
Photomorphogenic and Biochemical Effects of Radiation and Nitrate Availability on the Red Alga Plocamium cartilagineum

 , , , , and
, , , , and
Abstract
1. Introduction
2. Results
2.1. Photosynthetic Parameters
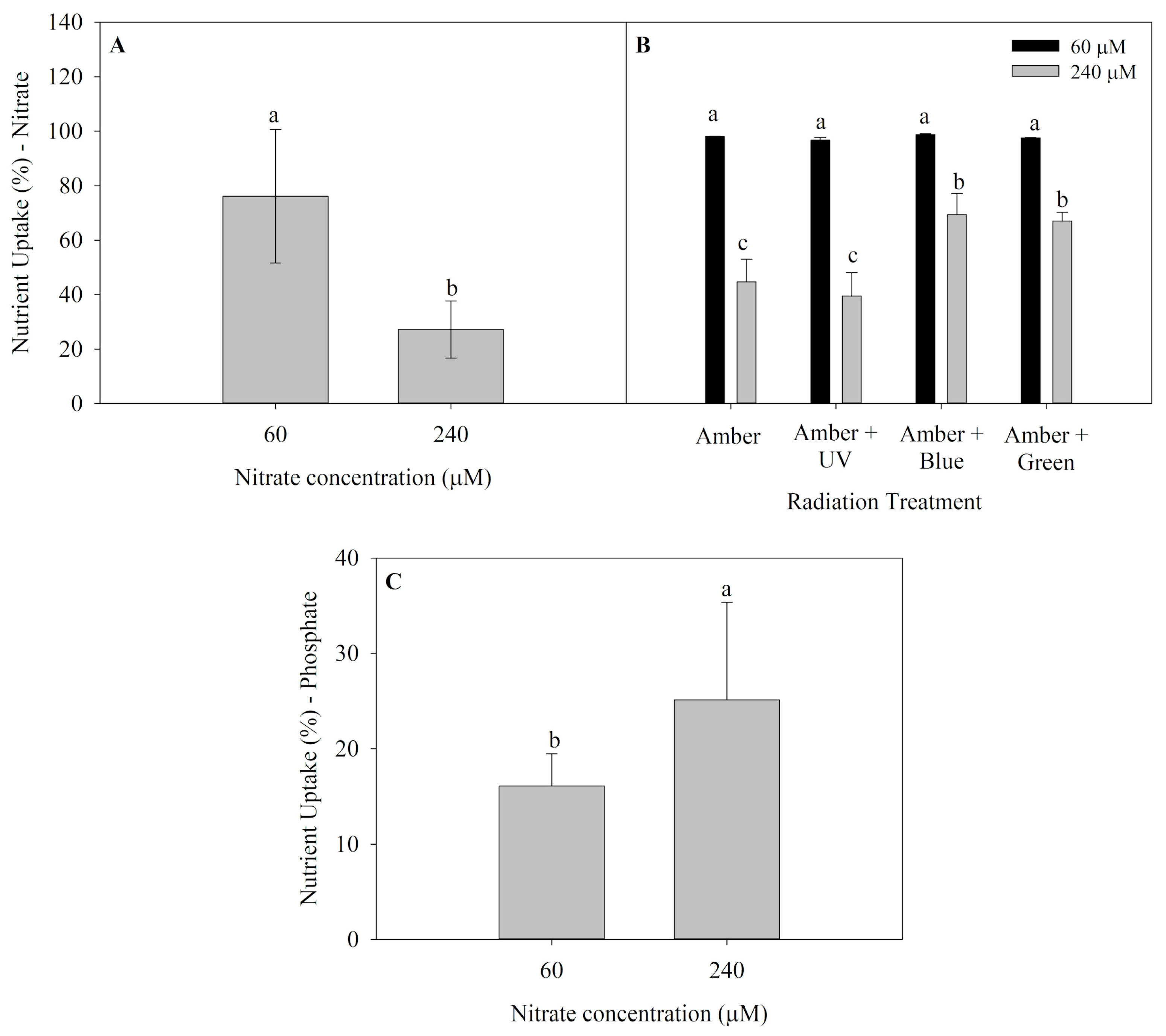
2.2. Nutrient Uptake
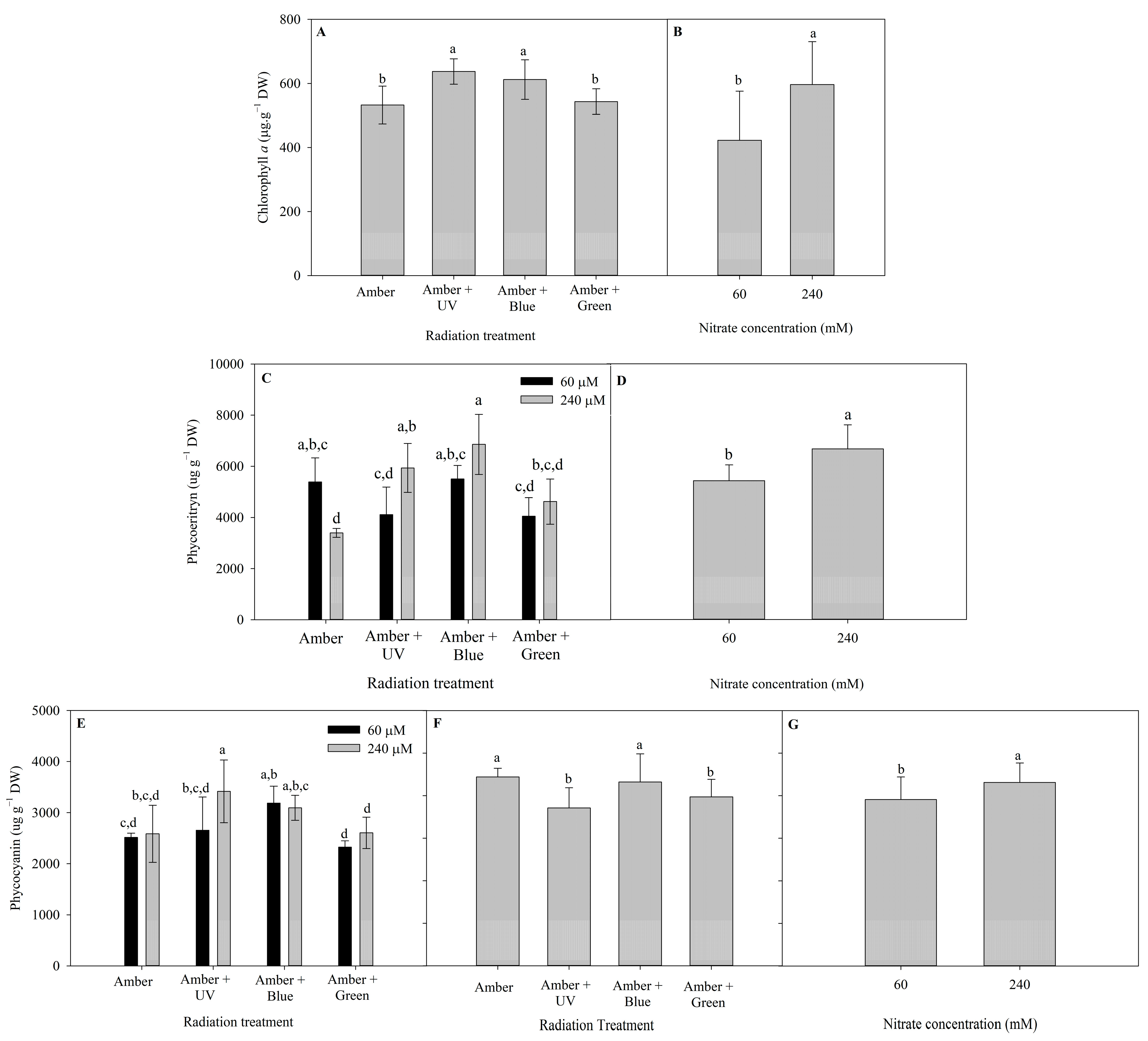
2.3. Photosynthetic Pigments
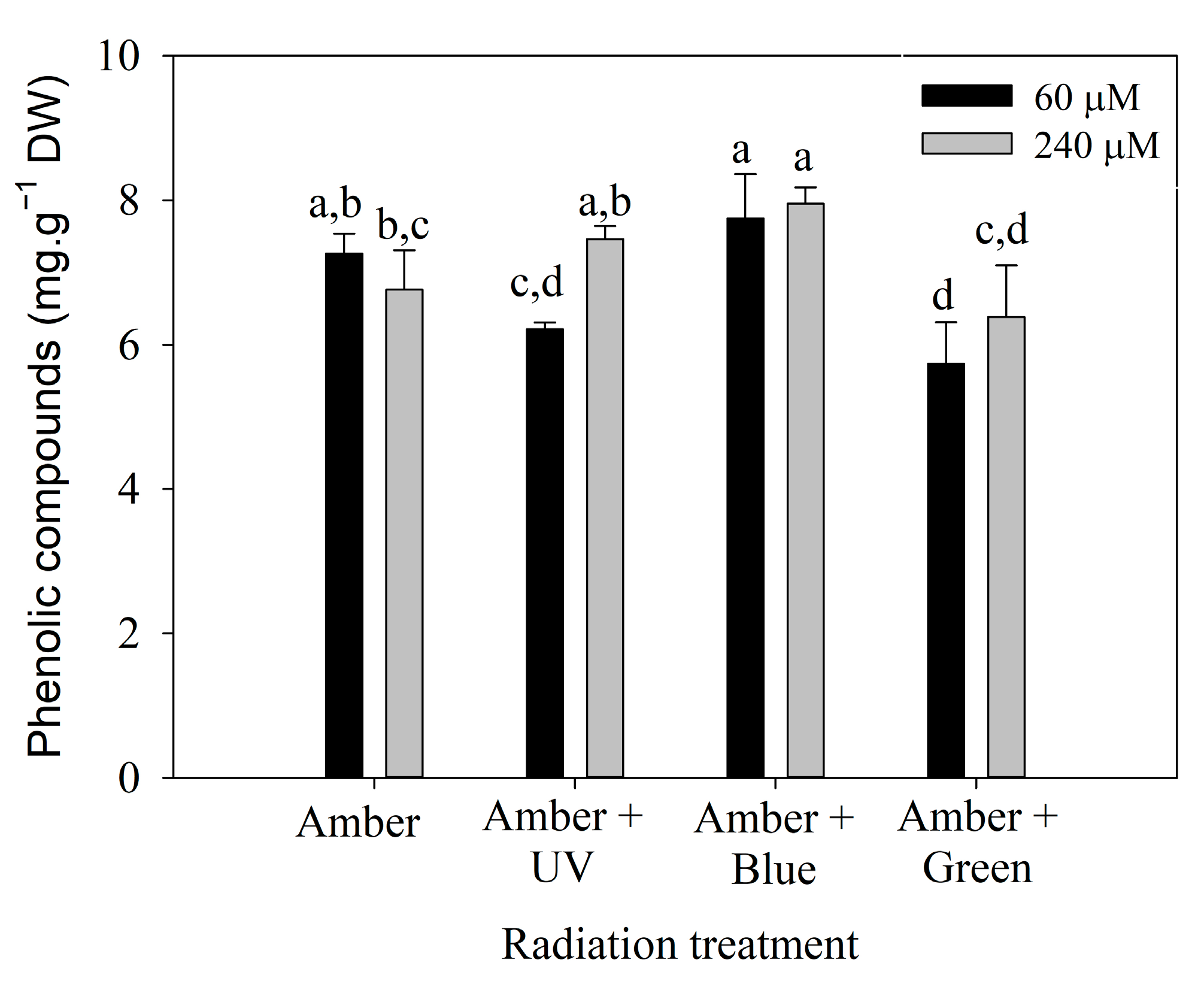
2.4. Phenolic Compounds
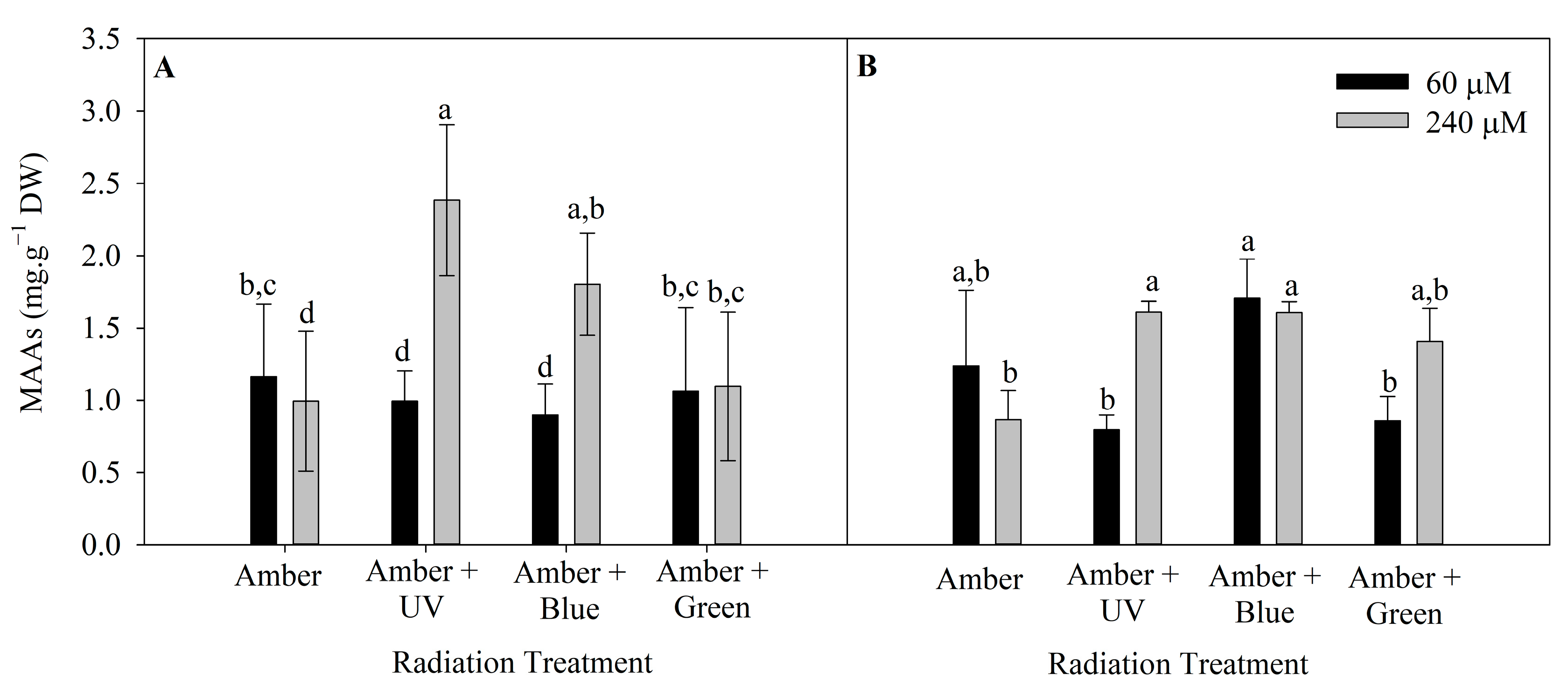
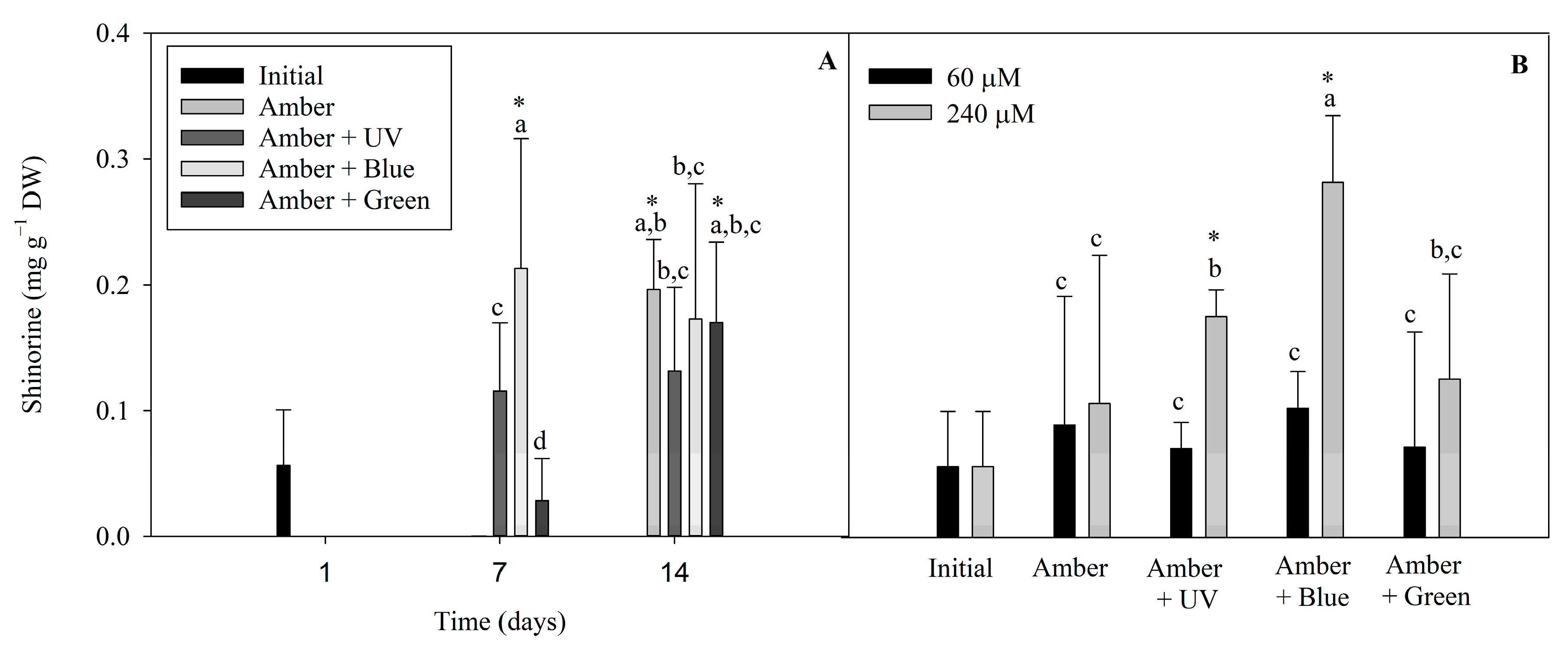
2.5. Mycosporine-like Amino Acids
3. Discussion
4. Materials and Methods
4.1. Biological Material and Acclimation
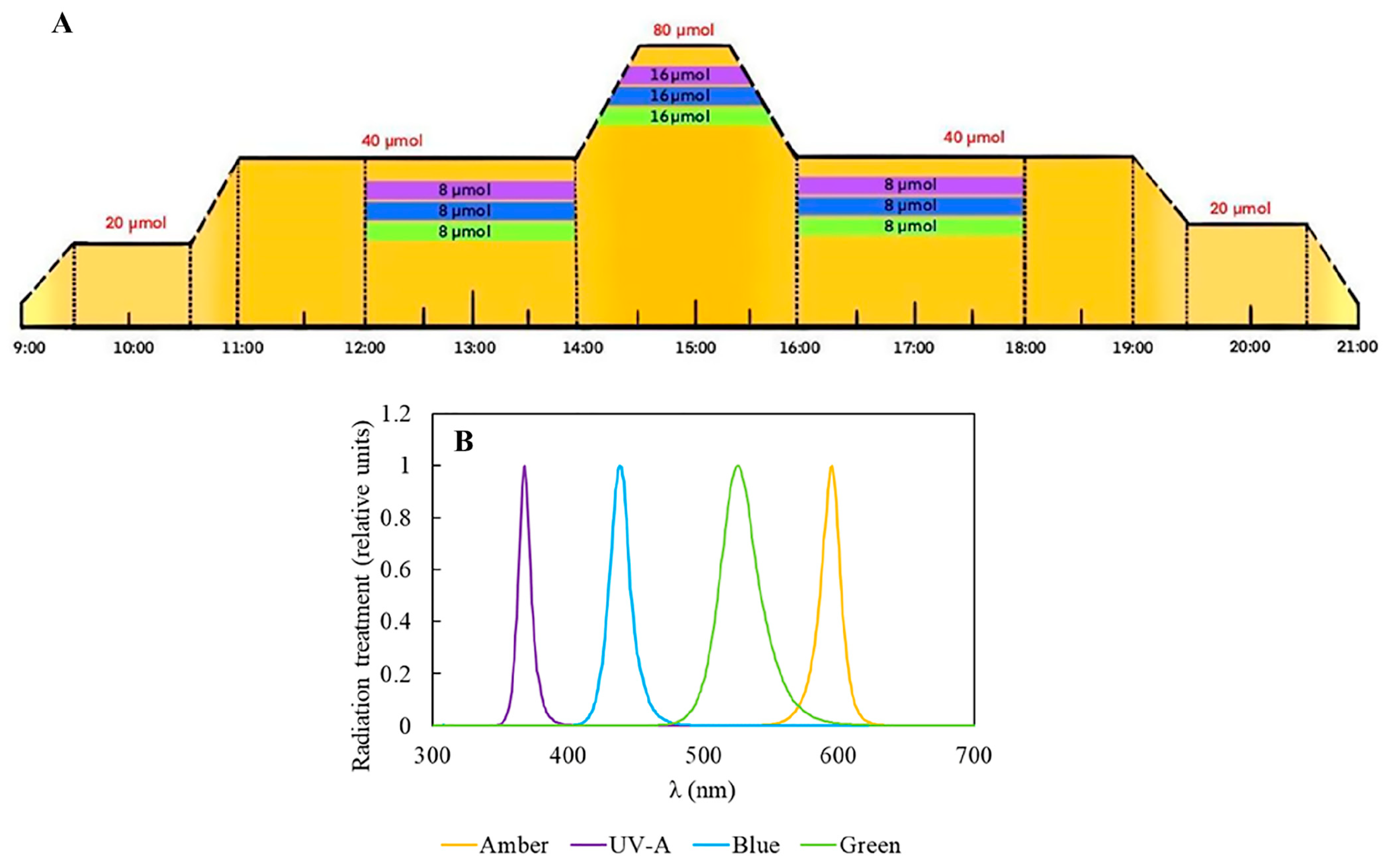
4.2. Experimental Design
4.3. Photosynthetic Performance
4.4. Nutrient Assimilation
4.5. Photosynthetic Pigments
4.6. Phenolic Compounds
4.7. Micosporine-like Amino Acids
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ANOVA | Analysis of variance |
| Chl a | Chlorophyll a |
| DW | Dry weight |
| ETR | Electron transport rate |
| MAA | Mycosporine-like amino acid |
| NU | Nutrient uptake |
| PC | Phycocyanin |
| PE | Phycoerythrin |
| UV-A | Ultraviolet A radiation |
References
- Cardona, T.; Shao, S.; Nixon, P.J. Enhancing Photosynthesis in Plants: The Light Reactions. Essays Biochem. 2018, 62, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Kianianmomeni, A.; Hallmann, A. Algal Photoreceptors: In Vivo Functions and Potential Applications. Planta 2014, 239, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Rüdiger, W.; López-Figueroa, F. Photoreceptors in Algae. Photochem. Photobiol. 1992, 55, 949–954. [Google Scholar] [CrossRef]
- Rockwell, N.C.; Duanmu, D.; Martin, S.S.; Bachy, C.; Price, D.C.; Bhattacharya, D.; Worden, A.Z.; Lagarias, J.C. Eukaryotic Algal Phytochromes Span the Visible Spectrum. Proc. Natl. Acad. Sci. USA 2014, 111, 3871–3876. [Google Scholar] [CrossRef]
- Hegemann, P.; Fuhrmann, M.; Kateriya, S. Algal Sensory Photoreceptors. J. Phycol. 2001, 37, 668–676. [Google Scholar]
- Sineshchekov, O.A.; Jung, K.H.; Spudich, J.L. Two Rhodopsins Mediate Phototaxis to Low- and High-Intensity Light in Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA 2002, 99, 8689–8694. [Google Scholar] [CrossRef]
- Luck, M.; Hegemann, P. The Two Parallel Photocycles of the Chlamydomonas Sensory Photoreceptor Histidine Kinase Rhodopsin 1. J. Plant Physiol. 2017, 217, 77–84. [Google Scholar] [CrossRef]
