A Revision of the Taxonomy and Identification of Epipactis greuteri (Orchidaceae, Neottieae)

 ,
,  , ,
, ,  , and
, and
Abstract

1. Introduction
2. Results
2.1. Herbarium Material Investigations—Reexamination of the Holotype and Isotype
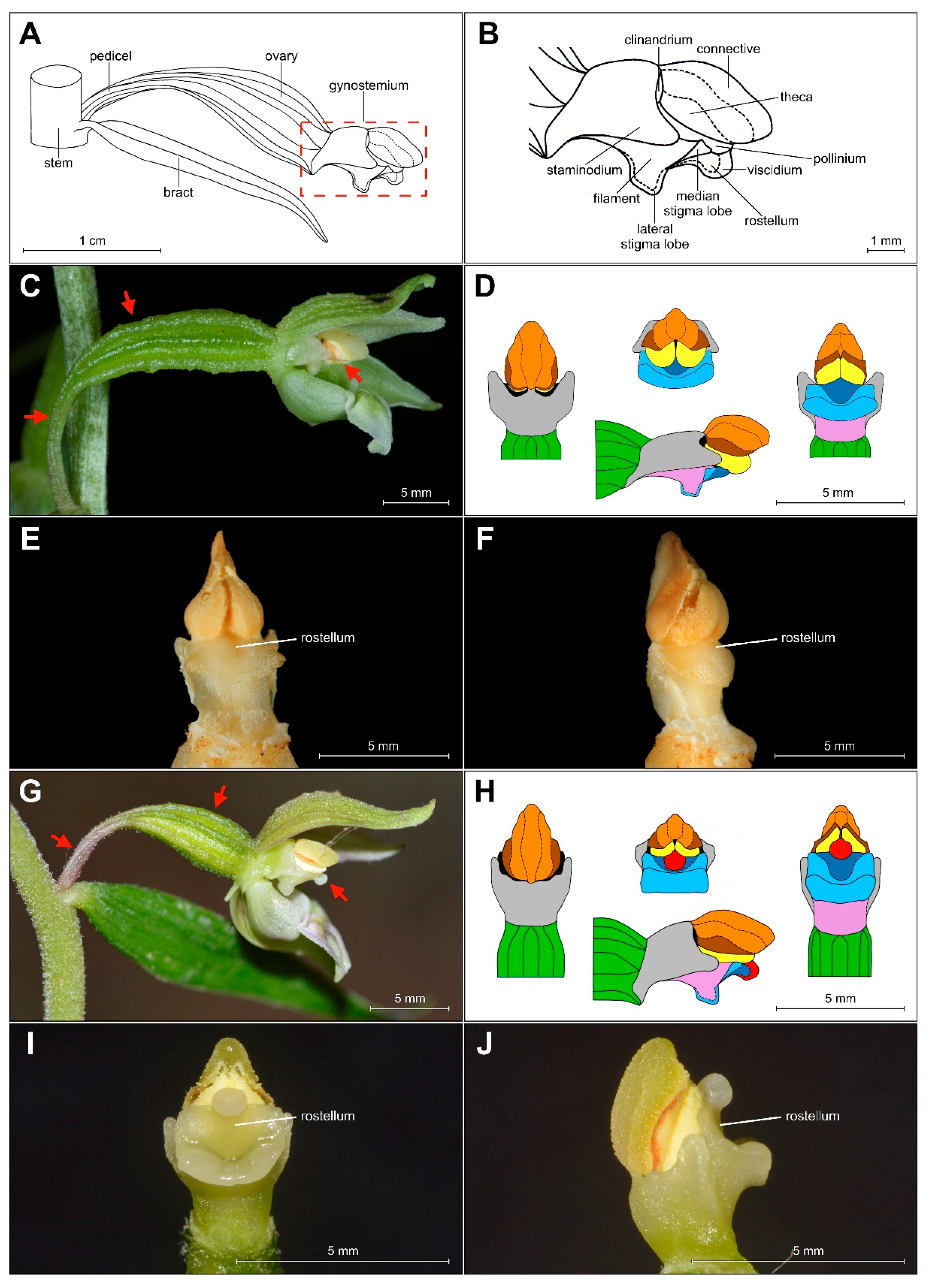
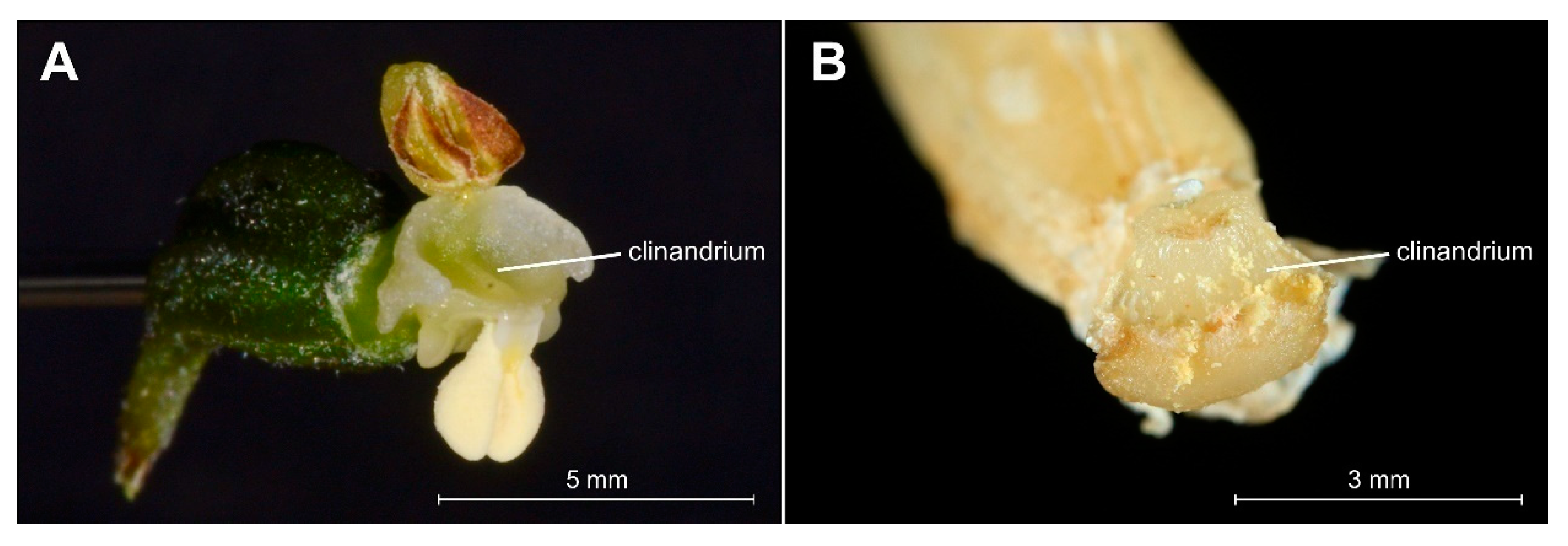
2.2. Field Research—Morphological Analyses
Micromorphology of Papillae
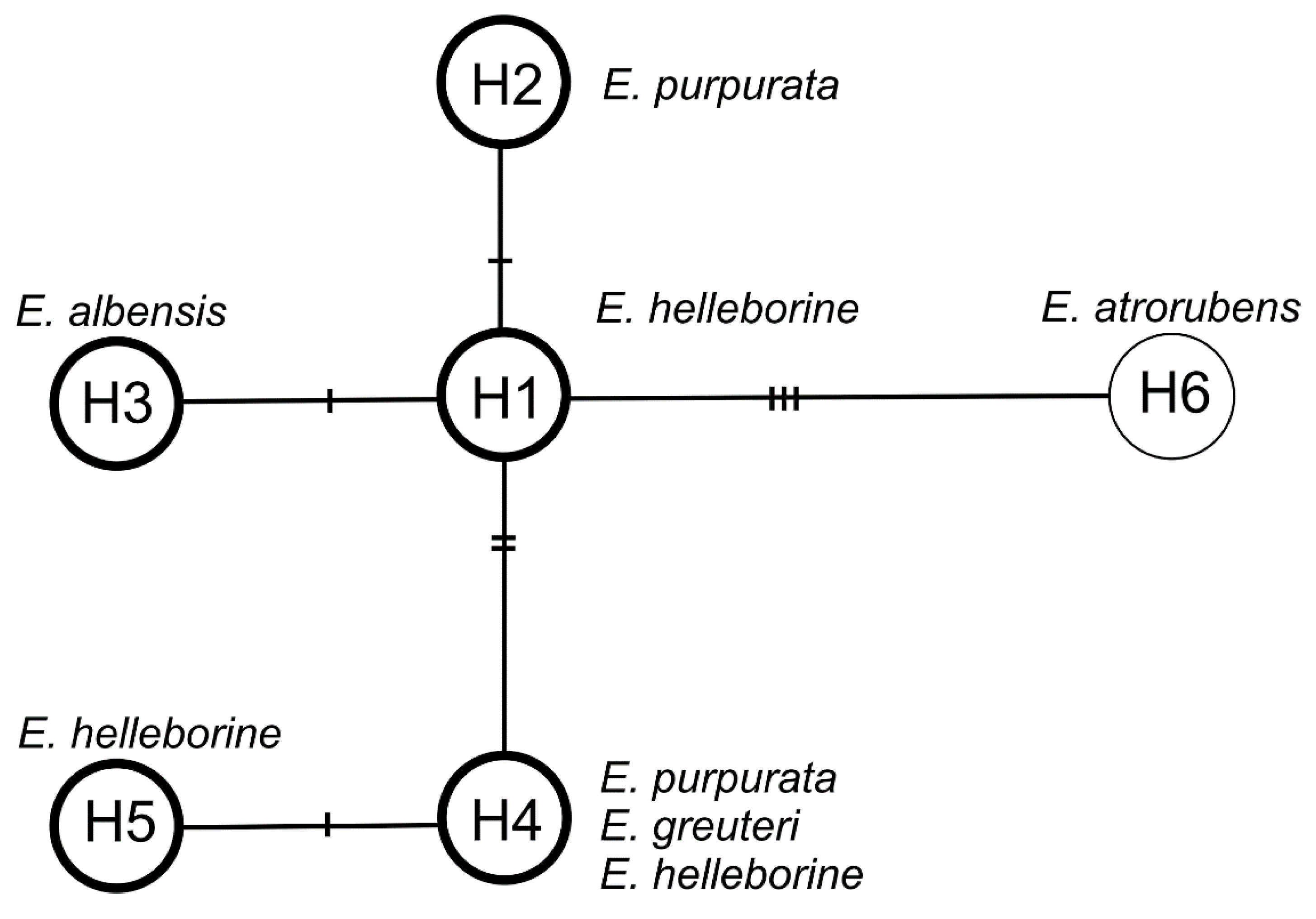
2.3. Molecular Data
3. Discussion
3.1. Diagnostic Characters in the Protologue of Epipactis greuteri
3.2. The Controversy on the Taxonomic Position of the Epipactis greuteri Infraspecific Taxa
3.3. Species Variability in Taxonomic Context
3.4. Molecular Data
4. Materials and Methods
4.1. Herbarium Material Investigations
4.2. Field Research
4.3. Microscopic Analyses of the Papillae Morphology
4.4. Molecular Analyses
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
References
- Delforge, P. Orchids of Europe, North Africa and the Middle East; A&C Black: London, UK, 2006. [Google Scholar]
