Effects of Bandwidth on Ear Differentiation and Grain Yield Formation of Maize in Strip Intercropping

 ,
,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
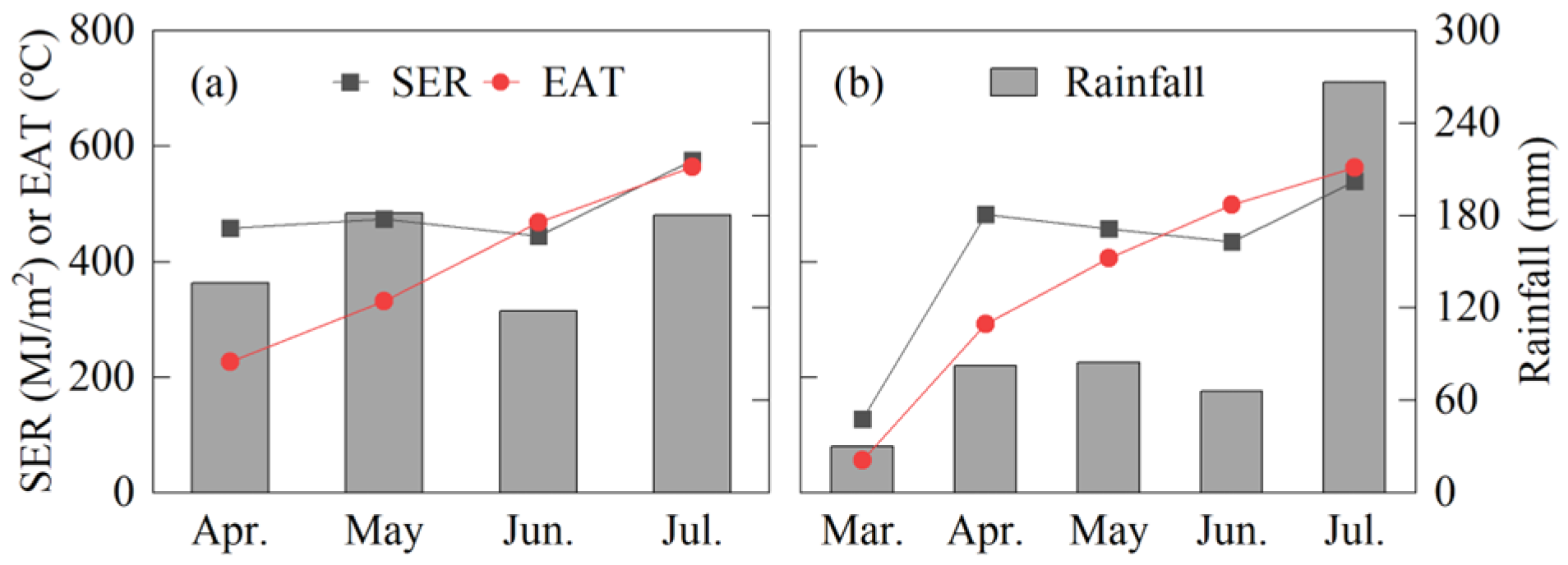
2.1. Plant Materials and Field Site
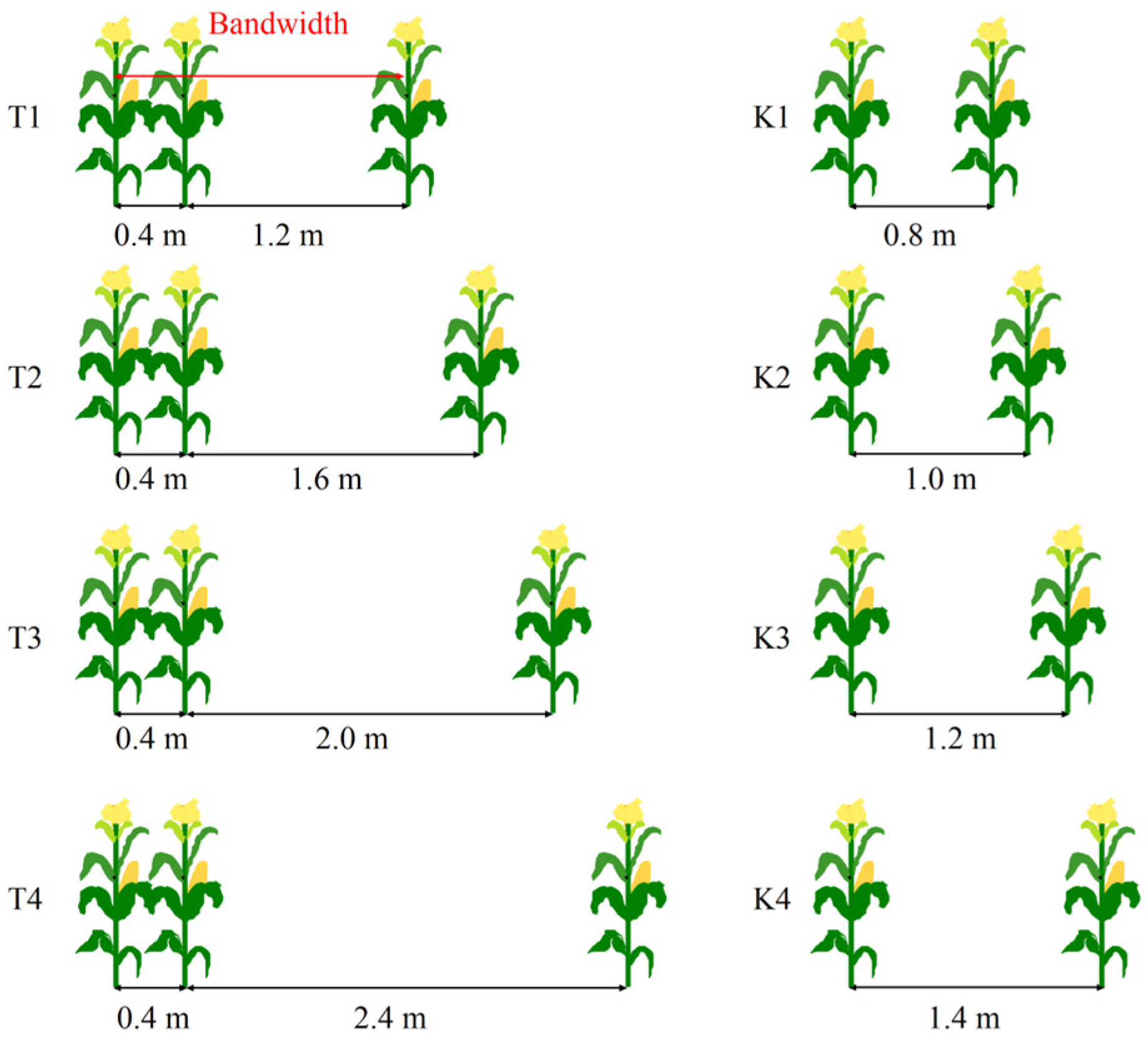
2.2. Experimental Design and Field Management
2.3. Sampling and Measurements
2.3.1. Light Transmittance of Maize Canopy
2.3.2. Ear Differentiation and Development
2.3.3. Maize Shoot Dry Matter
2.3.4. Yield and Yield Components
2.4. Statistical Analysis
3. Results
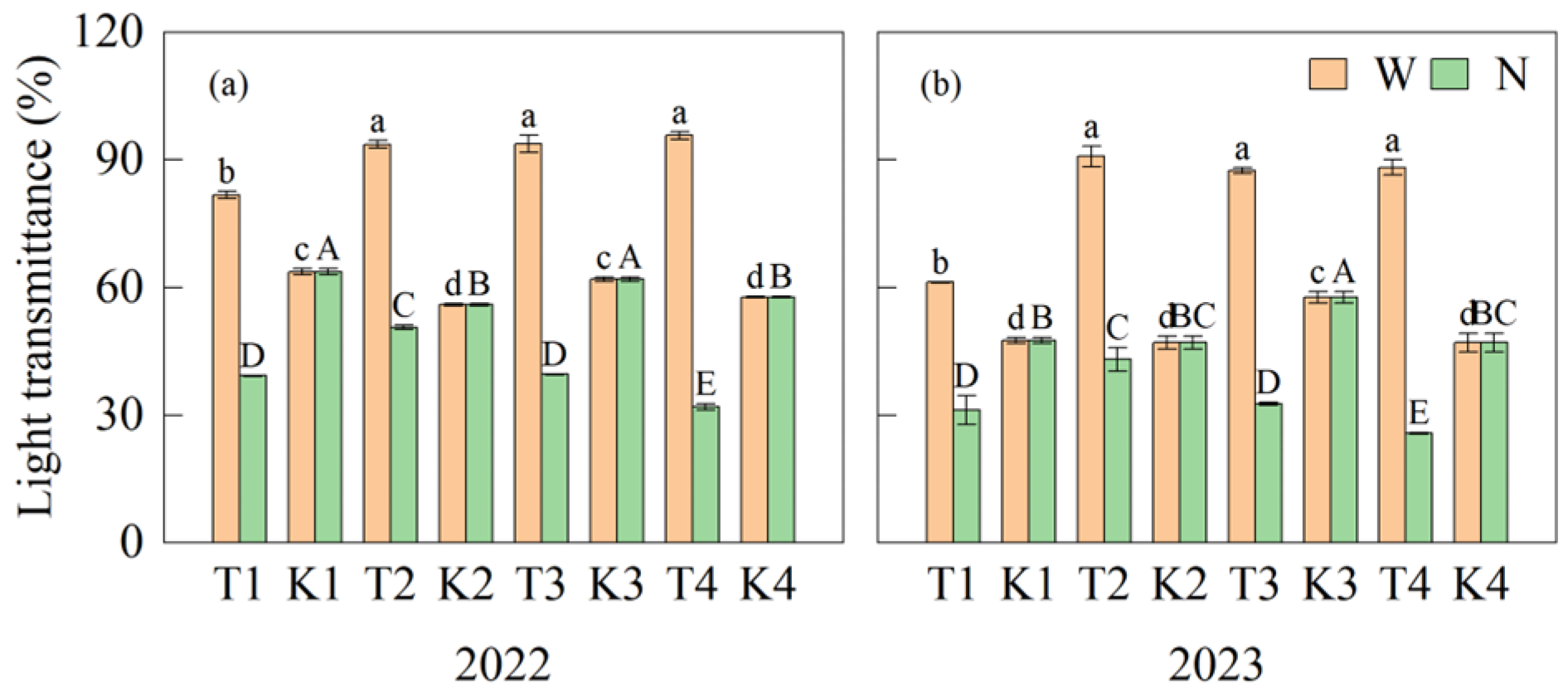
3.1. Differences in Light Transmittance of Wide Row and Narrow Row