- Kato, S.; Ozasa, K.; Maeda, M.; Tanno, Y.; Tamaki, S.; Higuchi-Takeuchi, M.; Numata, K.; Kodama, Y.; Sato, M.; Toyooka, K.; et al. Carotenoids in the Eyespot Apparatus Are Required for Triggering Phototaxis in Euglena gracilis. Plant J. 2020, 101, 1091–1102. [Google Scholar] [CrossRef]
- López-Figueroa, F.; Lindemann, P.; Braslavsky, S.E.; Schaffner, K.; Schneider-Poetsch, H.A.W.; Rüdiger, W. Detection of a Phytochrome-like Protein in Macroalgae. Bot. Acta 1989, 102, 178–180. [Google Scholar]
- López-Figueroa, F.; Lindemann, P.; Braslavsky, S.E.; Schaffner, K.; Schneider-Poetsch, H.A.; Rüdiger, W. Detection of Some Conserved Domains in Phytochrome-like Proteins from Algae. J. Plant Physiol. 1990, 136, 484–487. [Google Scholar] [CrossRef]
- Hegemann, P. Algal Sensory Photoreceptors. Annu. Rev. Plant Biol. 2008, 59, 167–189. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Figueroa, F.; Perez, R.; Niell, F.X. Effects of Red and Far-Red Light Pulses on the Chlorophyll and Biliprotein Accumulation in the Red Alga Corallina Elongata. J. Photochem. Photobiol. B Biol. 1989, 4, 185–193. [Google Scholar] [CrossRef]
- López-Figueroa, F.; Niell, F.X. Effects of Light Quality on Chlorophyll and Biliprotein Accumulation in Seaweeds. Mar. Biol. 1990, 104, 321–327. [Google Scholar] [CrossRef]
- Allorent, G.; Petroutsos, D. Photoreceptor-Dependent Regulation of Photoprotection. Curr. Opin. Plant Biol. 2017, 37, 102–108. [Google Scholar] [CrossRef]
- Schneider, G.; Figueroa, F.L.; Vega, J.; Avilés, A.; Chaves, P.; Horta, P.A.; Korbee, N.; Bonomi-Barufi, J. Physiological and Biochemical Responses Driven by Different UV-Visible Radiation in: Osmundea pinnatifida (Hudson) Stackhouse (Rhodophyta). Photochem. Photobiol. Sci. 2020, 19, 1650–1664. [Google Scholar] [CrossRef]
- Vega, J.; Schneider, G.; Moreira, B.R.; Herrera, C.; Bonomi-Barufi, J.; Figueroa, F.L. Mycosporine-like Amino Acids from Red Macroalgae: UV-Photoprotectors with Potential Cosmeceutical Applications. Appl. Sci. 2021, 11, 5112. [Google Scholar] [CrossRef]
- Figueroa, F.L.; Bonomi-Barufi, J.; Celis-Plá, P.S.M.; Nitschke, U.; Arenas, F.; Connan, S.; Abreu, M.H.; Malta, E.J.; Conde-Álvarez, R.; Chow, F.; et al. Short-Term Effects of Increased CO2, Nitrate and Temperature on Photosynthetic Activity in Ulva rigida (Chlorophyta) Estimated by Different Pulse Amplitude Modulated Fluorometers and Oxygen Evolution. J. Exp. Bot. 2021, 72, 491–509. [Google Scholar] [CrossRef]
