

Amniotic Fluid-Derived Mesenchymal Stem/Stromal Cell-Derived Secretome and Exosomes Improve Inflammation in Human Intestinal Subepithelial Myofibroblasts
 ,
,  , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Isolation, Characterization, and Culture of Human Intestinal Subepithelial Myofibroblasts
2.3. CM Preparation
2.4. Exosome Isolation, Western Blot Characterization, and Transmission Electron Microscopy (TEM) for Exosome Imaging
2.5. Inflammation Induction and CM/Exosome Incubation and RNA Extraction
2.6. cDNA Synthesis and Real Time PCR
2.7. Extracellular IL-1β Quantification
2.8. Statistical Analysis
3. Results
3.1. Exosome Characterization
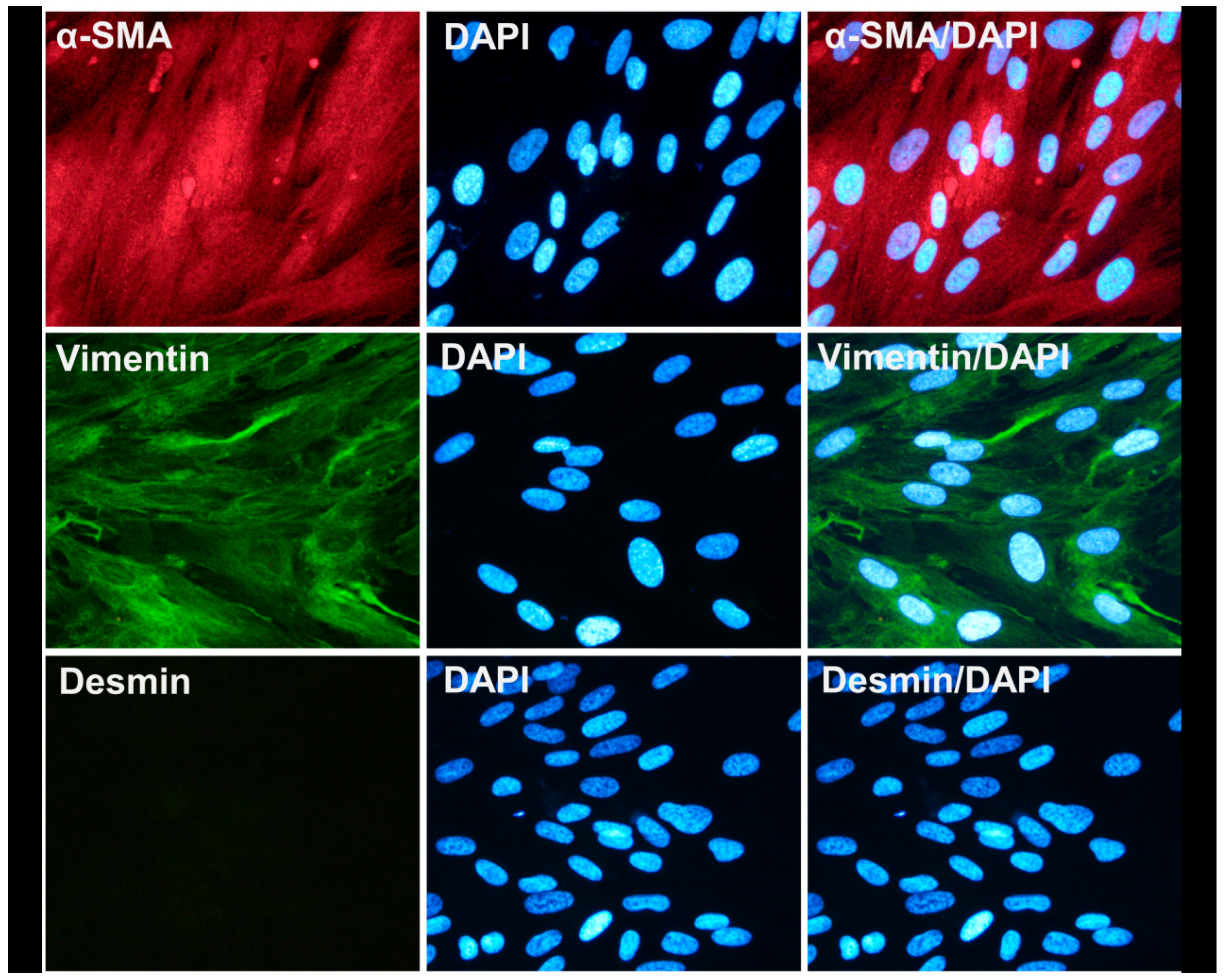
3.2. Characterization of Human Intestinal Subepithelial Myofibroblasts
3.3. Confirmation of Inflammation Model
3.4. Therapeutic Effects of CM and Exosomes
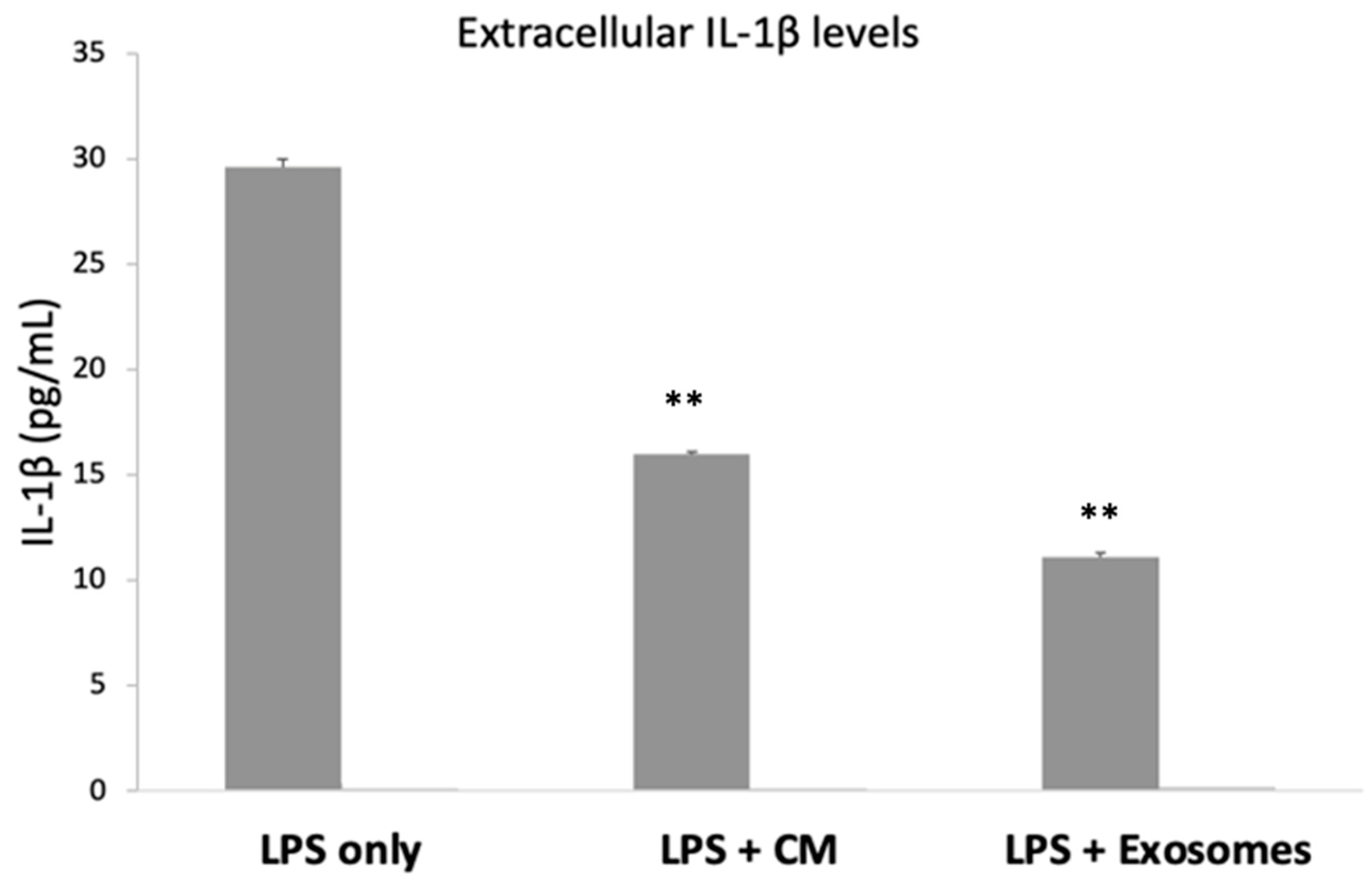
3.5. Extracellular IL-1β
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Seyed Tabib, N.S.; Madgwick, M.; Sudhakar, P.; Verstockt, B.; Korcsmaros, T.; Vermeire, S. Big data in IBD: Big progress for clinical practice. Gut 2020, 69, 1520–1532. [Google Scholar] [CrossRef] [PubMed]
- Verstockt, B.; Ferrante, M.; Vermeire, S.; Van Assche, G. New treatment options for inflammatory bowel diseases. J. Gastroenterol. 2018, 53, 585–590. [Google Scholar] [CrossRef] [PubMed]
- Bryant, R.V.; Winer, S.; Travis, S.P.; Riddell, R.H. Systematic review: Histological remission in inflammatory bowel disease. Is ‘complete’ remission the new treatment paradigm? An IOIBD initiative. J. Crohn’s Colitis. 2014, 8, 1582–1597. [Google Scholar] [CrossRef] [PubMed]
- Herrlinger, K.R.; Stange, E.F. Twenty-five years of biologicals in IBD: What’s all the hype about? J. Intern. Med. 2021, 290, 806–825. [Google Scholar] [CrossRef]
- Galderisi, U.; Peluso, G.; Di Bernardo, G. Clinical Trials Based on Mesenchymal Stromal Cells are Exponentially Increasing: Where are We in Recent Years? Stem Cell Rev. Rep. 2022, 18, 23–36. [Google Scholar] [CrossRef]
- Montemurro, T.; Viganò, M.; Ragni, E.; Barilani, M.; Parazzi, V.; Boldrin, V.; Lavazza, C.; Montelatici, E.; Banfi, F.; Lauri, E.; et al. Angiogenic and anti-inflammatory properties of mesenchymal stem cells from cord blood: Soluble factors and extracellular vesicles for cell regeneration. Eur. J. Cell Biol. 2016, 95, 228–238. [Google Scholar] [CrossRef]