3.2. Shoot Dry Matter Accumulation and Partitioning
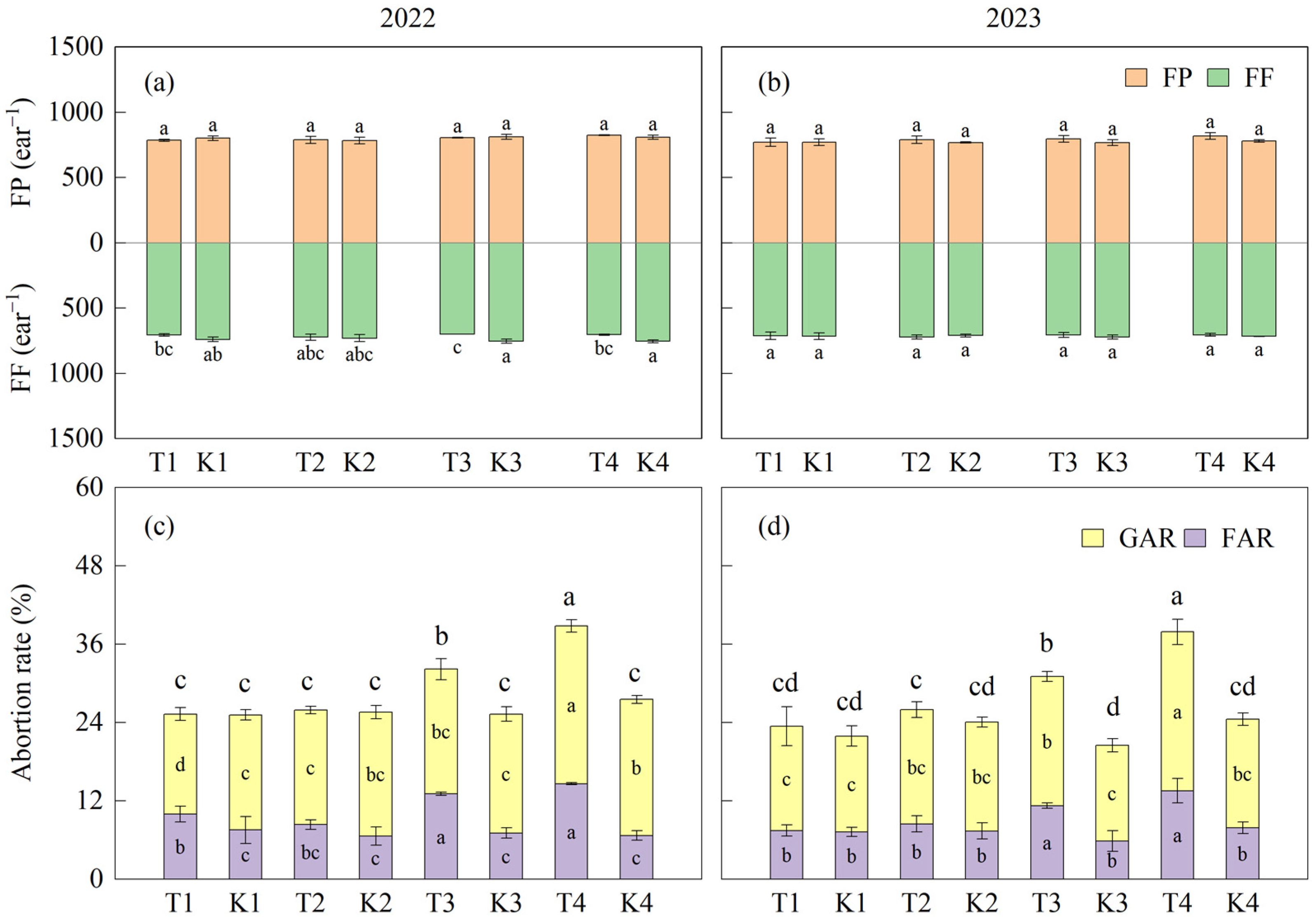
3.3. Differences in Ear Differentiation and Development
3.3.1. Ear Differentiation Process
3.3.2. Ear Differentiation Characteristics
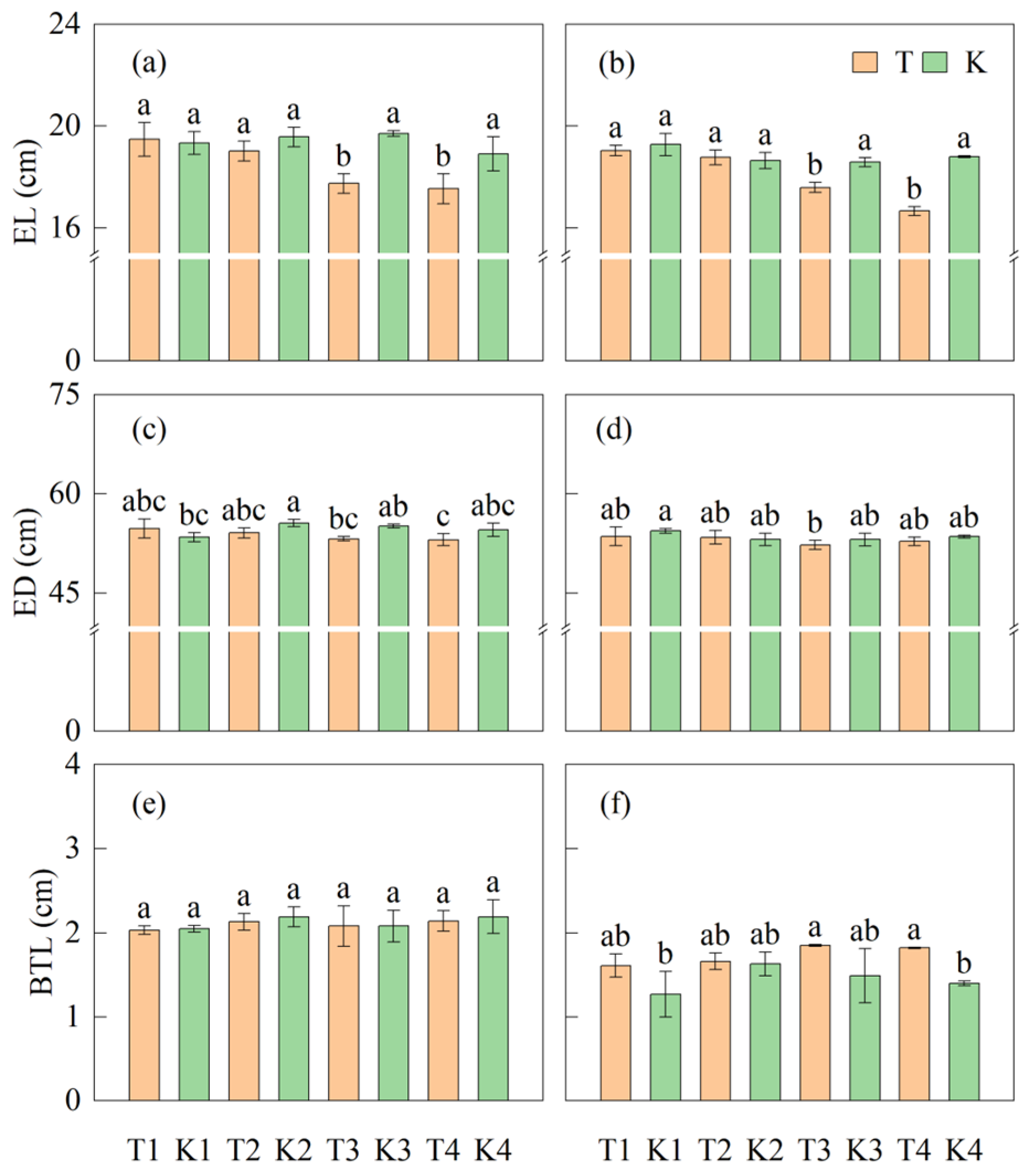
3.3.3. Ear Length and Diameter
3.4. Yield and Components
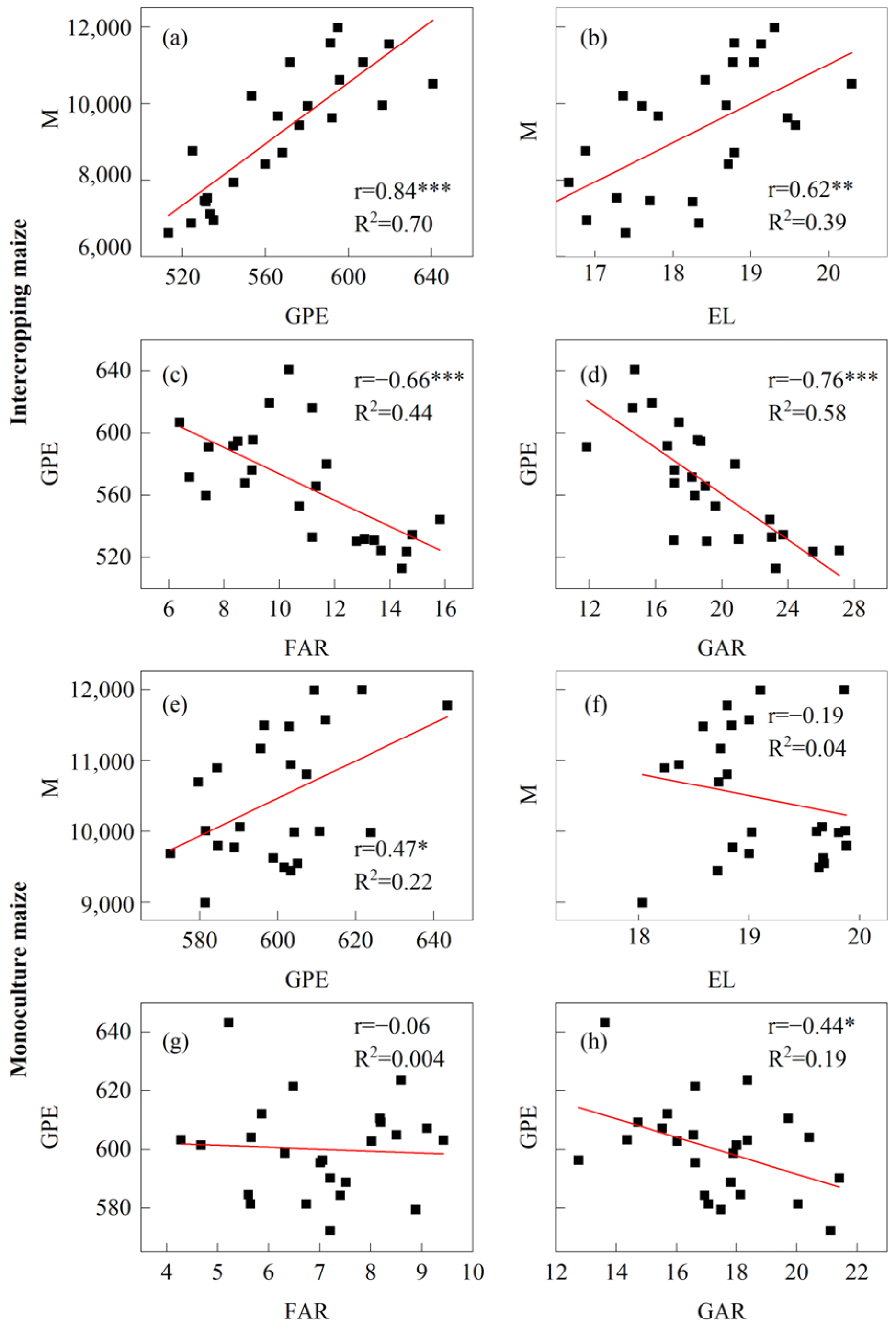
3.5. Correlation Analysis of Yield and Its Limiting Factors
4. Discussion
4.1. Appropriate Bandwidth Stable Dry Matter Partitioning to Ear
4.2. Intercropping Enhances the Sensitivity of Maize Floret Differentiation and Grain Development