- Bonomi Barufi, J.; Teresa Mata, M.; Cabral Oliveira, M.; Figueroa, F.L.; Teresa Mata, A.; Figueroa, E.L. Nitrate Reduces the Negative Effect of UV Radiation on Photosynthesis and Pigmentation in Gracilaria tenuistipitata (Rhodophyta): The Photoprotection Role of Mycosporine-like Amino Acids. Phycologia 2012, 51, 636–648. [Google Scholar] [CrossRef]
- Figueroa, F.L.; Israel, A.; Neori, A.; Martínez, B.; Malta, E.J.; Put, A.; Inken, S.; Marquardt, R.; Abdala, R.; Korbee, N. Effect of Nutrient Supply on Photosynthesis and Pigmentation to Short-Term Stress (UV Radiation) in Gracilaria conferta (Rhodophyta). Mar. Pollut. Bull. 2010, 60, 1768–1778. [Google Scholar] [CrossRef]
- Figueroa, F.L. Effects of Light Quality on Nitrate Reductase and Glutamine Synthetase Activities in the Red Alga Porphyra Leucosticta Thur in Le Jol. and Other Macroalgae. Sci. Mar. 1996, 60, 163–170. [Google Scholar]
- Borthwick, H.A.; Hendricks, S.B.; Toole, E.H.; Toole, V.K. Action of Light on Lettuce-Seed Germination. Bot. Gaz. 1954, 115, 205–225. [Google Scholar] [CrossRef]
- Figueroa, F.L.; Aguilera, J.; Niell, F.X. Red and Blue Light Regulation of Growth and Photosynthetic Metabolism in Porphyra umbilicalis (Bangiales, Rhodophyta). Eur. J. Phycol. 1995, 30, 11–18. [Google Scholar] [CrossRef]
- Aguilera, J.; Gordillo, F.J.L.; Karsten, U.; Figueroa, F.L.; Niell, F.X. Light Quality Effect on Photosynthesis and Efficiency of Carbon Assimilation in the Red Alga Porphyra Leucosticta. J. Plant Physiol. 2000, 157, 86–92. [Google Scholar] [CrossRef]
- Korbee, N.; Figueroa, F.L.; Aguilera, J. Effect of Light Quality on the Accumulation of Photosynthetic Pigments, Proteins and Mycosporine-like Amino Acids in the Red Alga Porphyra leucosticta (Bangiales, Rhodophyta). J. Photochem. Photobiol. B Biol. 2005, 80, 71–78. [Google Scholar] [CrossRef]
- Bonomi Barufi, J.; Figueroa, F.L.; Plastino, E.M. Effects of Light Quality on Reproduction, Growth and Pigment Content of Gracilaria birdiae (Rhodophyta: Gracilariales). Sci. Mar. 2015, 79, 15–24. [Google Scholar] [CrossRef]
- Pagels, F.; Bonomi Barufi, J.; Vega, J.; Abdala-Díaz, R.; Vasconcelos, V.; Guedes, A.C.; Figueroa, F.L. Light Regulating Metabolic Responses of Cyanobium sp. (Cyanobacteria). Fundam. Appl. Limnol. 2020, 193, 285–297. [Google Scholar] [CrossRef]
- Schneider, G.; Figueroa, F.L.; Vega, J.; Avilés, A.; Horta, P.A.; Korbee, N.; Bonomi-Barufi, J. Effects of UV–Visible Radiation on Growth, Photosynthesis, Pigment Accumulation and UV-Absorbing Compounds in the Red Macroalga Gracilaria Cornea (Gracilariales, Rhodophyta). Algal Res. 2022, 64, 3–10. [Google Scholar] [CrossRef]
