Nutritive Value Response of Native Warm-Season Forage Grasses to Harvest Intervals and Durations in Mixed Stands



Abstract
:1. Introduction
2. Results and Discussion
2.1. Chemical Composition of Forage Dry Matter
2.1.1. Crude Protein Concentration

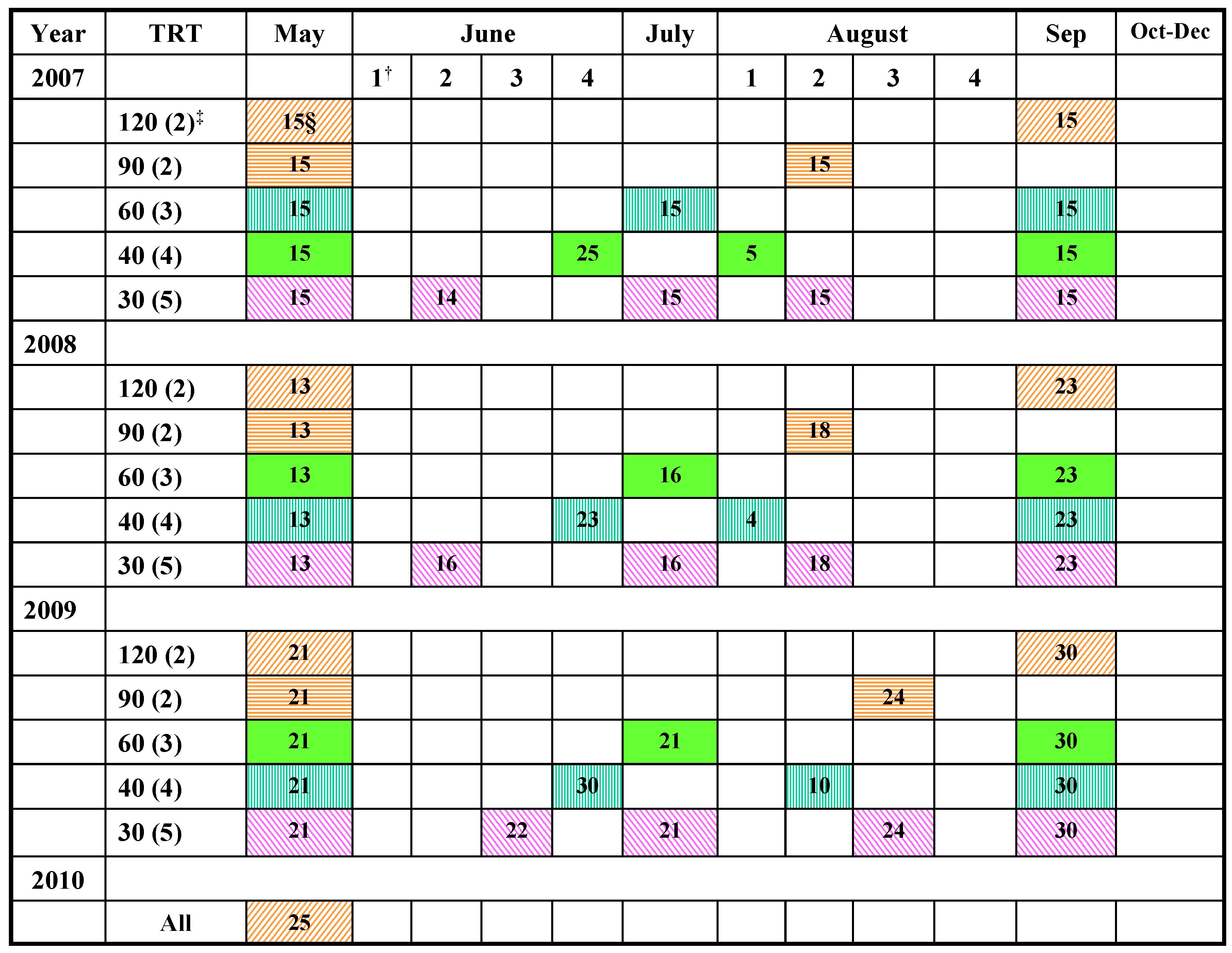
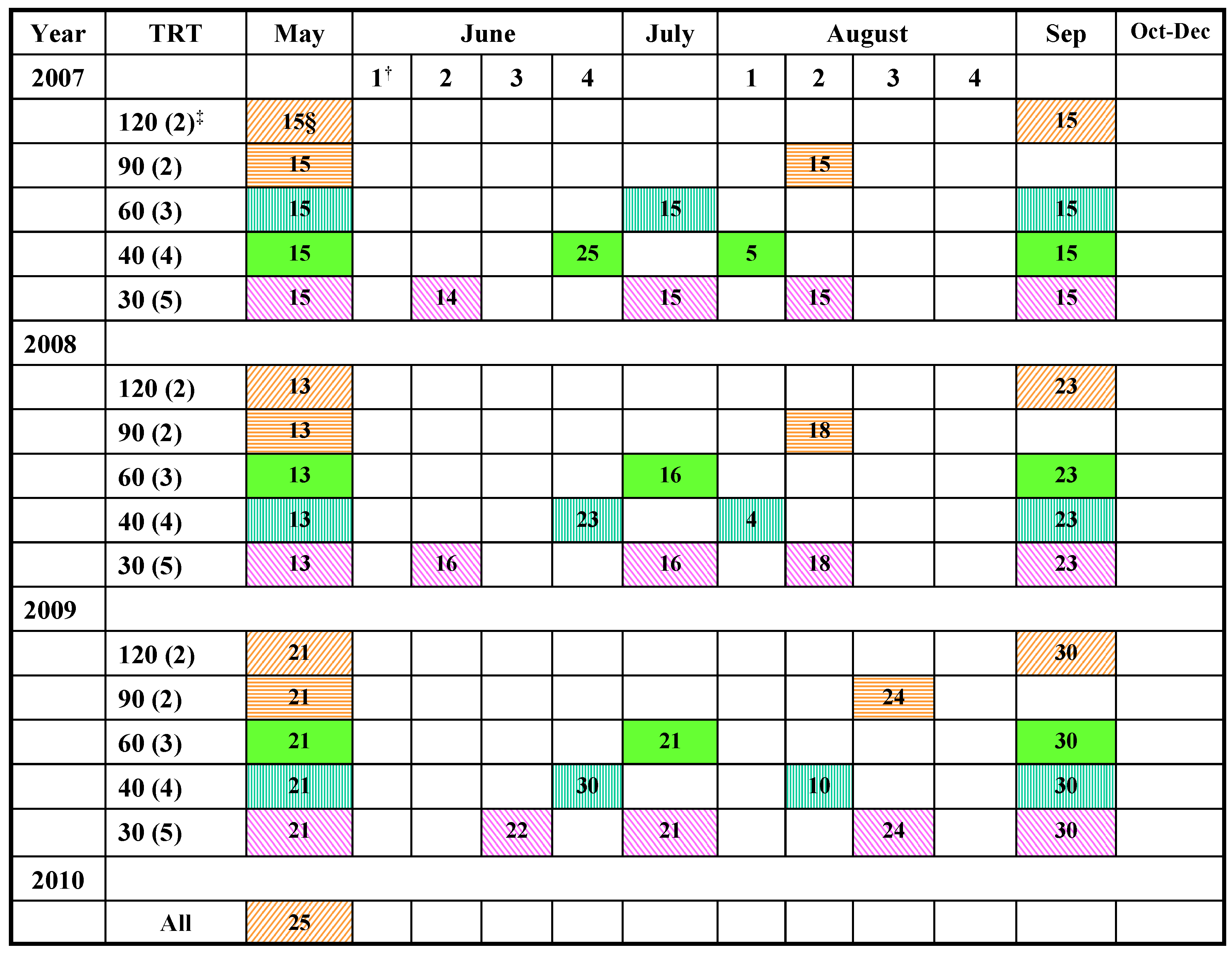{kind=link}
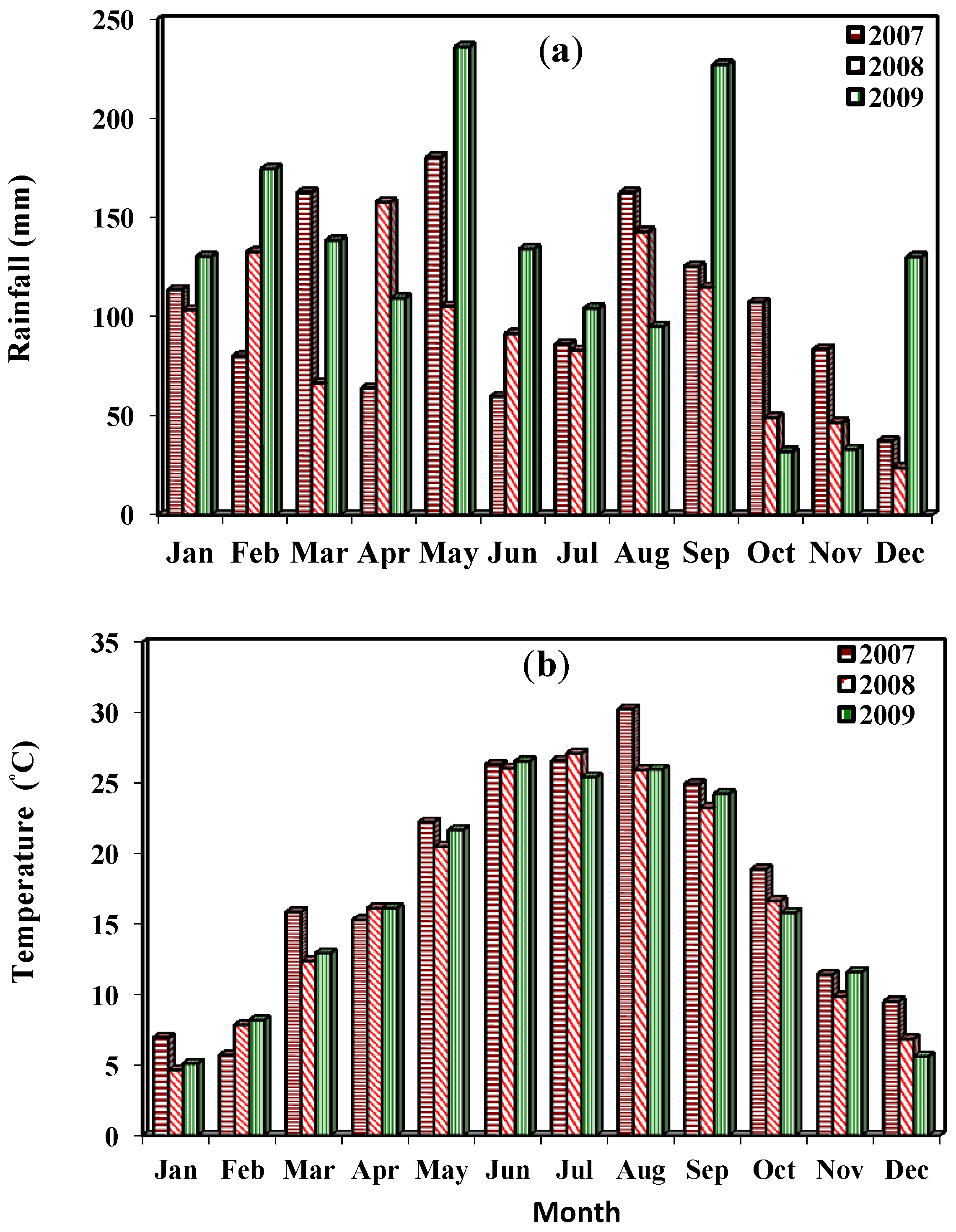{kind=link}
{kind=link}
| Frequency | Harvested regrowth | p > Fα § | |||
|---|---|---|---|---|---|
| 2nd | 3rd | 4th | 5th | ||
| g·kg−1 DM | |||||
| 120 (2) ‡ | 42 E | ||||
| 90 (2) | 44 DE | ||||
| 60 (3) | 54 C | 52 CD | 0.54 | ||
| 40 (4) | 69a # AB | 64a B | 52b CD | <0.01 | |
| 30 (5) | 74a A | 67ab AB | 63b B | 47c CDE | <0.01 |
2.1.2. Across-Season Crude Protein Fluctuations
2.1.3. Forage Fiber Concentration
2.1.3.1. Neutral Detergent Fiber (NDF) Concentration
| Interval (days) | June-September 2008 | June-September 2009 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 2nd | 3rd | 4th | 5th | p > Fα § | 2nd | 3rd | 4th | 5th | p > Fα | |
| g·kg−1 DM | g·kg−1 DM | |||||||||
| NDF | ||||||||||
| 120 (2) | 670 ABC # | - | - | - | - | 710 A | - | - | - | - |
| 90 (2) | 650 C | - | - | - | - | 660 B | - | - | - | - |
| 60 (3) | 680a AB | 650b | - | - | <0.02 | 640b BC | 680a | - | - | <0.01 |
| 40 (4) | 660 BC | 650 | 630 | - | 0.33 | 630b BC | 660ab | 670a | - | <0.02 |