- Hermjakob, G. Eine für Griechenland neue, autogame Epipactis-Art: E. leptochila (Godf.) Godf. (Orchidaceae). Ann. Mus. Goulandris 1973, 1, 99–103. [Google Scholar]
- Baumann, H.; Künkele, S. Beiträge zur Taxonomie europäischer Orchideenarten. Mitt. Bl. Arbeitskr. Heim. Orch. Baden Württ. 1981, 13, 337–374. [Google Scholar]
- Tsiftsis, S.; Antonopoulos, Z. Atlas of the Greek Orchids; Mediterraneo Editions: Rethimno, Greece, 2017; Volume I. [Google Scholar]
- Petrova, A.S.; Venkova, D.Y. Epipactis exilis and E. greuteri (Orchidaceae) in the Bulgarian flora. Phytol. Balc. 2008, 14, 69–73. [Google Scholar]
- Ardelean, C. Epipactis greuteri (Orchidaceae) a new species for Romanian. J. Eur. Orch. 2011, 43, 537. [Google Scholar]
- Průša, D. Orchideje, České Republiky; Computer Press: Brno, Czech, 2005. [Google Scholar]
- Robatsch, K. Beiträge zur Kenntnis der europäischer Epipactis-Arten (Orchidaceae). Linz. Biol. Beitr. 1988, 20, 161–172. [Google Scholar]
- Robatsch, K. Beiträge zur Kenntnis der europäischer Epipactis-Arten (Orchidaceae). Linz. Biol. Beitr. 1989, 21, 295–302. [Google Scholar]