- Valentão, P.; Trindade, P.; Gomes, D.; Guedes De Pinho, P.; Mouga, T.; Andrade, P.B. Codium Tomentosum and Plocamium Cartilagineum: Chemistry and Antioxidant Potential. Food Chem. 2010, 119, 1359–1368. [Google Scholar] [CrossRef]
- Moreira, B.R.; Vega, J.; Sisa, A.D.A.; Bernal, J.S.B.; Abdala-Díaz, R.T.; Maraschin, M.; Figueroa, F.L.; Bonomi-Barufi, J. Antioxidant and Anti-Photoaging Properties of Red Marine Macroalgae: Screening of Bioactive Molecules for Cosmeceutical Applications. Algal Res. 2022, 68, 102893. [Google Scholar] [CrossRef]
- Moreira, B.R.; Fernandes, D.R.P.; Vega, J.; Álvarez-Gómez, F.; Varela, P.C.; Bonomi-Barufi, J.; Figueroa, F.L. Daily Variations on Photosynthesis, Pigmentation and Photoprotection of Plocamium cartilagineum (Plocamiaceae, Rhodophyta). Phycologia 2024, 63, 399–410. [Google Scholar] [CrossRef]
- Harden, E.A.; Falshaw, R.; Carnachan, S.M.; Kern, E.R.; Prichard, M.N. Virucidal Activity of Polysaccharide Extracts from Four Algal Species against Herpes Simplex Virus. Antivir. Res. 2009, 83, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Knott, M.G. A Review of Secondary Metabolites Isolated from Plocamium Species Worldwide. Int. Sci. Technol. J. Namibia 2016, 6, 75–93. [Google Scholar]
- Sabry, O.M.M.; Goeger, D.E.; Valeriote, F.A.; Gerwick, W.H. Cytotoxic Halogenated Monoterpenes from Plocamium Cartilagineum. Nat. Prod. Res. 2017, 31, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Häder, D.P.; Lebert, M.; Mercado, J.; Aguilera, J.; Salles, S.; Flores-Moya, A.; Jiménez, C.; Figueroa, F.L. Photosynthetic Oxygen Production and PAM Fluorescence in the Brown Alga Padina pavonica Measured in the Field under Solar Radiation. Mar. Biol. 1996, 127, 61–66. [Google Scholar]
- Flores-Moya, A.; Hanelt, D.; Figueroa, F.L.; Altamirano, M.; Viñegla, B.; Salles, S. Involvement of Solar UV-B Radiation in Recovery of Inhibited Photosynthesis in the Brown Alga Dictyota dichotoma (Hudson) Lamouroux. J. Photochem. Photobiol. B Biol. 1999, 49, 129–135. [Google Scholar] [CrossRef]
- Figueroa, F.L.; Escassi, L.; Pérez-Rodríguez, E.; Korbee, N.; Giles, A.D.; Johnsen, G. Effects of Short-Term Irradiation on Photoinhibition and Accumulation of Mycosporine-like Amino Acids in Sun and Shade Species of the Red Algal Genus Porphyra. J. Photochem. Photobiol. B Biol. 2003, 69, 21–30. [Google Scholar] [CrossRef]