- Buscail, E.; Le Cosquer, G.; Gross, F.; Lebrin, M.; Bugarel, L.; Dedraison, C.; Vergnolle, N.; Bournet, B.; Gilletta, C.; Buscail, L. Adipose-Derived Stem Cells in the Treatment of Perianal Fistulas in Crohn’s Disease: Rationale, Clinical Results and Perspectives. Int. J. Mol. Sci. 2021, 22, 9967. [Google Scholar] [CrossRef]
- Duijvestein, M.; Vos, A.C.W.; Roelofs, H.; Wildenberg, M.E.; Wendrich, B.B.; Verspaget, H.W.; Kooy-Winkelaar, E.M.; Koning, F.; Zwaginga, J.J.; Fidder, H.H.; et al. Autologous bone marrow-derived mesenchymal stromal cell treatment for refractory luminal Crohn’s disease: Results of a phase I study. Gut 2010, 59, 1662–1669. [Google Scholar] [CrossRef]
- Xia, J.; Minamino, S.; Kuwabara, K.; Arai, S. Stem cell secretome as a new booster for regenerative medicine. Biosci. Trends 2019, 13, 299–307. [Google Scholar] [CrossRef]
- Musiał-Wysocka, A.; Kot, M.; Majka, M. The Pros and Cons of Mesenchymal Stem Cell-Based Therapies. Cell Transplant. 2019, 28, 801–812. [Google Scholar] [CrossRef]
- Ayyar, K.K.; Moss, A.C. Exosomes in Intestinal Inflammation. Front. Pharmacol. 2021, 12, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Ha, D.H.; Kim, H.K.; Lee, J.; Kwon, H.H.; Park, G.H.; Yang, S.H.; Jung, J.Y.; Choi, H.; Lee, J.H.; Sung, S.; et al. Mesenchymal Stem/Stromal Cell-Derived Exosomes for Immunomodulatory Therapeutics and Skin Regeneration. Cells 2020, 9, 1157. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Yang, X.; Xiao, X.; Xu, M.; Yang, Y.; Xue, C.; Li, X.; Wang, S.; Zhao, R.C. Human Adipose Mesenchymal Stem Cell-derived Exosomes Protect Mice from DSS-Induced Inflammatory Bowel Disease by Promoting Intestinal-stem-cell and Epithelial Regeneration. Aging Dis. 2021, 12, 1423–1437. [Google Scholar] [CrossRef]
- Spitzhorn, L.S.; Rahman, M.S.; Schwindt, L.; Ho, H.T.; Wruck, W.; Bohndorf, M.; Wehrmeyer, S.; Ncube, A.; Beyer, I.; Hagenbeck, C.; et al. Isolation and Molecular Characterization of Amniotic Fluid-Derived Mesenchymal Stem Cells Obtained from Caesarean Sections. Stem Cells Int. 2017, 2017, 5932706. [Google Scholar] [CrossRef] [PubMed]