- Savelli, P.R.; Alessandrini, A.; Liverani, P. Epipactis purpurata Smith ed E. greuteri H. Baumann et Künkele in Italia, con nuove località di E. persica (Soó) Nannfeldt (Orchidaceae). Arch. Bot. Ital. 1989, 64, 192–198. [Google Scholar]
- Bongiorni, L.; De Vito, R.; Fori, S.; Pisani, G.; Romolini, R. Epipactis pontica Taubenheim e Epipactis greuteri H. Baumann & Künkele (Orchidaceae) nuove località per l’Italia. J. Eur. Orch. 2006, 38, 907–916. [Google Scholar]
- Mrkvicka, A.C. Erstnachweis von Epipactis greuteri H. Baumann & Künkele in Österreich. Mitt. Bl. Arbeitskr. Heim. Orch. Baden Württ. 1992, 24, 450–456. [Google Scholar]
- Mereda, P.J. Epipactis greuteri (Orchidaceae)—A new species of the Slovak flora. Sect. Bot. 2000, 55, 49–55. [Google Scholar]
- Vlčko, J.; Dítě, D.; Kolník, M. Orchids of Slovakia; ZO SZOPK Orchidea: Zvolen, Slovakia, 2003. [Google Scholar]
- Nikolić, T.; Topić, J. (Eds.) Crvena Knjiga Vaskularne Flore Hrvatske; Ministarstvo kulture Republike Hrvatske, Državni zavod za zaštitu prirode: Zagreb, Croatia, 2005. [Google Scholar]