| 30 (5) | 690 A | 670 | 670 | 670 | 0.08 | 620b C | 670a | 670a | 660a | <0.03 |
| ADF | ||||||||||
| 120 (2) | 360 | - | - | - | - | 400 A | - | - | - | - |
| 90 (2) | 360 | - | - | - | - | 360 B | - | - | - | - |
| 60 (3) | 370 | 360 | - | - | 0.41 | 340b BC | 370a | - | - | <0.01 |
| 40 (4) | 350 | 350 | 350 | - | 0.90 | 330c C | 350b | 370a | - | <0.01 |
| 30 (5) | 380 | 360 | 380 | 370 | 0.13 | 340b BC | 370a | 330b | 380a | <0.01 |
| ADL | ||||||||||
| 120 (2) | 54 A | - | - | - | - | 52 A | - | - | - | - |
| 90 (2) | 53 A | - | - | - | - | 47 AB | - | - | - | - |
| 60 (3) | 51 AB | 58 | - | - | 0.19 | 37 C | 43 | - | - | 0.14 |
| 40 (4) | 37b C | 41b | 55a | - | <0.011 | 43a BC | 31b | 46a | - | <0.01 |
| 30 (5) | 42c BC | 48bc | 53b | 67a | <0.01 | 46 AB | 44 | 44 | 45 | 0.9 |
2.1.3.2. Acid Detergent Fiber (ADF) Concentration

2.1.3.3. Acid Detergent Lignin (ADL) Concentration
2.2. Forage in Vitro Digestibility
2.2.1. Harvest Interval and DM Digestibility
| Interval (days) | June–September 2008 | June–September 2009 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Harvest Timing | p > Fα § | Harvest Timing | p > Fα | |||||||
| 2nd | 3rd | 4th | 5th | 2nd | 3rd | 4th | 5th | |||
| g·kg−1 DM | g·kg−1 DM | |||||||||
| DM | ||||||||||
| 120 (2) | 420 | - | - | - | - | 450 D # | - | - | - | - |
