. EVs in TC
4.1. Role
4.2. TC-Derived EV-Mediated Role in Thyroid Cell Transformation
4.3. TC-Derived EV-Mediated Communication with the Tumor Microenvironment
4.4. Link with BRAF and Resistance to Treatment
4.5. Diagnosis
4.6. Treatment
5. EVs in AITD
5.1. Role
5.2. Diagnosis
5.3. Treatment
6. Concerns & Further Directions
6.1. EVs Production & Release
6.2. EVs Function
6.3. Diagnosis
6.4. Treatment
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Colin, I.M.; Denef, J.-F.; Lengelé, B.; Many, M.-C.; Gérard, A.-C. Recent insights into the cell biology of thyroid angiofollicular units. Endocr. Rev. 2013, 34, 209–238. [Google Scholar] [CrossRef]
- Durante, C.; Grani, G.; Lamartina, L.; Filetti, S.; Mandel, S.J.; Cooper, D.S. The diagnosis and management of thyroid nodules: A review. JAMA 2018, 319, 914–924. [Google Scholar] [CrossRef] [PubMed]
- Fagin, J.A.; Wells, S.A., Jr. Biologic and clinical perspectives on thyroid cancer. N. Engl. J. Med. 2016, 375, 1054–1067. [Google Scholar] [CrossRef]
- Cabanillas, M.E.; McFadden, D.G.; Durante, C. Thyroid cancer. Lancet. 2016, 388, 2783–2795. [Google Scholar] [CrossRef]
- Antonelli, A.; Ferrari, S.M.; Corrado, A.; Domenicantonio, A.D.; Fallahi, P. Autoimmune thyroid disorders. Autoimmun. Rev. 2015, 14, 174–180. [Google Scholar] [CrossRef]
- Colombo, M.; Raposo, G.; Théry, C. Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu. Rev. Cell Dev. Biol. 2014, 30, 255–289. [Google Scholar] [CrossRef] [PubMed]
- Yáñez-Mó, M.; Siljander, P.R.M.; Andreu, Z.; Bedina Zavec, A.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [PubMed]
- Van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef]
- Raposo, G.; Nijman, H.W.; Stoorvogel, W.; Liejendekker, R.; Harding, C.V.; Melief, C.J.; Geuze, H.J. B lymphocytes secrete antigen-presenting vesicles. J. Exp. Med. 1996, 183, 1161–1172. [Google Scholar] [CrossRef]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [PubMed]
- McAndrews, K.M.; Kalluri, R. Mechanisms associated with biogenesis of exosomes in cancer. Mol. Cancer 2019, 18, 52. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, A.; Costa-Silva, B.; Shen, T.-L.; Rodrigues, G.; Hashimoto, A.; Mark, M.T.; Molina, H.; Kohsaka, S.; Di Giannatale, A.; Ceder, S.; et al. Tumour exosome integrins determine organotropic metastasis. Nature 2015, 527, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Buzas, E.I. The roles of extracellular vesicles in the immune system. Nat. Rev. Immunol. 2022, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Whiteside, T.L. Extracellular vesicles isolation and their biomarker potential: Are we ready for testing? Ann. Transl. Med. 2017, 5, 54. [Google Scholar] [CrossRef]
- Jafarinia, M.; Alsahebfosoul, F.; Salehi, H.; Eskandari, N.; Ganjalikhani-Hakemi, M. Mesenchymal stem cell-derived extracellular vesicles: A novel cell-free therapy. Immunol. Investig. 2020, 49, 758–780. [Google Scholar] [CrossRef] [PubMed]
- Nunez, E.A.; Wallis, J.; Gershon, M.D. Secretory processes in follicular cells of the bat thyroid. III. The occurrence of extracellular vesicles and colloid droplets during arousal from hibernation. Am. J. Anat. 1974, 141, 179–201. [Google Scholar] [CrossRef] [PubMed]
- Tachiwaki, O.; Wollman, S.H. Shedding of dense cell fragments into the follicular lumen early in involution of the hyperplastic thyroid gland. Lab. Investig. 1982, 47, 91–98. [Google Scholar]
- Nilsson, M.; Ofverholm, T.; Ericson, L. In vivo shedding of apical plasma membrane in the thyroid follicle cells of the mouse. Cell Tissue Res. 1984, 236, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, M.; Fau-Jörtsö, E.M.J.; Fau-Smeds, S.J.E.; Fau-Ericson, L.E.S.S.; Ericson, L.E. Plasma membrane shedding and colloid vacuoles in hyperactive human thyroid tissue. Virchows Arch. B 1988, 56, 85–94. [Google Scholar] [CrossRef]
- Vlasov, P.; Doi, S.Q.; Sellitti, D.F. FRTL-5 rat thyroid cells release thyroglobulin sequestered in exosomes: A possible novel mechanism for thyroglobulin processing in the thyroid. J. Thyroid Res. 2016, 2016, 9276402. [Google Scholar] [CrossRef] [PubMed]
- Rosa, I.; Ibba-Manneschi, L.; Guasti, D.; Perigli, G.; Faussone-Pellegrini, M.; Manetti, M. Morphologic evidence of telocytes in human thyroid stromal tissue. J. Cell. Mol. Med. 2022, 26, 2477–2481. [Google Scholar] [CrossRef]