- Zagoura, D.S.; Roubelakis, M.G.; Bitsika, V.; Trohatou, O.; Pappa, K.I.; Kapelouzou, A.; Antsakli, A.; Anagnou, N.P. Therapeutic potential of a distinct population of human amniotic fluid mesenchymal stem cells and their secreted molecules in mice with acute hepatic failure. Gut 2012, 61, 894–906. [Google Scholar] [CrossRef]
- Zagoura, D.; Trohatou, O.; Makridakis, M.; Kollia, A.; Kokla, N.; Moukou, M.; Psaraki, A.; Eliopoulos, A.G.; Vlahou, A.; Roubelakis, M.G. Functional secretome analysis reveals Annexin-A1 as important paracrine factor derived from fetal mesenchymal stem cells in hepatic regeneration. EBioMedicine 2019, 45, 542–552. [Google Scholar] [CrossRef]
- Valatas, V.; Filidou, E.; Drygiannakis, I.; Kolios, G. Stromal and immune cells in gut fibrosis: The myofibroblast and the scarface. Ann. Gastroenterol. 2017, 30, 393–404. [Google Scholar] [CrossRef]
- Filidou, E.; Valatas, V.; Drygiannakis, I.; Arvanitidis, K.; Vradelis, S.; Kouklakis, G.; Kolios, G.; Bamias, G. Cytokine receptor profiling in human colonic subepithelial myofibroblasts: A differential effect of Th polarization associated cytokines in intestinal fibrosis. Inflamm. Bowel. Dis. 2018, 24, 2224–2241. [Google Scholar] [CrossRef]
- Bamias, G.; Filidou, E.; Goukos, D.; Valatas, V.; Arvanitidis, K.; Panagopoulou, M.; Kouklakis, G.; Daikos, G.L.; Ladas, S.D.; Kolios, G. Crohn’s disease-associated mucosal factors regulate the expression of TNF-like cytokine 1A and its receptors in primary subepithelial intestinal myofibroblasts and intestinal epithelial cells. Transl. Res. 2017, 180, 18–30. [Google Scholar] [CrossRef]
- Okogbule-Wonodi, A.C.; Li, G.; Anand, B.; Luzina, I.G.; Atamas, S.P.; Blanchard, T. Human foetal intestinal fibroblasts are hyper-responsive to lipopolysaccharide stimulation. Dig. Liver Dis. 2012, 44, 18–23. [Google Scholar] [CrossRef]
- Legaki, E.; Roubelakis, M.G.; Theodoropoulos, G.E.; Lazaris, A.; Kollia, A.; Karamanolis, G.; Marinos, E.; Gazouli, M. Therapeutic Potential of Secreted Molecules Derived from Human Amniotic Fluid Mesenchymal Stem/Stroma Cells in a Mice Model of Colitis. Stem Cell Rev. Rep. 2016, 12, 604–612. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.C.; Yeh, W.C.; Ohashi, P.S. LPS/TLR4 signal transduction pathway. Cytokine 2008, 42, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, S.; Wang, W.; Qin, W.; Cheng, K.; Coulup, S.; Chavez, S.; Jiang, S.; Raparia, K.; De Almeida, L.M.V.; Stelhik, C.; et al. TLR4-dependent fibroblast activation drives persistent organ fibrosis in skin and lung. JCI Insight 2018, 3, e98850. [Google Scholar] [CrossRef] [PubMed]