| 90 (2) | 470 | - | - | - | - | 530 C | - | - | - | - |
| 60 (3) | 460 | 480 | - | - | 0.48 | 580a †† BC | 490b | - | - | <0.01 |
| 40 (4) | 490 | 510 | 470 | - | 0.26 | 650a A | 570b | 480c | - | <0.01 |
| 30 (5) | 440 | 440 | 430 | 430 | 0.87 | 630a AB | 540b | 640a | 500b | <0.01 |
| NDF | ||||||||||
| 120 (2) | 560 C | - | - | - | - | 550 D | - | - | - | - |
| 90 (2) | 600 BC | - | - | - | - | 650 C | - | - | - | - |
| 60 (3) | 620 AB | 620 | - | - | 0.93 | 680a BC | 600b | - | - | <0.01 |
| 40 (4) | 670a A | 650ab | 610c | - | 0.02 | 740a A | 670b | 590c | - | <0.01 |
| 30 (5) | 620 AB | 620 | 590 | 570 | 0.11 | 720a AB | 650b | 740a | 600b | <0.01 |
2.2.2. Harvest Timing and DM Digestibility
| Interval (days) | LSR | SLA | ||||||
|---|---|---|---|---|---|---|---|---|
| 2008 | 2009 | 2008 | 2009 | |||||
| Y108 ‡ | Y207 | Y109 | Y208 | Y108 | Y207 | Y109 | Y208 | |
| cm−2·g−1 | ||||||||
| Indiangrass | ||||||||
| Control | - | - | 0.4b § | 0.4c | - | - | 98d | 98d |
| 120 (2) | 1.6 | 1.9 | 0.7b | 0.8bc | 109c | 102b | 101d | 110d |
| 90 (2) | 1.8 | 1.3 | 0.8b B | 1.4ab A | 107c | 100b | 108d | 110d |
| 60 (3) | 1.7 | 1.7 | 1.5a | 1.4ab | 112c | 117b | 227c | 243c |
| 40 (4) | 2.1 | 2.1 | 1.4a B | 1.8a A | 137b | 150a | 403b | 368b |
| 30 (5) | 1.8 | 2.0 | 1.3a B | 1.9a A | 152a | 154a | 567a A | 529a B |