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Li, G.; Liang, Y.; Liu, Q.; Zeng, J.; Ren, Q.; Guo, J.; Xiong, F.; Lu, D. Enhancing production efficiency through optimizing plant density in maize-soybean strip intercropping. Front. Plant Sci. 2024, 15, 1473786. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Sun, Z.; Su, Z.; Du, G.; Bai, W.; Wang, Q.; Wang, R.; Nie, J.; Sun, T.; Feng, C.; et al. Root plasticity and interspecific complementarity improve yields and water use efficiency of maize/soybean intercropping in a water-limited condition. Field Crops Res. 2022, 282, 108523. [Google Scholar] [CrossRef]
- Qian, X.; Zang, H.; Xu, H.; Hu, Y.; Ren, C.; Guo, L.; Wang, C.; Zeng, Z. Relay strip intercropping of oat with maize, sunflower and mung bean in semi-arid regions of Northeast China: Yield advantages and economic benefits. Field Crops Res. 2018, 223, 33–40. [Google Scholar] [CrossRef]
- Chen, G.; Liu, M.; Zhao, X.; Bawa, G.; Liang, B.; Feng, L.; Pu, T.; Yong, T.; Liu, W.; Liu, J.; et al. Improved photosynthetic performance under unilateral weak light conditions in a wide-narrow-row intercropping system is associated with altered sugar transport. J. Exp. Bot. 2024, 75, 258–273. [Google Scholar] [CrossRef]
- Raza, M.A.; Bin Khalid, M.H.; Zhang, X.; Feng, L.Y.; Khan, I.; Hassan, M.J.; Ahmed, M.; Ansar, M.; Chen, Y.K.; Fan, Y.F.; et al. Effect of planting patterns on yield, nutrient