- Jogan, N. Epipactis Zinn–močvirnica. In Mala Flora Slovenije. Ključ za Določanje Praprotnic in Semenk. Četrta, Dopolnjena in Spremenjena Izdaja; Martinčič, A., Wraber, T., Jogan, N., Podobnik, A., Turk, B., Vreš, B., Ravnik, V., Frajman, B., Strgulc Krajšek, S., et al., Eds.; Tehniška Založba Slovenije: Ljubljana, Slovenia, 2007; pp. 763–767. [Google Scholar]
- Szeląg, Z.; Bernacki, L.; Pawelec, J.; Stawowczyk, K.; Wolanin, M. Epipactis greuteri (Orchidaceae) in Poland. Pol. Bot. J. 2017, 62, 117–121. [Google Scholar] [CrossRef]
- Delforge, P. Epipactis phyllanthes G.E. Smith en France et en Espagne—Données nouvelles, révision systématique et conséquences taxonomiques dans le genre Epipactis. Nat. Belg. 1997, 78, 223–256. [Google Scholar]
- Jakubska-Busse, A.; Gola, E. Validation of leaf undulation traits in the taxonomy of Epipactis muelleri Godfery, 1921 (Orchidaceae: Neottieae). Plant Syst. Evol. 2014, 7, 1707–1717. [Google Scholar] [CrossRef]
- AHO-Bayern e.V. Available online: http://www.aho-bayern.de (accessed on 15 May 2020).