- Delcorte, O.; Spourquet, C.; Lemoine, P.; Degosserie, J.; Van Der Smissen, P.; Dauguet, N.; Loriot, A.; Knauf, J.A.; Gatto, L.; Marbaix, E.; et al. BRAFV600E induction in thyrocytes triggers important changes in the miRNAs content and the populations of extracellular vesicles released in thyroid tumor microenvironment. Biomedicines 2022, 10, 755. [Google Scholar] [CrossRef]
- Degosserie, J.; Heymans, C.; Spourquet, C.; Halbout, M.; D’Auria, L.; Van Der Smissen, P.; Vertommen, D.; Courtoy, P.J.; Tyteca, D.; Pierreux, C.E. Extracellular vesicles from endothelial progenitor cells promote thyroid follicle formation. J. Extracell. Vesicles 2018, 7, 1487250. [Google Scholar] [CrossRef]
- Hick, A.-C.; Delmarcelle, A.-S.; Bouquet, M.; Klotz, S.; Copetti, T.; Forez, C.; Van Der Smissen, P.; Sonveaux, P.; Collet, J.-F.; Feron, O.; et al. Reciprocal epithelial: Endothelial paracrine interactions during thyroid development govern follicular organization and C-cells differentiation. Dev. Biol. 2013, 381, 227–240. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Liu, W.; Wang, Z.; Wang, C.; Ai, Z. Exosomal ANXA1 derived from thyroid cancer cells is associated with malignant transformation of human thyroid follicular epithelial cells by promoting cell proliferation. Int. J. Oncol. 2021, 59, 104. [Google Scholar] [CrossRef]
- Surman, M.; Kędracka-Krok, S.; Wilczak, M.; Rybczyński, P.; Jankowska, U.; Przybyło, M. Comparative proteomic profiling of ectosomes derived from thyroid carcinoma and normal thyroid cells uncovers multiple proteins with functional implications in cancer. Cells 2018, 11, 1184. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; He, Y.; Liu, X.; Luo, F.; Jiang, Y.; Xiang, M.; Zhao, R. Cancer stem cell-like cells-derived exosomal CDKN2B-AS1 stabilizes CDKN2B to promote the growth and metastasis of thyroid cancer via TGF-β1/Smad2/3 signaling. Exp. Cell Res. 2022, 419, 113268. [Google Scholar] [CrossRef] [PubMed]
- Luo, D.; Zhan, S.; Xia, W.; Huang, L.; Ge, W.; Wang, T. Proteomics study of serum exosomes from papillary thyroid cancer patients. Endocr. Relat. Cancer 2018, 25, 879–891. [Google Scholar] [CrossRef]
- Grzanka, M.; Stachurska-Skrodzka, A.; Adamiok-Ostrowska, A.; Gajda, E.; Czarnocka, B. Extracellular vesicles as signal carriers in malignant thyroid tumors? Int. J. Mol. Sci. 2022, 23, 3262. [Google Scholar] [CrossRef]
- Lee, J.C.; Zhao, J.-T.; Gundara, J.; Serpell, J.; Bach, L.A.; Sidhu, S. Papillary thyroid cancer–derived exosomes contain miRNA-146b and miRNA-222. J. Surg. Res. 2015, 196, 39–48. [Google Scholar] [CrossRef]
- Hardin, H.; Helein, H.; Meyer, K.; Robertson, S.; Zhang, R.; Zhong, W.; Lloyd, R.V. Thyroid cancer stem-like cell exosomes: Regulation of EMT via transfer of lncRNAs. Lab. Investig. 2018, 98, 1133–1142. [Google Scholar] [CrossRef] [PubMed]
- Wen, D.; Liu, W.-L.; Lu, Z.-W.; Cao, Y.-M.; Ji, Q.-H.; Wei, W.-J. SNHG9, a papillary thyroid cancer cell exo-some-enriched lncRNA, inhibits Cell autophagy and promotes cell apoptosis of normal thyroid epithelial cell Nthy-ori-3 through YBOX3/P21 pathway. Front. Oncol. 2021, 11, 647034. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Wang, S.; Zhang, M.; Li, Y.; Liu, Q.; Sun, N.; Zhang, X.; Liu, Y.; Zhang, J.; He, L.; et al. EV PD-L1 is correlated with clinical features and contributes to T cell suppression in pediatric thyroid cancer. J. Clin. Endocrinol. Metab. 2020, 105, e2970–e2981. [Google Scholar] [CrossRef] [PubMed]
- Bravo-Miana, R.D.C.; Vedova, A.B.D.; De Paul, A.L.; Remedi, M.M.; Guantay, M.L.; Gilardoni, M.B.; Pellizas, C.G.; Donadio, A.C. Thyroid tumor cells-fibroblasts crosstalk: Role of extracellular vesicles. Endocr. Connect. 2020, 9, 506–518. [Google Scholar] [CrossRef] [PubMed]
- Bravo-Miana, R.D.C.; Soler, M.F.; Ceschin, D.G.; Royo, F.; Negretti-Borga, D.M.; Azkargorta, M.; Elortza, F.; Montesinos, M.D.M.; Pellizas, C.G.; Falcón-Pérez, J.M.; et al. Extracellular vesicles from thyroid cancer harbor a functional machinery involved in extracellular matrix remodeling. Eur. J. Cell Biol. 2022, 101, 151254. [Google Scholar] [CrossRef]
- Wang, Y.; Cen, A.; Yang, Y.; Ye, H.; Li, J.; Liu, S.; Zhao, L. miR-181a, delivered by hypoxic PTC-secreted exosomes, inhibits DACT2 by downregulating MLL3, leading to YAP-VEGF-mediated angiogenesis. Mol. Ther. Nucleic Acids 2021, 24, 610–621. [Google Scholar] [CrossRef]