| p > Fα# | 0.57 | 0.13 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| Little bluestem | ||||||||
| Control | - | - | 0.4b | 0.4c | - | - | 118d | 118d |
| 120 (2) | 1.2 | 1.0 | 0.6b | 0.7bc | 132b | 134c | 119d | 128d |
| 90 (2) | 1.3 | 2.1 | 0.8b B | 1.2ab A | 133b | 136c | 131d | 156d |
| 60 (3) | 1.5 | 1.6 | 1.2a B | 1.5a A | 162a | 156bc | 284c | 306c |
| 40 (4) | 1.7B | 1.9A | 1.3a | 1.2ab | 168a | 166ab | 475ab | 470b |
| 30 (5) | 1.9 | 2.3 | 1.4a | 1.5a | 162a | 194a | 732a A | 674a B |
| p > Fα | 0.14 | 0.21 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
2.2.3. Harvest Interval on NDF Digestibility
2.2.4. Harvest Timing on NDF Digestibility
2.3. Species Tiller Leafiness
2.3.1. Tiller Leaf-to-Stem Ratio
2.3.2. Tiller Specific Leaf Area
3. Experimental Section
3.1. Study Location and Field Layout
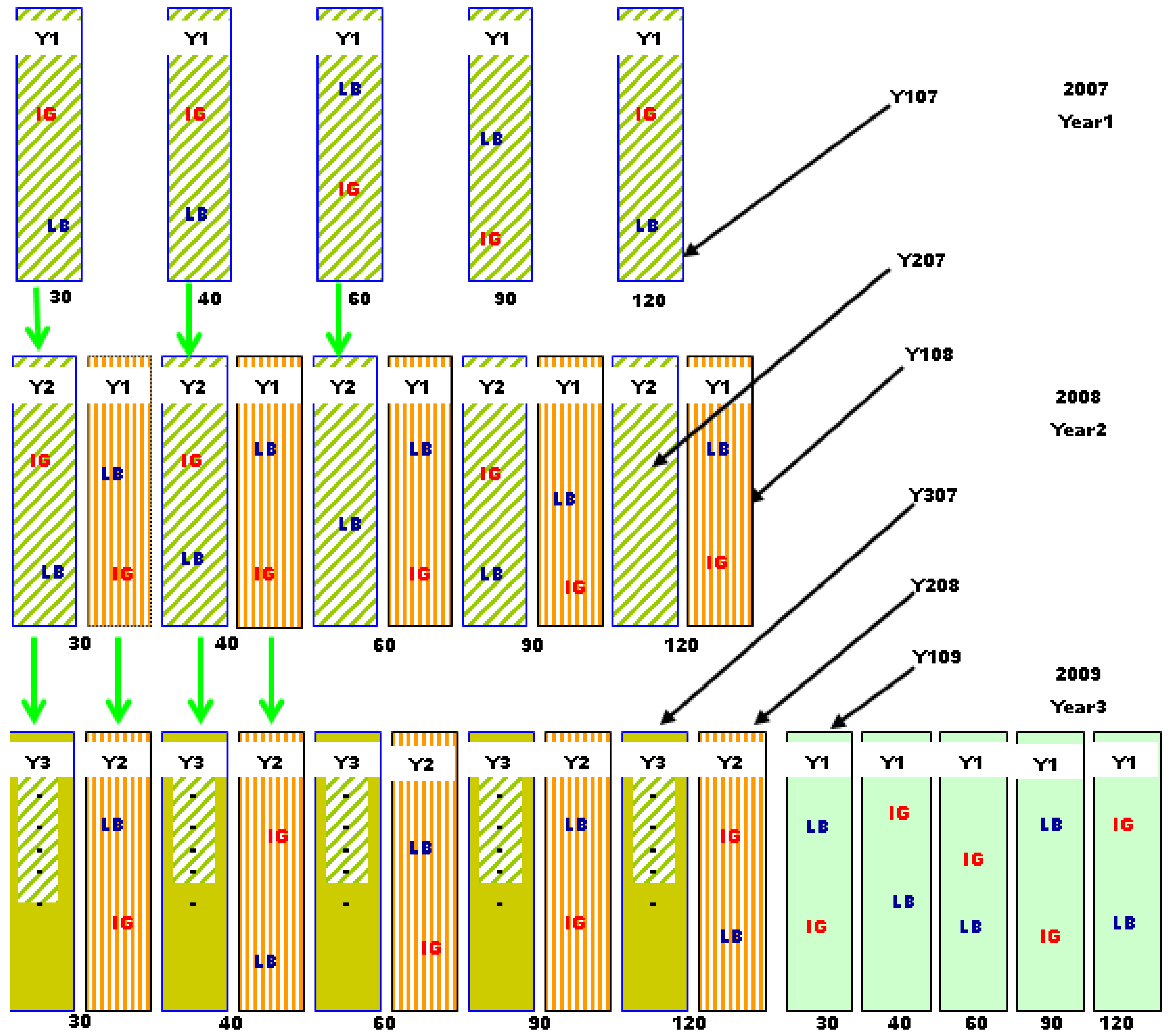
3.2. Harvesting and Forage Sampling
3.3. Species Morphological Assessment
3.4. Forage Nutritive Value Assessment
3.5. Data Analyses
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Shea, A.B. The Return of Native Grasses to Tennessee. 1999. Available online: http://www.tn.gov/environment/conservationist/archive/grass.htm (accessed on 12 August 2009). [Google Scholar]
- Jones, J.; Coggin, D.S.; Cummins, J.L.; Hill, J. Restoring and Managing Native Prairies: A Handbook for Mississippi Landowners; Wildlife Mississippi: Starkville, MS, USA, 2007. [Google Scholar]