- Gordon, I.O.; Agrawal, N.; Willis, E.; Goldblum, J.R.; Lopez, R.; Allende, D.; Liu, X.; Patil, D.Y.; Yerian, L.; El-Khider, F.; et al. Fibrosis in ulcerative colitis is directly linked to severity and chronicity of mucosal inflammation. Aliment Pharmacol. Ther. 2018, 47, 922–939. [Google Scholar] [CrossRef]
- Alfredsson, J.; Wick, M.J. Mechanism of fibrosis and stricture formation in Crohn’s disease. Scand. J. Immunol. 2020, 92, e12990. [Google Scholar] [CrossRef] [PubMed]
- Roubelakis, M.G.; Bitsika, V.; Zagoura, D.; Trohatou, O.; Pappa, K.I.; Makridakis, M.; Antsaklis, A.; Vlahou, A.; Anagnou, N.P. In vitro and in vivo properties of distinct populations of amniotic fluid mesenchymal progenitor cells. J. Cell. Mol. Med. 2011, 15, 1896–1913. [Google Scholar] [CrossRef]
- Zagoura, D.; Trohatou, O.; Bitsika, V.; Makridakis, M.; Pappa, K.I.; Vlahou, A.; Roubelakis, M.G.; Anagnou, N.P. AF-MSCs fate can be regulated by culture conditions. Cell Death Dis. 2013, 4, e571. [Google Scholar] [CrossRef]
- Furuta, T.; Miyaki, S.; Ishitobi, H.; Ogura, T.; Kato, Y.; Kamei, N.; Miyado, K.; Higashi, Y.; Ochi, M. Mesenchymal Stem Cell-Derived Exosomes Promote Fracture Healing in a Mouse Model. Stem Cells Transl. Med. 2016, 5, 1620–1630. [Google Scholar] [CrossRef]
- Elkouris, M.; Kouroupi, G.; Vourvoukelis, A.; Papagiannakis, N.; Kaltezioti, V.; Matsas, R.; Stefanis, L.; Xilouri, M.; Politis, P.K. Long Non-coding RNAs Associated With Neurodegeneration-Linked Genes Are Reduced in Parkinson’s Disease Patients. Front. Cell Neurosci. 2019, 13, 58. [Google Scholar] [CrossRef]
- Li, C.; Kuemmerle, J.F. The fate of myofibroblasts during the development of fibrosisin Crohn’s disease. J. Dig. Dis. 2020, 21, 326–331. [Google Scholar] [CrossRef]
- Strober, W.; Fuss, I.J. Proinflammatory cytokines in the pathogenesis of inflammatory bowel diseases. Gastroenterology 2011, 140, 1756–1767. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Liang, X.; Song, J.; Li, C.; Liu, A.; Luo, Y.; Ma, H.; Tan, Y.; Zhang, X. A novel therapeutic approach for inflammatory bowel disease by exosomes derived from human umbilical cord mesenchymal stem cells to repair intestinal barrier via TSG-6. Stem Cell Res. Ther. 2021, 12, 315. [Google Scholar] [CrossRef] [PubMed]