- Sramkó, G.; Paun, O.; Brandrud, M.K.; Laczkó, L.; Molnár, A.; Bateman, R.M. Iterative allogamy–autogamy transitions drive actual and incipient speciation during the ongoing evolutionary radiation within the orchid genus Epipactis (Orchidaceae). Ann. Bot. 2019, 124, 481–497. [Google Scholar] [CrossRef]
- Baumann, H.; Künkele, S.; Lorenz, R. Orchideen Europas: Mit angrenzenden Gebieten; Eugen Ulmer: Stuttgart, Germany, 2006. [Google Scholar]
- Darwin, C.R. On the Various Contrivances by which British and Foreign Orchids Are Fertilised by Insects; 7th Impression of the 2nd ed; John Murray: London, UK, 1904. [Google Scholar]
- Richard, L.C.M. De Orchideis Europaeis Annotationes. Mém. Mus. Hist. Nat. 1818, 4, 23–61. [Google Scholar]
- Rasmussen, F.N. The gynostemium of the neottioid orchids. Opera Bot. 1982, 65, 1–96. [Google Scholar]
- Rasmussen, F.N. Orchids. In The Families of the Monocotyledons—Structure, Evolution and Taxonomy; Dahlgren, R., Cliford, H.T., Yeo, P.F., Eds.; Springer: Berlin, Germany, 1985; pp. 249–274. [Google Scholar]
- Burns-Balogh, P.; Funk, V.A. A phylogenetic analysis of the Orchidaceae. Smithson. Contrib. Bot. 1986, 611, 1–79. [Google Scholar] [CrossRef]
- Hagerup, O. The morphology and biology of some primitive orchid flowers. Phytomorphology 1952, 2, 134–138. [Google Scholar]
- Vermeulen, P. The different structure of the rostellum in Ophrydeae and Neottieae. Acta Bot. Neerl. 1959, 8, 338–355. [Google Scholar] [CrossRef]
- Vermeulen, P. The system of the Orchidales. Acta Bot. Neerl. 1966, 15, 224–253. [Google Scholar] [CrossRef]
- Garay, L.A. On the origin of the Orchidaceae. Bot. Mus. Leafl. Harv. Univ. 1960, 19, 57–96. [Google Scholar]
- Garay, L.A. On the origin of the Orchidaceae. II. J. Arnold Arbor. 1972, 53, 202–215. [Google Scholar]
- Dressler, R.L. Recent advances in orchid phylogeny. Lindleyana 1986, 1, 5–20. [Google Scholar]
- Dressler, R.L. The vandoid orchids: A polyphyletic grade? Lindleyana 1989, 4, 89–93. [Google Scholar]
- Jakubska-Busse, A.; Proćków, J.; Górniak, M.; Gola, E.M. Is Epipactis pseudopurpurata distinct from E. purpurata (Orchidaceae)? Evidence from morphology, anatomy, DNA and pollination biology. Bot. J. Linn. Soc. 2012, 170, 243–256. [Google Scholar] [CrossRef][Green Version]
- Savelli, P.R.; Alessandrini, A. Epipactis flaminia Savelli et Alessandrini, sp. nov. (Orchidaceae) in the Apennines of Romagna. Webbia 1994, 49, 25–29. [Google Scholar] [CrossRef]
- Seiser, K. Eine neue Subspezies der Epipactis greuteri (Orchidaceae) am Fuβ der Rax (Niederösterreich). Neilrechia 2001, 1, 137–148. [Google Scholar]
- Species 2000 & ITIS Catalogue of Life, 2019 Annual Checklist. Available online: http://www.catalogueoflife.org/annual-checklist/2019/ (accessed on 15 May 2020).
- World Checklist of Selected Plant Families (WCSP). Available online: http://wcsp.science.kew.org/prepareChecklist.do;jsessionid=BD13BC5465862F5D74079A6ACC47EE0B.kppapp06-wcsp?checklist=selected_families%40%40135140520202307081 (accessed on 15 May 2020).
- Průša, D. Orchideje České Republiky; Albatros Media: Praha, Czech, 2019. [Google Scholar]
- Batoušek, P. Zwei neue Hybriden aus der Tschechischen Republik. J. Eur. Orch. 1997, 29, 641–652. [Google Scholar]
- Štěpánková, J.; Hejný, S. (Eds.) Květena České Republiky; 8 Academia: Praha, Czech Republic, 2011. [Google Scholar]
- Madesis, P.; Mastrogianni, A.; Pelteki, N.; Tsiftsis, S.; Drouzas, A. DNA barcoding of the Epipactis taxa native in Greece. In What Future for Orchids, Proceedings of the 18th European Orchid Council Conference and Exhibition, Paris, France, 24–25 March 2018; Prat, D., Ed.; SFO: Paris, France, 2018; pp. 84–85. [Google Scholar]