- Griffin, J.L.; Jung, G.A. Leaf and stem forage quality of big bluestem and switchgrass. Agron. J. 1983, 75, 723–726. [Google Scholar] [CrossRef]
- Mitchell, R.B.; Vogel, K.P.; Klopfenstein, T.J.; Anderson, B.E.; Masters, R.A. Grazing evaluation of big bluestems bred for improved forage yield and digestibility. Crop Sci. 2005, 45, 2288–2292. [Google Scholar] [CrossRef]
- Forwood, J.R.; Magai, M.M. Clipping frequency and intensity effects on big bluestem yield, quality, and persistence. J. Range Manag. 1992, 45, 554–559. [Google Scholar] [CrossRef]
- Al Haj Khaled, R.; Duru, M.; Decruyenaere, V.; Journey, C.; Cruz, P. Using leaf traits to rank native grasses according to their nutritive value. Rangel. Ecol. Manag. 2006, 59, 648–654. [Google Scholar] [CrossRef]
- Cherney, D.J.R.; Cherney, J.H.; Lucey, R.F. In vitro digestion kinetics and quality of perennial grasses as influenced by forage maturity. J. Dairy Sci. 1993, 76, 790–797. [Google Scholar] [CrossRef]
- Twidwell, E.K.; Johnson, K.D.; Cherney, J.H.; Volenec, J.J. Forage quality and digestion kinetics of switchgrass herbage and morphological components. Crop Sci. 1988, 28, 778–782. [Google Scholar] [CrossRef]
- Linn, J.G.; Martin, N.P. Forage Quality Test and Interpretations; AG-FO-2637; Minnesota Extension Service, University of Minnesota: St. Paul, MN, USA, 1999. [Google Scholar]