- Ocansey, D.K.W.; Zhang, L.; Wang, Y.; Yan, Y.; Qian, H.; Zhang, X.; Xu, W.; Mao, F. Exosome-mediated effects and applications in inflammatory bowel disease. Biol. Rev. 2020, 95, 1287–1307. [Google Scholar] [CrossRef] [PubMed]
- Mao, F.; Wu, Y.; Tang, X.; Kang, J.; Zhang, B.; Yan, Y.; Qian, H.; Zhang, X.; Xu, W. Exosomes Derived from Human Umbilical Cord Mesenchymal Stem Cells Relieve Inflammatory Bowel Disease in Mice. BioMed Res. Int. 2017, 2017, 5356760. [Google Scholar] [CrossRef]
- D’Haens, G.R.; van Deventer, S. 25 years of anti-TNF treatment for inflammatory bowel disease: Lessons from the past and a look to the future. Gut 2021, 70, 1396–1405. [Google Scholar] [CrossRef]
- Vulliemoz, M.; Brand, S.; Juillerat, P.; Mottet, C.; Ben-Horin, S.; Michetti, P. TNF-Alpha Blockers in Inflammatory Bowel Diseases: Practical Recommendations and a User’s Guide: An Update. Digestion 2020, 101, 16–26. [Google Scholar] [CrossRef]
- Allocca, M.; Jovani, M.; Fiorino, G.; Schreiber, S.; Danese, S. Anti-IL-6 treatment for inflammatory bowel diseases: Next cytokine, next target. Curr. Drug Targets 2013, 14, 1508–1521. [Google Scholar] [CrossRef]
- Walana, W.; Ye, Y.; Li, M.; Wang, M.; Wang, B.; Cheng, J.W.; Gordon, J.R.; Li, F. IL-8 antagonist, CXCL8(3-72)K11R/G31P coupled with probiotic exhibit variably enhanced therapeutic potential in ameliorating ulcerative colitis. Biomed. Pharmacother. 2018, 103, 253–261. [Google Scholar] [CrossRef]
- Chen, Y.T.; Tsai, M.J.; Hsieh, N.; Ming-Jei, L.; Meng-Jen, L.; Cheng, H.; Huang, W.C. The superiority of conditioned medium derived from rapidly expanded mesenchymal stem cells for neural repair. Stem Cell Res. Ther. 2019, 10, 390. [Google Scholar] [CrossRef]
- Shen, Q.; Huang, Z.; Yao, J.; Jin, Y. Extracellular vesicles-mediated interaction within intestinal microenvironment in inflammatory bowel disease. J. Adv. Res. 2022, 37, 221–233. [Google Scholar] [CrossRef]
- Dinarello, C.A.; Simon, A.; van der Meer, J.W. Treating inflammation by blocking interleukin-1 in a broad spectrum of diseases. Nat. Rev. Drug Discov. 2012, 11, 633–652. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Corbett, A.L.; Taatizadeh, E.; Tasnim, N.; Little, J.P.; Garnis, C.; Daugaard, M.; Guns, E.; Hoorfar, M.; Li, I.T.S. Challenges and opportunities in exosome research—Perspectives from biology, engineering, and cancer therapy. APL Bioeng. 2019, 3, 011503. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| IL-1β | 5′-CCTTGTCGAGAATGGGCAGT-3′ | 5′-TCCTGTCGACAATGCTGCCT-3′ |