- Wu, F.L.F.; Lin, X.; Xu, F.; Cui, R.; Zhong, J.; Zhu, T.; Shan, S.; Liao, X.; Yuan, L.; Mo, Z. Exosomes increased angiogenesis in papillary thyroid cancer microenvironment. Endocr. Relat. Cancer 2019, 26, 525–538. [Google Scholar] [CrossRef]
- Yin, G.; Kong, W.; Zheng, S.; Shan, Y.; Zhang, J.; Ying, R.; Wu, H. Exosomal miR-130a-3p promotes the progression of differentiated thyroid cancer by targeting insulin-like growth factor 1. Oncol. Lett. 2021, 21, 283. [Google Scholar] [CrossRef]
- Vella, L.J.; Behren, A.; Coleman, B.; Greening, D.W.; Hill, A.F.; Cebon, J. Intercellular resistance to BRAF inhibition can be mediated by extracellular vesicle–associated PDGFRβ. Neoplasia 2017, 19, 932–940. [Google Scholar] [CrossRef]
- Samsonov, B.V.; Shtam, T.; RadzhabovA, Z.; Vasilyev, D.; Tsyrlina, E.; Titov, S.; Ivanov, M.; Berstein, L.; Filatov, M.; Kolesnikov, N.; et al. Plasma exosomal miR-21 and miR-181a differentiates follicular from papillary thyroid cancer. Tumour Biol. 2016, 37, 12011–12021. [Google Scholar] [CrossRef]
- Maggisano, V.; Capriglione, F.; Verrienti, A.; Celano, M.; Gagliardi, A.; Bulotta, S.; Sponziello, M.; Mio, C.; Pecce, V.; Durante, C.; et al. Identification of exosomal microRNAs and their targets in papillary thyroid cancer cells. Biomedicines 2022, 10, 961. [Google Scholar] [CrossRef] [PubMed]
- Xing, M.; Alzahrani, A.S.; Carson, K.A.; Viola, D.; Elisei, R.; Bendlova, B.; Yip, L.; Mian, C.; Vianello, F.; Tuttle, R.M.; et al. Association between BRAF V600E mutation and mortality in patients with papillary thyroid cancer. JAMA 2013, 309, 1493–1501. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, K.; Saji, M.; Lazaroff, S.M.; Palmer, A.F.; Ringel, M.D.; Paulaitis, M.E. Analysis of exosome release as a cellular response to MAPK pathway inhibition. Langmuir 2015, 31, 5440–5448. [Google Scholar] [CrossRef] [PubMed]
- Taral, R.L.; Cheng, L.; Berglind, O.E.; Bagge, R.O.; Muralidharan, S.V.; Robyn, A.S.; Lässer, C.; Yong, S.G.; Andrew, F.H.; Jonas, A.N.; et al. BRAFV600 inhibition alters the microRNA cargo in the vesicular secretome of malignant melanoma cells. Proc. Natl. Acad. Sci. USA 2017, 114, E5930–E5939. [Google Scholar]
- Kasumova, G.G.; Shi, A.; Cintolo-Gonzalez, J.A.; Chein, I.; Frederick, D.T.; Alpatov, R.; Michaud, W.A.; Plana, D.; Panka, D.J.; Corcoran, R.B.; et al. Abstract A35: BRAF inhibition increases exosomal PD-L1 protein expression in melanoma. Cancer Immunol. Res. 2018, 6, A35. [Google Scholar] [CrossRef]
- Jensen, C.B.; Saucke, M.C.; Francis, D.O.; Voils, C.I.; Pitt, S.C. From overdiagnosis to overtreatment of low-risk thyroid cancer: A thematic analysis of attitudes and beliefs of endocrinologists, surgeons, and patients. Thyroid 2020, 30, 696–703. [Google Scholar] [CrossRef]
- Zabegina, L.; Nazarova, I.; Knyazeva, M.; Nikiforova, N.; Slyusarenko, M.; Titov, S.; Vasilyev, D.; Sleptsov, I.; Malek, A. MiRNA let-7 from TPO(+) extracellular vesicles is a potential marker for a differential diagnosis of follicular thyroid nodules. Cells 2020, 9, 1917. [Google Scholar] [CrossRef]
- Huang, T.Y.; Wang, C.-Y.; Chen, K.-Y.; Huang, L.-T. Urinary exosomal thyroglobulin in thyroid cancer patients with post-ablative therapy: A new biomarker in thyroid cancer. Front. Endocrinol. 2020, 11, 382. [Google Scholar] [CrossRef]
- Bavisotto, C.C.; Cipolla, C.; Graceffa, G.; Barone, R.; Bucchieri, F.; Bulone, D.; Cabibi, D.; Campanella, C.; Gammazza, A.M.; Pitruzzella, A.; et al. Immunomorphological pattern of molecular chaperones in normal and pathological thyroid tissues and circulating exosomes: Potential use in clinics. Int. J. Mol. Sci. 2019, 20, 4496. [Google Scholar] [CrossRef]
- Wen, Q.; Wang, Y.; Li, X.; Jin, X.; Wang, G. Decreased serum exosomal miR-29a expression and its clinical significance in papillary thyroid carcinoma. J. Clin. Lab. Anal. 2020, 35, e23560. [Google Scholar] [CrossRef]
- Wang, Z.; Lv, J.; Zou, X.; Huang, Z.; Zhang, H.; Liu, Q.; Jiang, L.; Zhou, X.; Zhu, W. A three plasma microRNA signature for papillary thyroid carcinoma diagnosis in Chinese patients. Gene 2019, 693, 37–45. [Google Scholar] [CrossRef]
- Liang, M.; Yu, S.; Tang, S.; Bai, L.; Cheng, J.; Gu, Y.; Li, S.; Zheng, X.; Duan, L.; Wang, L.; et al. A panel of plasma exosomal miRNAs as potential biomarkers for differential diagnosis of thyroid nodules. Front. Genet. 2020, 11, 449. [Google Scholar] [CrossRef] [PubMed]