- Wilson, J.R.; Hatfield, R.D. Structural and chemical changes of cell wall types during stem development: Consequences for fibre degradation by rumen microflora. Aust. J. Agric. Res. 1997, 48, 165–180. [Google Scholar] [CrossRef]
- Vona, L.C.; Jung, G.A.; Reid, R.L.; Sharp, W.C. Nutritive value of warm-season grass hays for beef cattle and sheep; digestibility, intake and mineral utilization. J. Anim. Sci. 1984, 59, 1582–1593. [Google Scholar]
- Buxton, D.R.; Redfearn, D.D. Plant limitations to fiber digestion and utilization. J. Nutr. 1997, 127, 814S–818S. [Google Scholar]
- Difante, G.S.; Nascimento Júnior, D.; Silva, S.C.; Euclides, V.P.B.; Zanine, A.M.; Adese, B. Tillering dynamics of marandu palisadegrass submitted to two cutting heights and three cutting intervals. Rev. Bras. Zootec. 2008, 37, 189–196. [Google Scholar] [CrossRef]
- Manske, L.L.; Ske, L.L. General Description of Grass Growth and Development and Defoliation Resistance Mechanisms; Range Management Report DREC 98–1022, USA; NDSU Dickinson Research Extension Center: Dickinson, ND, USA, 1998. [Google Scholar]
- Hannaway, D.B.; Hannaway, K.J.; Sohn, P.; Griffith, S.; Wycoff, H. Grass Growth and Regrowth for Improved Management. 2000. Available online: http://www.fsl.orst.edu/forages/projects/regrowth/main.cfm?PageID=7 (accessed on 30 August 2008).