- Tranchida-Lombardo, V.; Cafasso, D.; Cristaudo, A.; Cozzolino, S. Phylogeographic patterns, genetic affinities and morphological differentiation between Epipactis helleborine and related lineages in a Mediterranean glacial refugium. Ann. Bot. 2011, 107, 427–436. [Google Scholar] [CrossRef]
- Chase, M.W.; Hills, H.G. Silica gel: An ideal material for field preservation of leaf samples for DNA studies. Taxon 1991, 40, 215–220. [Google Scholar] [CrossRef]
- Douzery, E.J.P.; Pridgeon, A.M.; Kores, P.; Linder, H.P.; Kurzweil, H.; Chase, M.W. Molecular phylogenetics of Diseae (Orchidaceae): A contribution from nuclear ribosomal ITS sequences. Am. J. Bot. 1999, 86, 887–899. [Google Scholar] [CrossRef]
- Shaw, J.; Lickey, E.B.; Beck, J.T.; Farmer, S.B.; Liu, W.; Miller, J.; Siripun, K.C.; Winder, C.T.; Schilling, E.E.; Small, R.L. The tortoise and the hare II: Relative utility of 21 noncoding chloroplast DNA sequences for phylogenetic analysis. Am. J. Bot. 2005, 92, 142–166. [Google Scholar] [CrossRef] [PubMed]
- Johnson, L.A.; Soltis, E. matK DNA sequence and phylogenetic reconstruction in Saxifragaceae s.s. Syst. Bot. 1994, 19, 143–156. [Google Scholar] [CrossRef]
- Shaw, J.; Lickey, E.B.; Schilling, E.E.; Small, R.L. Comparison of whole chloroplast genome sequences to choose noncoding regions for phylogenetic studies in angiosperms: The tortoise and the hare III. Am. J. Bot. 2007, 94, 275–288. [Google Scholar] [CrossRef]
- Bandelt, H.J.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characters | Variable Type |
|---|---|
| Stem number | 1 or a few clustered |
| Stem height (cm) | (12‒)20‒60(‒76) |
| Stem color | Dark green, greenish-grey, greenish-violet, sometimes washed violet at base |
| Stem indumentum | Dense, whitish pubescence |
| Leaves color | Dark green |
| Cauline leaves number | 4–8 |
| Bract length (long/width, cm) | (2‒)5.5 × (2‒)3.5‒5.5(‒7.5) |
| Upper leaf size (cm) | 4‒6 × 2‒4.5 |
| Median leaf sizes (cm) | 3.5‒11 × 2‒6.5 |
| Lower leaf size (cm) | 2‒5 × 1.3‒3.5 |
| Superior cauline leaf aspect | Bract-like |
| Median leaf shape | Lanceolate, spreading-erect, narrow, arching; lower 1–3 leaves short and rounded |
| Leaf margin edge | Regularly serrated, papillae of different height and length |
| Number of flowers | 5‒25(‒30) |
| Inflorescence length (cm) | Up to 18 cm |
| Inflorescence aspect | Elongated, occupying up to half the stem |
| Flower aspect | ± opening, bell-shaped, pendant |
| Sepal size (mm) | 9‒12 × 4‒5 |
| Sepal color | Pale green, greenish with a pink tint and apex bent backwards, pale pink |
| Sepal shape | Oval-acuminate, keeled |
| Petal size (mm) | Near equal as sepal |
| Petal color | Whitish with the center and apex yellowish-green or seldom light pink |
| Hypochile size (mm) | 4‒5 × 4‒5 |
| Hypochile color outside | Greenish-white |
| Hypochile color inside | Yellowish-green, olive green to reddish-brown and shiny |
| Hypochile aspect | Nectariferous |
| Hypochile-epichil connection | “U”-shaped |
| Epichile size (mm) | 3‒4.5 × 4‒5 |
| Epichile color | White, greenish or pinkish |
| Epichile bosses | 2 attenuated |
| Labellum size/aspect | 7‒9 mm long, × 4‒5 mm, margins sometimes undulate |
| Ovary size (mm) | 6‒15 × 3‒6 |
| Ovary shape | Peer-shaped, narrow |
| Ovary color | Yellowish-green, |
| Ovary indumentum | Glabrescent |
| Ovary pedicel length | Very elongated, (4‒)4.5–12 mm |
| Ovary pedicel color | Green at base, washed yellowish |
| Gynostemium size (mm) | Ca. 3.5‒5.5 × 1.5‒3.5 |
| Viscidium | Absent |