- Xin, Y.; Meng, K.; Guo, H.; Chen, B.; Zheng, C.; Shou, X.; Yu, K. Exosomal hsa-miR-129-2 and hsa-miR-889 from a 6-microRNA signature might be a potential biomarker for predicting the prognosis of papillary thyroid carcinoma. Comb. Chem. High Throughput Screen. 2022, 25, 819–830. [Google Scholar] [PubMed]
- Delcorte, O.; Craps, J.; Mahibullah, S.; Spourquet, C.; D’Auria, L.; Van Der Smissen, P.; Dessy, C.; Marbaix, E.; Mourad, M.; E Pierreux, C. Two miRNAs enriched in plasma extracellular vesicles are potential biomarkers for thyroid cancer. Endocr. Relat. Cancer 2022, 29, 389–401. [Google Scholar] [CrossRef] [PubMed]
- Capriglione, F.V.A.; Celano, M.; Maggisano, V.; Sponziello, M.; Pecce, V.; Gagliardi, A.; Giacomelli, L.; Aceti, V.; Durante, C.; Bulotta, S.; et al. Analysis of serum microRNA in exosomal vehicles of papillary thyroid cancer. Endocrine 2021, 75, 185–193. [Google Scholar] [CrossRef]
- Jiang, K.; Li, G.; Chen, W.; Song, L.; Wei, T.; Li, Z.; Gong, R.; Lei, J.; Shi, H.; Zhu, J. Plasma exosomal miR-146b-5p and miR-222-3p are potential biomarkers for lymph node metastasis in papillary thyroid carcinomas. OncoTargets Ther. 2020, 13, 1311–1319. [Google Scholar] [CrossRef]
- Chen, W.; Li, G.; Li, Z.; Zhu, J.; Wei, T.; Lei, J. Evaluation of plasma exosomal miRNAs as potential diagnostic biomarkers of lymph node metastasis in papillary thyroid carcinoma. Endocrine 2021, 75, 846–855. [Google Scholar] [CrossRef]
- Yao, X.; Zhang, Q. Function and clinical significance of circular RNAs in thyroid cancer. Front. Mol. Biosci. 2022, 9, 925389. [Google Scholar] [CrossRef]
- Yang, C.; Wei, Y.; Yu, L.; Xiao, Y. Identification of altered circular RNA expression in serum exosomes from patients with papillary thyroid carcinoma by high-throughput sequencing. Med. Sci. Monit. 2019, 25, 2785–2791. [Google Scholar] [CrossRef]
- Pan, Q.; Zhao, J.; Li, M.; Liu, X.; Xu, Y.; Li, W.; Wu, S.; Su, Z. Exosomal miRNAs are potential diagnostic biomarkers between malignant and benign thyroid nodules based on next-generation sequencing. Carcinogenesis 2020, 41, 18–24. [Google Scholar] [CrossRef]
- Dai, D.; Tan, Y.; Guo, L.; Tang, A.; Zhao, Y. Identification of exosomal miRNA biomarkers for diagnosis of papillary thyroid cancer by small RNA sequencing. Eur. J. Endocrinol. 2020, 182, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, R.; Paudel, S.; Gangadaran, P.; Oh, J.; Oh, E.; Hong, C.; Lee, S.; Chung, H.; Lee, J.; Ahn, B.-C. Extracellular vesicles act as nano-transporters of tyrosine kinase inhibitors to revert iodine avidity in thyroid cancer. Pharmaceutics 2021, 13, 248. [Google Scholar] [CrossRef] [PubMed]
- Son, S.H.; Gangadaran, P.; Ahn, B.-C. A novel strategy of transferring NIS protein to cells using extracellular vesicles leads to increase in iodine uptake and cytotoxicity. Int. J. Nanomed. 2019, 14, 1779–1787. [Google Scholar] [CrossRef]
- Lee, J.H.; Jung, K.-H.; Mina, K.; Lee, K.-H. Extracellular vesicles deliver sodium iodide symporter protein and promote cancer cell radioiodine therapy. Sci. Rep. 2022, 12, 11190. [Google Scholar] [CrossRef] [PubMed]
- Zheng, T.; Zhou, Y.; Xu, X.; Qi, X.; Liu, J.; Pu, Y.; Zhang, S.; Gao, X.; Luo, X.; Li, M.; et al. MiR-30c-5p loss-induced PELI1 accumulation regulates cell proliferation and migration via activating PI3K/AKT pathway in papillary thyroid carcinoma. J. Transl. Med. 2022, 20, 1–16. [Google Scholar] [CrossRef]
- Wang, M.H.; Ye, Y.; Zhang, M.; Zhou, B.R.; Wang, J.N.; Song, Y.N.; Xia, W. Exosome-mediated delivery of SCD-1 siRNA promoted the death of anaplastic thyroid carcinoma cells via regulating ROS level. Clin. Transl. Oncol. 2021, 24, 288–296. [Google Scholar] [CrossRef]
- Ryder, M.; Ghossein, R.A.; Ricarte-Filho, J.C.M.; Knauf, J.A.; Fagin, J.A. Increased density of tumor-associated macro-phages is associated with decreased survival in advanced thyroid cancer. Endocr. Relat. Cancer 2008, 15, 1069–1074. [Google Scholar] [CrossRef]
- Yang, Z.; Wei, X.; Pan, Y.; Xu, J.; Si, Y.; Min, Z.; Yu, B. A new risk factor indicator for papillary thyroid cancer based on immune infiltration. Cell Death Dis. 2021, 12, 51. [Google Scholar] [CrossRef]
- Zhu, L.; Gangadaran, P.; Kalimuthu, S.; Oh, J.M.; Baek, S.H.; Jeong, S.Y.; Lee, S.-W.; Lee, J.; Ahn, B.-C. Novel alternatives to extracellular vesicle-based immunotherapy-exosome mimetics derived from natural killer cells. Artif. Cells Nanomed. Biotechnol. 2018, 46, S166–S179. [Google Scholar] [CrossRef]