- National Research Council. Nutrient Requirement of Beef Cattle, 7th revised ed.; NRC, National Academy Press: Washington, DC, USA, 1996. [Google Scholar]
- Nordheim-Viken, H.; Jørgensen, M.; Volden, H. Effects of maturity stage, temperature and photoperiod on growth and nutritive value of timothy (Phleum pratense L.). Anim. Feed Sci. Technol. 2009, 152, 204–218. [Google Scholar] [CrossRef]
- Beaty, E.R.; Engel, J.L. Forage quality measurements and forage research: A review, critique and interpretation. J. Range Manag. 1980, 33, 49–54. [Google Scholar]
- Sanderson, M.A.; Voigt, P.; Jones, R.M. Yield and quality of warm-season grasses in central Texas. J. Range Manag. 1999, 52, 145–150. [Google Scholar]
- Ball, D.M.; Hoveland, C.S.; Lacefield, G.D. Southern Forages, 1st ed.; Potash & Phosphate Inst. and the Foundation for Agronomic Research: Atlanta, GA, USA, 1991; p. 322. [Google Scholar]
- Briske, D.D. Developmental morphology and physiology of grasses. In Grazing Management: An Ecological Perspective; Heitschmidt, R.K., Sahlu, T., Eds.; Timber Press: Portland, OR, USA, 1991; pp. 85–108. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar]
- Weiss, W.P.; Eastridge, M.L.; Underwood, J.F. Forages for Dairy Cattle. 2010. Available online: http://ohioline.osu.edu/as-fact/0002.html (accessed on 20 January 2011).