- Thomas, B.; Dickinson, H.G. Evidence for Two Photoreceptors Controlling Growth in De-Etiolated Seedlings. Planta 1979, 146, 545–550. [Google Scholar]
- Castro-Varela, P.A.; Celis-Plá, P.S.M.; Abdala-Díaz, R.; Figueroa, F.L. Photobiological Effects on Biochemical Composition in Porphyridium cruentum (Rhodophyta) with a Biotechnological Application. Photochem. Photobiol. 2021, 97, 1032–1042. [Google Scholar] [CrossRef]
- Ghedifa, A.B.; Vega, J.; Korbee, N.; Mensi, F.; Felix, F.L.; Sadok, S. Effects of Light Quality on the Photosynthetic Activity and Biochemical Composition of Gracilaria gracilis (Rhodophyta). J. Appl. Phycol. 2021, 33, 3413–3425. [Google Scholar] [CrossRef]
- Wu, B.S.; Mansoori, M.; Trumpler, K.; Addo, P.W.; MacPherson, S.; Lefsrud, M. Effect of Amber (595 Nm) Light Supplemented with Narrow Blue (430 Nm) Light on Tomato Biomass. Plants 2023, 12, 2457. [Google Scholar] [CrossRef]
- López-Figueroa, F. Red, Green and Blue Light Photoreceptors Controlling Chlorophyll a, Biliprotein and Total Protein Synthesis in the Red Alga Chondrus crispus. Br. Phycol. Soc. 1991, 26, 383–393. [Google Scholar] [CrossRef]
- Petroutsos, D.; Tokutsu, R.; Maruyama, S.; Flori, S.; Greiner, A.; Magneschi, L.; Cusant, L.; Kottke, T.; Mittag, M.; Hegemann, P.; et al. A Blue-Light Photoreceptor Mediates the Feedback Regulation of Photosynthesis. Nature 2016, 537, 563–566. [Google Scholar] [CrossRef] [PubMed]
- Segovia, M.; Gordillo, F.J.L.; Schaap, P.; Figueroa, F.L. Light Regulation of Cyclic-AMP Levels in the Red Macroalga Porphyra leucosticta. J. Photochem. Photobiol. B Biol. 2001, 64, 69–74. [Google Scholar] [CrossRef]
- Segovia, M.; Gordillo, F.J.L.; Figueroa, F.L. Cyclic-AMP Levels in the Lichen Evernia prunastri Are Modulated by Light Quantity and Quality. J. Photochem. Photobiol. B Biol. 2003, 70, 145–151. [Google Scholar] [CrossRef]
- Navarro, N.P.; Mansilla, A.; Figueroa, F.L.; Korbee, N.; Jofre, J.; Plastino, E. Short-Term Effects of Solar UV Radiation and NO3− Supply on the Accumulation of Mycosporine-like Amino Acids in Pyropia columbina (Bangiales, Rhodophyta) under Spring Ozone Depletion in the Sub-Antarctic Region, Chile. Bot. Mar. 2014, 57, 9–20. [Google Scholar] [CrossRef]
- Yu, D.; Yin, J.; Wang, Y.; Lu, A.; He, Y.; Shen, S. Nitrogen Assimilation-Associated Enzymes and Nitrogen Use Efficiency of Pyropia yezoensis (Rhodophyta) in Nitrate-Sufficient Conditions. Algal Res. 2022, 64, 102682. [Google Scholar] [CrossRef]