- Zhu, L.; Kalimuthu, S.; Oh, J.M.; Gangadaran, P.; Baek, S.H.; Jeong, S.Y.; Lee, S.-W.; Lee, J.; Ahn, B.-C. Enhancement of antitumor potency of extracellular vesicles derived from natural killer cells by IL-15 priming. Biomaterials 2018, 190–191, 38–50. [Google Scholar] [CrossRef]
- Edo, N.; Kawakami, K.; Fujita, Y.; Morita, K.; Uno, K.; Tsukamoto, K.; Onose, H.; Ishikawa, T.; Ito, M. Exosomes ex-pressing thyrotropin receptor attenuate autoantibody-mediated stimulation of cyclic adenosine monophosphate production. Thyroid 2019, 29, 1012–1017. [Google Scholar] [CrossRef] [PubMed]
- Rossi, M.; Taddei, A.R.; Fasciani, I.; Maggio, R.; Giorgi, F. The cell biology of the thyroid-disrupting mechanism of dichlorodiphenyltrichloroethane (DDT). J. Endocrinol. Investig. 2018, 41, 67–73. [Google Scholar] [CrossRef]
- Cui, X.; Liu, Y.; Wang, S.; Zhao, N.; Qin, J.; Li, Y.; Fan, C.; Shan, Z.; Teng, W. Circulating exosomes activate dendritic cells and induce unbalanced CD4+ T Cell differentiation in hashimoto thyroiditis. J. Clin. Endocrinol. Metab. 2019, 104, 4607–4618. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Wang, S.; Zhao, N.; Wang, S.; Wang, Z.; Huang, M.; Liu, Y.; Qin, J.; Shan, Z.; Teng, W.; et al. Thyrocyte-derived exosome-targeted dendritic cells stimulate strong CD4+ T lymphocyte responses. Mol. Cell. Endocrinol. 2020, 506, 110756. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Huang, M.; Wang, S.; Zhao, N.; Huang, T.; Wang, Z.; Qiao, J.; Wang, S.; Shan, Z.; Teng, W.; et al. Circulating exosomes from patients with graves’ disease induce an inflammatory immune response. Endocrinology 2020, 162, bqaa236. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Muñoz, A.; Martinez-Hernandez, R.; Ramos-Leví, A.M.; Serrano-Somavilla, A.; Gonzalez-Amaro, R.; Sánchez-Madrid, F.; De La Fuente, H.; Marazuela, M. Circulating microvesicles regulate treg and Th17 differentiation in human autoimmune thyroid disorders. J. Clin. Endocrinol. Metab. 2015, 100, E1531–E1539. [Google Scholar] [CrossRef]
- Han, J.-S.; Kim, S.; Jin, J.-Q.; Park, N.; Lee, J.-Y.; Kim, H.; Lee, S.-B.; Yang, S.-W.; Lim, D.-J. Tear-derived exosome proteins are increased in patients with thyroid eye disease. Int. J. Mol. Sci. 2021, 22, 1115. [Google Scholar] [CrossRef] [PubMed]
- Hiratsuka, I.; Yamada, H.; Munetsuna, E.; Hashimoto, S.; Itoh, M. Circulating microRNAs in graves’ disease in relation to clinical activity. Thyroid 2016, 26, 1431–1440. [Google Scholar] [CrossRef]
- Mobarrez, F.; Gatica, K.A.; Friberg, I.; Antovic, A.; Pisetsky, D.S.; Jörneskog, G.; Wallén, H.; Abraham-Nordling, M. The expression of microvesicles in the blood of patients with Graves’ disease and its relationship to treatment. Clin. Endocrinol. 2015, 84, 729–735. [Google Scholar] [CrossRef]
- Jia, X.; Zhai, T.; Zhang, J.-A. Circulating exosome involves in the pathogenesis of autoimmune thyroid diseases through immunomodulatory proteins. Front. Immunol. 2021, 12, 730089. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Yuan, T.; Tschannen, M.; Sun, Z.; Jacob, H.; Du, M.; Liang, M.; Dittmar, R.L.; Liu, Y.; Liang, M.; et al. Characterization of human plasma-derived exosomal RNAs by deep sequencing. BMC Genom. 2013, 14, 319. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Wang, W.; Tang, Y.; Wang, D.; Li, L.; Na, M.; Jiang, G.; Li, Q.; Chen, S.; Zhou, J. Microarray profiling and functional analysis of differentially expressed plasma exosomal circular RNAs in Graves’ disease. Biol. Res. 2020, 53, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Chen, Y.; Yin, G.; Xie, Q. Therapeutic prospects of MicroRNAs carried by mesenchymal stem cells-derived extracellular vesicles in autoimmune diseases. Life Sci. 2021, 277, 119458. [Google Scholar] [CrossRef]
- Lamparelli, E.; Ciardulli, M.; Scala, P.; Scognamiglio, M.; Charlier, B.; Di Pietro, P.; Izzo, V.; Vecchione, C.; Maffulli, N.; Della Porta, G. Lipid nano-vesicles for thyroid hormone encapsulation: A comparison between different fabrication technologies, drug loading, and an in vitro delivery to human tendon stem/progenitor cells in 2D and 3D culture. Int. J. Pharm. 2022, 624, 122007. [Google Scholar] [CrossRef]
- Ochieng, J.; Pratap, S.; Khatua, A.K.; Sakwe, A. Anchorage-independent growth of breast carcinoma cells is mediated by serum exosomes. Exp. Cell Res. 2009, 315, 1875–1888. [Google Scholar] [CrossRef] [PubMed]