- Reid, R.L.; Jung, G.A.; Thayne, W.V. Relationships between nutritive quality and fiber components of cool season and warm season forages: A retrospective study. J. Anim. Sci. 1988, 66, 1275–1291. [Google Scholar]
- Buxton, D.R. Quality-related characteristics of forages as influenced by plant environment and agronomic factors. Anim. Feed Sci. Technol. 1996, 59, 37–49. [Google Scholar] [CrossRef]
- Moore, K.J.; Jung, H.J.G. Lignin and fiber digestion. J. Range Manag. 2001, 54, 420–430. [Google Scholar] [CrossRef]
- Mulkey, V.R.; Owens, V.N.; Lee, D.K. Management of warm-season grass mixtures for biomass production in South Dakota USA. Bioresour. Technol. 2008, 99, 609–617. [Google Scholar] [CrossRef]
- Hendrickson, J.R.; Moser, L.E.; Moore, K.J.; Waller, S.S. Leaf nutritive value related to tiller development in warm-season grasses. J. Range Manag. 1997, 50, 116–122. [Google Scholar] [CrossRef]
- Hoffman, P.C.; Lundberg, K.M.; Bauman, L.M.; Shaver, R.D. The effect of maturity on NDF digestibility. Focus Forage 2003, 5. No. 15. [Google Scholar]
- Hoffman, P.C.; Lundberg, K.M.; Barman, L.M.; Shaver, R.D. NDF digestibility: Reference values for forages, byproducts and total mixed rations. Focus Forage 2006, 5, No. 17. [Google Scholar]
- Sbrissia, A.F.; Silva, S.C.; Matthew, C.; Carvalho, C.A.B.; Carnevalli, R.A.; Pinto, L.F.M.; Fagundes, J.L.; Pedreira, C.G.S. Tiller size/density compensation in grazed Tifton 85 bermudagrass swards. Pesqui. Agropecu. Bras. 2003, 38, 1459–1468. [Google Scholar]
- Association of Official Analytical Chemists. Official Methods of Analysis; Association of Official Analytical Chemist: Washington, DC, USA, 2001. [Google Scholar]
- ANKOM Technologies Method 3. In Vitro True Digestibility Using the DaisyII Inclubator. ANKOM Technologies -08/05. 2005. Available online: http://www.ankom.com/media/documents/IVDMD_0805_D200.pdf (accessed on 13 September 2010).
- SAS Institute. SAS Version 9.2, SAS Institute: Cary, NC, USA, 2007.
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Temu, V.W.; Rude, B.J.; Baldwin, B.S. Nutritive Value Response of Native Warm-Season Forage Grasses to Harvest Intervals and Durations in Mixed Stands. Plants 2014, 3, 266-283. https://doi.org/10.3390/plants3020266
Temu VW, Rude BJ, Baldwin BS. Nutritive Value Response of Native Warm-Season Forage Grasses to Harvest Intervals and Durations in Mixed Stands. Plants. 2014; 3(2):266-283. https://doi.org/10.3390/plants3020266
Chicago/Turabian StyleTemu, Vitalis W., Brian J. Rude, and Brian S. Baldwin. 2014. "Nutritive Value Response of Native Warm-Season Forage Grasses to Harvest Intervals and Durations in Mixed Stands" Plants 3, no. 2: 266-283. https://doi.org/10.3390/plants3020266
APA StyleTemu, V. W., Rude, B. J., & Baldwin, B. S. (2014). Nutritive Value Response of Native Warm-Season Forage Grasses to Harvest Intervals and Durations in Mixed Stands. Plants, 3(2), 266-283. https://doi.org/10.3390/plants3020266