- López-Figueroa, F.; Rüdiger, W. Stimulation of Nitrate Net Uptake and Reduction by Red and Blue Light and Reversion by Far-Red Light in the Green Alga Ulva rigida. J. Phycol. 1991, 27, 389–394. [Google Scholar] [CrossRef]
- Peinado, N.K.; Abdala Díaz, R.T.; Figueroa, F.L.; Helbling, E.W. Ammonium and UV Radiation Stimulate the Accumulation of Mycosporine-like Amino Acids in Porphyra columbina (Rhodophyta) from Patagonia, Argentina. J. Phycol. 2004, 40, 248–259. [Google Scholar] [CrossRef]
- Kräbs, G.; Watanabe, M.; Wiencke, C. A Monochromatic Action Spectrum for the Photoinduction of the UV-Absorbing Mycosporine-like Amino Acid Shinorine in the Red Alga Chondrus crispus. Photochem. Photobiol. 2004, 79, 515. [Google Scholar] [CrossRef]
- Navarro, N.P.; Figueroa, F.L.; Korbee, N.; Mansilla, A.; Plastino, E.M. Differential Responses of Tetrasporophytes and Gametophytes of Mazzaella laminarioides (Gigartinales, Rhodophyta) under Solar UV Radiation. J. Phycol. 2016, 52, 451–462. [Google Scholar] [CrossRef]
- Álvarez-Gómez, F.; Korbee, N.; Casas-Arrojo, V.; Abdala-Díaz, R.T.; Figueroa, F.L. UV Photoprotection, Cytotoxicity and Immunology Capacity of Red Algae Extracts. Molecules 2019, 24, 341. [Google Scholar] [CrossRef] [PubMed]
- Navarro, N.P.; Figueroa, F.L.; Korbee, N.; Mansilla, A.; Matsuhiro, B.; Barahona, T.; Plastino, E.M. The Effects of NO3 Supply on Mazzaella laminarioides (Rhodophyta, Gigartinales) from Southern Chile. Photochem. Photobiol. 2014, 90, 1299–1307. [Google Scholar] [CrossRef] [PubMed]
- Rosic, N.; Climstein, M.; Boyle, G.M.; Thanh Nguyen, D.; Feng, Y. Exploring Mycosporine-like Amino Acid UV-Absorbing Natural Products for a New Generation of Environmentally Friendly Sunscreens. Mar. Drugs 2023, 21, 253. [Google Scholar] [CrossRef]
- Eilers, P.H.C.; Peeters, J.C.H. A Model for the Relationship between Light Intensity and the Rate of Photosynthesis in Phytoplankton. Ecol. Modell. 1988, 42, 199–215. [Google Scholar] [CrossRef]
- Figueroa, F.L.; Conde-Álvarez, R.; Bonomi Barufi, J.; Celis-Plá, P.S.; Flores, P.; Malta, E.J.; Stengel, D.B.; Meyerhoff, O.; Pérez-Ruzafa, A. Continuous Monitoring of in Vivo Chlorophyll a Fluorescence in Ulva rigida (Chlorophyta) Submitted to Different CO2, Nutrient and Temperature Regimes. Aquat. Biol. 2014, 22, 195–212. [Google Scholar] [CrossRef]
- Grzymski, J.; Johnsen, G.; Sakshaug, E. The Significance of Intracellular Self-Shading on the Biooptical Properties of Brown, Red and Green Macroalgae. J. Phycol. 1997, 33, 408–414. [Google Scholar] [CrossRef]