- Saiselet, M.; Floor, S.; Tarabichi, M.; Dom, G.; Hébrant, A.; van Staveren, W.C.G.; Maenhaut, C. Thyroid cancer cell lines: An overview. Front. Endocrinol. 2012, 3, 133. [Google Scholar] [CrossRef]
- Foster, H.; Wade, M.; England, J.; Greenman, J.; Green, V. Isolation and characterisation of graves’ disease-specific extracellular vesicles from tissue maintained on a bespoke microfluidic device. Organs-A-Chip 2021, 3, 100011. [Google Scholar] [CrossRef]
- Khoruzhenko, A.; Miot, F.; Massart, C.; Van Sande, J.; Dumont, J.-E.; Beauwens, R.; Boom, A. Functional model of rat thyroid follicles cultured in matrigel. Endocr. Connect. 2021, 10, 570–578. [Google Scholar] [CrossRef]
- Antonica, F.; Kasprzyk, D.F.; Opitz, R.; Iacovino, M.; Liao, X.H.; Dumitrescu, A.M.; Refetoff, S.; Peremans, K.; Manto, M.; Kyba, M.; et al. Generation of functional thyroid from embryonic stem cells. Nature 2012, 491, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Van der Vaart, J.; Bosmans, L.; Sijbesma, S.F.; Knoops, K.; van de Wetering, W.J.; Otten, H.G.; Begthel, H.; Rinkes, I.H.M.B.; Korving, J.; Lentjes, E.G.W.M.; et al. Adult mouse and human organoids derived from thyroid follicular cells and modeling of Graves’ hyperthyroidism. Proc. Natl. Acad. Sci. USA 2021, 118, e2117017118. [Google Scholar] [CrossRef] [PubMed]
- Debbi, L.; Guo, S.; Safina, D.; Levenberg, S. Boosting extracellular vesicle secretion. Biotechnol. Adv. 2022, 59, 107983. [Google Scholar] [CrossRef]
- Lucien, F.; Leong, H. S The role of extracellular vesicles in cancer microenvironment and metastasis: Myths and challenges. Biochem. Soc. Trans. 2019, 47, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Piffoux, M.; Volatron, J.; Silva, A.K.A.; Gazeau, F. Thinking quantitatively of RNA-based information transfer via extracellular vesicles: Lessons to learn for the design of RNA-loaded EVs. Pharmaceutics 2021, 13, 1931. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Han, Y. The overdiagnosis of thyroid micropapillary carcinoma: The rising incidence, inert biological behavior, and countermeasures. J. Oncol. 2021, 2021, 5544232. [Google Scholar] [CrossRef] [PubMed]
- Zaridze, D.; Maximovitch, D.; Smans, M.; Stilidi, I. Thyroid cancer overdiagnosis revisited. Cancer Epidemiol. 2021, 74, 102014. [Google Scholar] [CrossRef] [PubMed]
- Matrone, A.; Campopiano, M.C.; Nervo, A.; Sapuppo, G.; Tavarelli, M.; de Leo, S. Differentiated thyroid cancer, from active surveillance to advanced therapy: Toward a personalized medicine. Front. Endocrinol. 2020, 10, 884. [Google Scholar] [CrossRef]
- Chou, R.; Dana, M.T.; Haymart, M.R.; Leung, A.M.; Tufano, R.P.; Sosa, J.A.; Ringel, M.D. Active surveillance versus thyroid surgery for differentiated thyroid cancer: A systematic review. Thyroid 2022, 32, 351–367. [Google Scholar] [CrossRef]
- Théry, C.W.K.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; Ayre, D.C.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the international society for extracellular vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Fan, J.; Hsu, Y.-M.S.; Lyon, C.J.; Ning, B.; Hu, T.Y. Extracellular vesicles as cancer liquid biopsies: From discovery, validation, to clinical application. Lab A Chip 2019, 19, 1114–1140. [Google Scholar] [CrossRef] [PubMed]
- Pogliaghi, G. Liquid biopsy in thyroid cancer: From circulating biomarkers to a new prospective of tumor monitoring and therapy. Minerva Endocrinol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Danielson, K.; Estanislau, J.; Tigges, J.; Toxavidis, V.; Camacho, V.; Felton, E.J.; Khoory, J.; Kreimer, S.; Ivanov, A.R.; Mantel, P.-Y.; et al. Diurnal variations of circulating extracellular vesicles measured by nano flow cytometry. PLoS ONE 2016, 11, e0144678. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, G.; Porto, W.; Palu, C.; Pereira, L.M.; Petriz, B.; Almeida, J.A.; Viana, J.; Filho, N.N.A.; Franco, O.L.; Pereira, R. Effects of acute aerobic exercise on rats serum extracellular vesicles diameter, concentration and small RNAs content. Front. Physiol. 2018, 9, 532. [Google Scholar] [CrossRef] [PubMed]
- Newman, L.; Fahmy, A.; Sorich, M.; Best, O.; Rowland, A.; Useckaite, Z. Importance of between and within subject variability in extracellular vesicle abundance and cargo when performing biomarker analyses. Cells 2021, 10, 485. [Google Scholar] [CrossRef]