| Anther | Sessile, projecting beyond rostellum |
| Rostellum | Presence |
| Clinandrium | Strongly reduced |
| Stigma | Orientated downwards or towards the anther |
| Pollinia Flowering period | Friable and then crumbling, disintegrating onto the stigma Late June to early September |
| Haplotypes | trnS-trnG | 5’trnK-matK | rpl16 | Specimens | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 47 | 311–324 | 486 | 519 | 193 | 395 | 483 | 562 | 948 | 366–375 | ||
| H6 | T | - | C | A9 | C | G | A9 | T8 | C | - | E. atrorubens |
| H5 | G | +A | A | A10 | C | A | A9 | T11 | C | + | E. helleborine |
| H4 | G | +C | A | A10 | C | A | A9 | T12 | C | + | E. helleborine E. purpurata E. greuteri |
| H3 | G | - | A | A9 | C | A | A11 | T10 | T | - | E. albensis |
| H2 | G | - | A | A9 | A | A | A11 | T11 | C | - | E. purpurata |
| H1 | G | - | A | A9 | C | A | A11 | T11 | C | - | E. helleborine |
| Classification of Epipactis greuteri | ||
| A species complex (aggregate) (distinguished only by Delforge [1]) | Epipactis leptochila agg. | |
| Species | Epipactis greuteri H.Baumann & Künkele | |
| Nomenclature Investigation | ||
| Infraspecific Taxa Previously Classified to E. greuteri | Synonyms | Current Accepted Names |
| Epipactis greuteri subsp. preinensis Seiser | Epipactis preinensis (Seiser) Landolt | Epipactis flaminia P.R.Savelli & Aless.* |
| Epipactis greuteri var. preinensis (Seiser) P.Delforge | ||
| Epipactis greuteri subsp. flaminia (P.R.Savelli & Aless.) H.Baumann, Künkele & R.Lorenz | – | Epipactis flaminia P.R.Savelli & Aless. |
| Epipactis greuteri var. flaminia (P.R.Savelli & Aless.) Kreutz | – | |
| Epipactis greuteri var. aspromontana (Bartolo, Pulv. & Robatsch) P.Delforge | Epipactis aspromontana Bartolo, Pulv. & Robatsch Epipactis helleborine subsp. aspromontana (Bartolo, Pulv. & Robatsch) H.Baumann & R.Lorenz | Epipactis leptochila subsp. aspromontana (Bartolo, Pulv. & Robatsch) Kreutz |
| Epipactis greuteri subsp. sancti-bruni Bongiorni, De Vivo, Fori & Pisani | – | Epipactis greuteri H.Baumann & Künkele |
| Amplified Region | Initial Denaturation, °C/time | Denaturation, °C/time | Annealing, °C/time | Elongation, °C/time | Final Elongation, °C/time | No. of Cycles |
|---|---|---|---|---|---|---|
| ITS | 94/3 min | 94/45 s | 52/45 s | 72/60 s | 72/5 min | 29 |
| rpL16 | 80/5 min | 95/60 s | 50/60 s | 65/4 min | 65/7 min | 30 |
| matK-5’trnk | 80/5 min | 94/60 s | 50/60 s | 75/2 min | 72/7 min | 28 |
| trnS-trnG-trnG | 80/5 min | 95/60 s | 50/60 s | 65/4 min | 65/7 min | 30 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jakubska-Busse, A.; Żołubak, E.; Górniak, M.; Łobas, Z.; Tsiftsis, S.; Steiu, C. A Revision of the Taxonomy and Identification of Epipactis greuteri (Orchidaceae, Neottieae). Plants 2020, 9, 783. https://doi.org/10.3390/plants9060783
Jakubska-Busse A, Żołubak E, Górniak M, Łobas Z, Tsiftsis S, Steiu C. A Revision of the Taxonomy and Identification of Epipactis greuteri (Orchidaceae, Neottieae). Plants. 2020; 9(6):783. https://doi.org/10.3390/plants9060783
Chicago/Turabian StyleJakubska-Busse, Anna, Elżbieta Żołubak, Marcin Górniak, Zbigniew Łobas, Spyros Tsiftsis, and Corina Steiu. 2020. "A Revision of the Taxonomy and Identification of Epipactis greuteri (Orchidaceae, Neottieae)" Plants 9, no. 6: 783. https://doi.org/10.3390/plants9060783
APA StyleJakubska-Busse, A., Żołubak, E., Górniak, M., Łobas, Z., Tsiftsis, S., & Steiu, C. (2020). A Revision of the Taxonomy and Identification of Epipactis greuteri (Orchidaceae, Neottieae). Plants, 9(6), 783. https://doi.org/10.3390/plants9060783