- Figueroa, F.L.; Conde-Alvarez, R.; Gómez, I. Relations between Electron Transport Rates Determined by Pulse Amplitude Modulated Chlorophyll Fluorescence and Oxygen Evolution in Macroalgae under Different Light Conditions. Photosynth. Res. 2003, 75, 259–275. [Google Scholar] [CrossRef]
- Grasshoff, K.; Kremling, K.; Ehrhardt, M. Methods of Seawater Analysis; Wiley: Hoboken, NJ, USA, 1999. [Google Scholar]
- Massocato, T.F.; Robles-Carnero, V.; Moreira, B.R.; Castro-Varela, P.; Pinheiro-Silva, L.; Oliveira, W.D.; Vega, J.; Avilés, A.; Bonomi-Barufi, J.; Rörig, L.R.; et al. Growth, Biofiltration and Photosynthetic Performance of Ulva spp. Cultivated in Fishpond Effluents: An Outdoor Study. Front. Mar. Sci. 2022, 9, 1550. [Google Scholar] [CrossRef]
- Ritchie, R.J. Universal Chlorophyll Equations for Estimating Chlorophylls a, b, c, and d and Total Chlorophylls in Natural Assemblages of Photosynthetic Organisms Using Acetone, Methanol, or Ethanol Solvents. Photosynthetica 2008, 46, 115–126. [Google Scholar] [CrossRef]
- Sampath-Wiley, P.; Neefus, C.D. An Improved Method for Estimating R-Phycoerythrin and R-Phycocyanin Contents from Crude Aqueous Extracts of Porphyra (Bangiales, Rhodophyta). J. Appl. Phycol. 2007, 19, 123–129. [Google Scholar] [CrossRef]
- Folin, O.; Ciocalteu, V. On Tyrosine and Tryptophane Determinations in Proteins. J. Biol. Chem. 1927, 73, 627–648. [Google Scholar] [CrossRef]
- Vega, J.; Bárcenas-Pérez, D.; Fuentes-Ríos, D.; López-Romero, J.M.; Hrouzek, P.; Figueroa, F.L.; Cheel, J. Isolation of Mycosporine-like Amino Acids from Red Macroalgae and a Marine Lichen by High-Performance Countercurrent Chromatography: A Strategy to Obtain Biological UV-Filters. Mar. Drugs 2023, 21, 357. [Google Scholar] [CrossRef] [PubMed]
- Karsten, U.; Sawall, T.; Hanelt, D.; Bischof, K.; Figueroa, F.L.; Flores-Moya, A.; Wiencke, C. An Inventory of UV-Absorbing Mycosporine-like Amino Acids in Macroalgae from Polar to Warm-Temperate Regions. Bot. Mar. 1998, 41, 443–453. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Fv/Fm | αETR | ETRmax | Ek | ||
|---|---|---|---|---|---|---|
| Time | Radiation | NO3 | ||||
| Initial | - | - | 0.474 ± 0.07 | 0.054 ± 0.01 | 4.2 ± 0.7 | 79.3 ± 5.3 |
| 7 days | Amber | 60 | 0.506 ± 0.05 | 0.054. ± 0.03 | 5.4 ± 1.4 | 64.3 ± 8.6 |
| Amber | 240 | 0.536 ± 0.07 | 0.07 ± 0.01 | 6.2 ± 0.2 | 83.3 ± 8.1 | |