- Grangier, A.; Branchu, J.; Volatron, J.; Piffoux, M.; Gazeau, F.; Wilhelm, C.; Silva, A.K. Technological advances towards extracellular vesicles mass production. Adv. Drug Deliv. Rev. 2021, 176, 113843. [Google Scholar] [CrossRef] [PubMed]
- Adlerz, K.; Patel, D.; Rowley, J.; Ng, K.; Ahsan, T. Strategies for scalable manufacturing and translation of MSC-derived extracellular vesicles. Stem Cell Res. 2020, 48, 101978. [Google Scholar] [CrossRef]
- Royo, F.; Cossío, U.; de Angulo, A.R.; Llop, J.; Falcon-Perez, J.M. Modification of the glycosylation of extracellular vesicles alters their biodistribution in mice. Nanoscale 2019, 11, 1531–1537. [Google Scholar] [CrossRef]
- Driedonks, T.; Jiang, L.; Carlson, B.; Han, Z.; Liu, G.; Queen, S.E.; Shirk, E.N.; Gololobova, O.; Nyberg, L.H.; Lima, G.; et al. Pharmacokinetics and biodistribution of extracellular vesicles administered intravenously and intranasally to Macaca nemestrina. bioRxiv 2022, 2021, 454192. [Google Scholar] [CrossRef]
- Gangadaran, P.; Li, X.J.; Kalimuthu, S.K.; Min, O.J.; Hong, C.M.; Rajendran, R.L.; Lee, H.W.; Zhu, L.; Baek, S.H.; Jeong, S.Y.; et al. New optical imaging reporter-labeled anaplastic thyroid cancer-derived extra-cellular vesicles as a platform for in vivo tumor targeting in a mouse model. Sci. Rep. 2018, 8, 13509. [Google Scholar] [CrossRef]
- Smyth, T.; Kullberg, M.; Malik, N.; Smith-Jones, P.; Graner, M.W.; Anchordoquy, T.J. Biodistribution and delivery efficiency of unmodified tumor-derived exosomes. J. Control. Release 2015, 199, 145–155. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Reference | EV Isolation Technique | Identified Content | EV Origin | Recipient Cells | Dose | Effects | In Vivo? | Link between Content and Effect |
|---|---|---|---|---|---|---|---|---|
| (Li et al., 2021) [25] | Kit | ANXA1 | SW579, Nthy-ori3-1 | SW579, Nthy-ori3-1 | ? | Malignant transformation | Xenograft | OE, KD |
| (Surman et al., 2022) [26] | dUC | Proteins | 8305C, Nthy-ori3-1 | 8305C, Nthy-ori3-1 | 30–60 µg/1 × 104 cells or /well | Viability, migration | / | / |
| (Wu et al., 2022) [27] | dUC | Cdkn2b, Cdkn2b-AS1 | Nthy-ori3-1, SW579 | TPC-1, SW579 | ? | Viability, migration, invasion | / | OE, KD |
| (Luo et al., 2018) [28] | dUC | SRC, TLN1, ITGB2 and CAPNS1 | PTC serum | BCPAP, BHT101 | 30–100 µg/3 × 104 cells | Invasion | / | / |
| (Grzanka et al., 2022) [29] | ExoQuick | / | Nthy-ori3-1, FTC6133, CGTH-W-1, BCPAP, 8505C, TPC-1 | Nthy-ori3-1, HUVEC, PBMC | ? | No effect on Nthy-ori3-1 and monocytes, ↘tube formation by HUVEC | / | / |
| (Lee et al., 2015) [30] | dUC | miR-146b, miR-222 | TPC-1, Nthy-ori3-1 | TPC-1, Nthy-ori3-1 | ? | Inhibition of proliferation | / | / |
| (Hardin et al., 2018) [31] | ExoQuick | lncRNA | TPC-1, THJ-16T, Nthy-ori3-1 | TPC-1, Nthy-ori3-1 | 5–10 µg/1 × 105 cells or 5 mg/1 × 104 cells | Proliferation, invasion | / | / |
| (Wen et al., 2021a) [32] | PEG | SNHG9 lncRNA | TPC-1, K-1, Nthy-ori3-1 | Nthy-ori3-1 | ?/co-culture | Autophagy, apoptosis | / | OE, KD |
| (Delcorte et al., 2022b) [22] | dUC+DG | miRNAs | CTL and BRAFV600E thyroid tissues | BMDM | 2 × 103 EVs/cells | BMDM polarization | / | / |
| (Wang et al., 2020) [33] | Kit | PD-L1 | PTC plasma | Activated T cells from PBMCs | ? | Immunosuppression | / | Anti-PD-L1 ab |
| (Bravo-Miana et al., 2020) (Bravo-Miana et al., 2022) [34,35] | dUC | Proteins | Fb, 8505C, TPC-1, Nthy-ori3-1/co-culture | Fb, 8505C, TPC-1, Nthy-ori3-1 | ? | MMP2 activation | / | / |
| (Wang et al., 2021) [36] | dUC, ExoQuick | miR-181a | BCPAP, K-1, Nthy-ori3-1 | HUVEC | ? | Tube formation | Xenograft | anti-miR-181a |
| (Wu F, 2019) [37] | dUC | miR-21-5p | BCPAP, KTC-1, Nthy-ori3-1 | HUVEC | ? | Angiogenesis | / | anti-miR-21-5p |
| (Yin et al., 2021) [38] | Kit | miR-130a-3p | DTC plasma | TPC-1 | ? | ↗IGF-1, p-PI3K, p-AKT and migration | / | siRNA anti-Igf1 |
| (Vella et al., 2017) [39] | dUC+DG | PDGFRb | Melanoma cell lines | Melanoma cell lines | 50–200 µg/mL on 4 × 105 cells | Resistance to treatment | / | neutralizing PDGFRβ ab |
| Reference | Initial Material | Patients | EV Isolation Technique | Content Analysis | Differential Content |
|---|---|---|---|---|---|
| (Zabegina et al., 2020) [47] | Plasma | 30 FTC, 30 FA | dUC, immuno-capture | qRT-PCR | let-7 |
| (Huang et al., 2020) [48] | Urine | 16 PTC, FTC | Kit | LC-MRM/MS | Thyroglobulin |