| IL-6 | 5′-CCAGCTATGAACTCCTTCT-3′ | 5′-GCTTGTTCCTCACATCTCT-3′ |
| IL-8 | 5′-ACTGAGAGTGATTGAGAGTGGAC-3′ | 5′-AACCCTCTGCACCCAGTTTTC-3′ |
| IL-10 | 5′-TCTCCGAGATGCCTTCAGCAGA-3′ | 5′-TCAGACAAGGCTTGGCAACCCA-3′ |
| ANXA1 | 5′-GCAGGCCTGGTTTATTGAAA-3′ | 5′-GCTGTGCATTGTTTCGCTTA-3′ |
| TNF-α | 5′-CACGTCGTAGCAAACCACCAAGTGG-3′ | 5′-GATAGCAAATCGGCTGACGGTGTGG-3′ |
| TLR-4 | 5′-GATTAGCATACTTAGACTAC-3′ | 5′-GATCAACTTCTGAAAAAGCATTCCCAC-3′ |
| GAPDH | 5′-TTCACCACCATGGAGAAGGC-3′ | 5′-GGCATGGACTGTGGTCATGA-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Katifelis, H.; Filidou, E.; Psaraki, A.; Yakoub, F.; Roubelakis, M.G.; Tarapatzi, G.; Vradelis, S.; Bamias, G.; Kolios, G.; Gazouli, M. Amniotic Fluid-Derived Mesenchymal Stem/Stromal Cell-Derived Secretome and Exosomes Improve Inflammation in Human Intestinal Subepithelial Myofibroblasts. Biomedicines 2022, 10, 2357. https://doi.org/10.3390/biomedicines10102357
Katifelis H, Filidou E, Psaraki A, Yakoub F, Roubelakis MG, Tarapatzi G, Vradelis S, Bamias G, Kolios G, Gazouli M. Amniotic Fluid-Derived Mesenchymal Stem/Stromal Cell-Derived Secretome and Exosomes Improve Inflammation in Human Intestinal Subepithelial Myofibroblasts. Biomedicines. 2022; 10(10):2357. https://doi.org/10.3390/biomedicines10102357
Chicago/Turabian StyleKatifelis, Hector, Eirini Filidou, Adriana Psaraki, Farinta Yakoub, Maria G. Roubelakis, Gesthimani Tarapatzi, Stergios Vradelis, Giorgos Bamias, George Kolios, and Maria Gazouli. 2022. "Amniotic Fluid-Derived Mesenchymal Stem/Stromal Cell-Derived Secretome and Exosomes Improve Inflammation in Human Intestinal Subepithelial Myofibroblasts" Biomedicines 10, no. 10: 2357. https://doi.org/10.3390/biomedicines10102357
APA StyleKatifelis, H., Filidou, E., Psaraki, A., Yakoub, F., Roubelakis, M. G., Tarapatzi, G., Vradelis, S., Bamias, G., Kolios, G., & Gazouli, M. (2022). Amniotic Fluid-Derived Mesenchymal Stem/Stromal Cell-Derived Secretome and Exosomes Improve Inflammation in Human Intestinal Subepithelial Myofibroblasts. Biomedicines, 10(10), 2357. https://doi.org/10.3390/biomedicines10102357