| Amber + UV-A | 60 | 0.526 ± 0.05 | 0.053 ± 0.02 | 6.4 ± 0.9 | 108.3 ± 29.1 | |
| Amber + UV-A | 240 | 0.498 ± 0.07 | 0.067 ± 0.02 | 5.8 ± 0.6 | 88.7 ± 15.2 | |
| Amber + Blue | 60 | 0.492 ± 0.04 | 0.058 ± 0.03 | 6.3 ± 1.1 | 82.5 ± 21.6 | |
| Amber + Blue | 240 | 0.589 ± 0.04 | 0.072 ± 0.01 | 6.9 ± 1.1 | 96.7 ± 23.5 | |
| Amber + Green | 60 | 0.603 ± 0.01 | 0.076 ± 0.01 | 6.4 ± 0.9 | 85.3 ± 17.9 | |
| Amber + Green | 240 | 0.500 ± 0.02 | 0.048 ± 0.01 | 6.4 ± 0.4 | 162.5 ± 12.5 | |
| 14 days | Amber | 60 | 0.527 ± 0.03 | 0.082 ± 0.01 | 7.7 ± 0.9 a | 94.1 ± 3.4 |
| Amber | 240 | 0.466 ± 0.06 | 0.068 ± 0.01 | 6.0 ± 0.4 a,b | 88.6 ± 12.2 | |
| Amber + UV-A | 60 | 0.492 ± 0.03 | 0.081 ± 0.01 | 7.3 ± 1.4 a,b | 89.7 ± 19.2 | |
| Amber + UV-A | 240 | 0.500 ± 0.05 | 0.070 ± 0.01 | 7.2 ± 0.7 a,b | 102.7 ± 13.6 | |
| Amber + Blue | 60 | 0.480 ± 0.06 | 0.088 ± 0.02 | 6.5 ± 1.7 a,b | 77.6 ± 30.0 | |
| Amber + Blue | 240 | 0.536 ± 0.06 | 0.074 ± 0.01 | 8.0 ± 1.5 a | 109.8 ± 29.1 | |
| Amber + Green | 60 | 0.551 ± 0.02 | 0.070 ± 0.01 | 4.6 ± 0.7 b | 65.7 ± 7.5 | |
| Amber + Green | 240 | 0.534 ± 0.10 | 0.073 ± 0.02 | 7.5 ± 0.7 a,b | 105.5 ± 18.4 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreira, B.R.; Vega, J.; García-Sánchez, M.; González-Fernández, C.; Avilés, A.; Bonomi-Barufi, J.; Figueroa, F.L. Photomorphogenic and Biochemical Effects of Radiation and Nitrate Availability on the Red Alga Plocamium cartilagineum. Plants 2025, 14, 1121. https://doi.org/10.3390/plants14071121
Moreira BR, Vega J, García-Sánchez M, González-Fernández C, Avilés A, Bonomi-Barufi J, Figueroa FL. Photomorphogenic and Biochemical Effects of Radiation and Nitrate Availability on the Red Alga Plocamium cartilagineum. Plants. 2025; 14(7):1121. https://doi.org/10.3390/plants14071121
Chicago/Turabian StyleMoreira, Bruna Rodrigues, Julia Vega, Marta García-Sánchez, Cristina González-Fernández, Antonio Avilés, José Bonomi-Barufi, and Félix L. Figueroa. 2025. "Photomorphogenic and Biochemical Effects of Radiation and Nitrate Availability on the Red Alga Plocamium cartilagineum" Plants 14, no. 7: 1121. https://doi.org/10.3390/plants14071121
APA StyleMoreira, B. R., Vega, J., García-Sánchez, M., González-Fernández, C., Avilés, A., Bonomi-Barufi, J., & Figueroa, F. L. (2025). Photomorphogenic and Biochemical Effects of Radiation and Nitrate Availability on the Red Alga Plocamium cartilagineum. Plants, 14(7), 1121. https://doi.org/10.3390/plants14071121