| (Caruso Bavisotto et al., 2019) [49] | Plasma | 13 PTC, 18 MNG | dUC | WB | HSP27, HSP60, HSP90 |
| (Luo et al., 2018) [28] | Serum | 16 PTC-N1, 17 PTC-N0 | dUC | LC-MS/MS | SRC, TLN1, ITGB2 and CAPNS1 |
| (Samsonov, 2016) [40] | Plasma | 60 PTC, FTC, BN | dUC | qRT-PCR | miR-21, miR-181a |
| (Wen et al., 2021b) [50] | Serum | 119 PTC, 100 HC | ExoQuick | qRT-PCR | miR-29a |
| (Yin et al., 2021) [38] | Plasma | 40 DTC, 40 BN | Kit | qRT-PCR | miR-130a-3p |
| (Wang et al., 2020) [51] | Plasma | 43 PTC | Kit | ELISA | PD-L1 |
| (Liang, 2020) [52] | Plasma | 51 PTC, 38 MNG | Kit | NGS | 6 miRNA signature |
| (Xin et al., 2022) [53] | TGCA data | 491 PTC | / | / | 6 miRNA signature |
| (Delcorte et al., 2022a) [54] | Plasma | 19 MNG, 17 PTC | IDC, SEC, UF | qRT-PCR | miR-146b-5p, miR-21a-5p |
| (Capriglione F, 2021) [55] | Serum | 56 PTC-N1, 58 PTC-N0 | ExoQuick | miRNA array cards | miR24-3p, miR146a-5p, miR181a-5p and miR382-5p |
| (Jiang et al., 2020) [56] | Plasma | 49 PTC-N1, 15 PTC-N0 | Kit | qRT-PCR | miR-146b-5p, miR-222-3p |
| (Chen et al., 2022) [57] | Plasma | 34 PTC-N1, 34 PTC-N0 | dUC | microarray | miR-6774-3p, miR-6879-5p |
| (Yang et al., 2019) [59] | Serum | 3 PTC, 3 MNG | Kit | RNA seq | CircRNAs |
| (Pan et al., 2019) [60] | Plasma | 13 PTC, 7 MNG | dUC, DG | NGS | 126 DE miRNAs |
| (Dai et al., 2020) [61] | Serum | 136 PTC, 92 MNG, 51 HC | dUC | small RNA seq | miR-485-3p, miR-4433a-5p |
| Reference | EV Isolation | Content Analysis | Differential Content | EV origin | Recipient Cells | Dose | Effects |
|---|---|---|---|---|---|---|---|
| (Cui et al., 2019) [73] | High-speed dUC | WB | TPO, HSP60, MHC-II | HT serum | DCs, PBMCs | 600 µg- EVs/1 × 106 cells | Action via TLR2/3 DCs: ↗IL-6, NF-kB PBMCs: Th1, Th17, Treg diff. |
| (Cui et al., 2020) [74] | High-speed dUC | WB | TPO, HSP60, MHC-II | IFN-γ-treated Nthy | DCs | Transwell co-culture | DC activation, CD4+ T ly pro-inflammatory response |
| (Cui et al., 2020) [75] | High-speed dUC | WB | IGF-1R, HSP60 | GD serum | PBMCs | 400 µg-EVs/2 × 106 cells | Action via TLR2/3 ↗IL-6, IL-1b NF-Kb pathway |
| (Rodríguez-Muñoz et al.) [76] | Medium-speed UC | qRT-PCR | miR-146a, miR-155 | AITD plasma | Naive CD4+ T cells | 1mL plasma-MVs/2 × 105 cells | ↗ Th17 and ↘Treg differentiation |
| (Han et al., 2021) [77] | Kit | Proteome profiler | VBD, CRP, CHI3L1, MMP-9, VCAM-1 | GD tear fluids | Orbital fibroblasts | 5 µg/1 × 104 cells | ↗ IL-6, IL-8, MCP-1 |
| (Hiratsuka et al., 2016) [78] | Kit | / | / | GD serum | PBMCs | ? | ↗ expression of TNF-a, IL-1b, IL-6 |
| Reference | Initial Material | Patients | EV Isolation Technique | Content Analysis | Findings |
|---|---|---|---|---|---|
| (Cui et al., 2019) [73] | Serum | 30 HT, 30 HC | High-speed dUC | WB | TPO, HSP60, MHCII: HT > HC. No difference in TG, HMGB1, ICAM1 |
| (Cui et al., 2021) [75] | Serum | 33 GD, HC | High-speed dUC | WB | IGF-1R, HSP60: GD > HC |
| (Jia et al., 2021) [80] | Plasma | 12 GD, 10 HT, 7 HC | High-speed dUC | Proteomics | List of differential proteins implicated in the immune and metabolic systems |
| (Rodríguez-Muñoz et al.) [76] | Plasma | 33 GD, 29 HT, 45 HC | Medium-speed UC | qRT-PCR | Mir-146a, miR-155: AITD > HC |
| (Sun et al., 2020) [82] | Plasma | 25 GD, 25 HC | High-speed dUC | CircRNA microarray | 15 DEcRs hsa_circRNA_000102: GD > HC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delcorte, O.; Degosserie, J.; Pierreux, C.E. Role of Extracellular Vesicles in Thyroid Physiology and Diseases: Implications for Diagnosis and Treatment. Biomedicines 2022, 10, 2585. https://doi.org/10.3390/biomedicines10102585
Delcorte O, Degosserie J, Pierreux CE. Role of Extracellular Vesicles in Thyroid Physiology and Diseases: Implications for Diagnosis and Treatment. Biomedicines. 2022; 10(10):2585. https://doi.org/10.3390/biomedicines10102585
Chicago/Turabian StyleDelcorte, Ophélie, Jonathan Degosserie, and Christophe E. Pierreux. 2022. "Role of Extracellular Vesicles in Thyroid Physiology and Diseases: Implications for Diagnosis and Treatment" Biomedicines 10, no. 10: 2585. https://doi.org/10.3390/biomedicines10102585
APA StyleDelcorte, O., Degosserie, J., & Pierreux, C. E. (2022). Role of Extracellular Vesicles in Thyroid Physiology and Diseases: Implications for Diagnosis and Treatment. Biomedicines, 10(10), 2585. https://doi.org/10.3390/biomedicines10102585