accumulation and distribution in maize and soybean under relay intercropping systems. Sci. Rep. 2019, 9, 4947. [Google Scholar] [CrossRef] [PubMed]
- Raza, M.A.; Feng, L.Y.; van der Werf, W.; Cai, G.R.; Khalid, M.H.B.; Iqbal, N.; Hassan, M.J.; Meraj, T.A.; Naeem, M.; Khan, I.; et al. Narrow-wide-row planting pattern increases the radiation use efficiency and seed yield of intercrop species in relay-intercropping system. Food Energy Secur. 2019, 8, e170. [Google Scholar] [CrossRef]
- Liu, X.; Rahman, T.; Song, C.; Yang, F.; Su, B.; Cui, L.; Bu, W.; Yang, W. Relationships among light distribution, radiation use efficiency and land equivalent ratio in maize-soybean strip intercropping. Field Crops Res. 2018, 224, 91–101. [Google Scholar] [CrossRef]
- Wang, Z.; Zhao, X.; Wu, P.; Gao, Y.; Yang, Q.; Shen, Y. Border row effects on light interception in wheat/maize strip intercropping systems. Field Crops Res. 2017, 214, 1–13. [Google Scholar] [CrossRef]
- Wang, R.; Sun, Z.; Zhang, L.; Yang, N.; Feng, L.; Bai, W.; Zhang, D.; Wang, Q.; Evers, J.B.; Liu, Y.; et al. Border-row proportion determines strength of interspecific interactions and crop yields in maize/peanut strip intercropping. Field Crops Res. 2020, 253, 107819. [Google Scholar] [CrossRef]
- Yang, F.; Liao, D.; Fan, Y.; Gao, R.; Wu, X.; Rahman, T.; Yong, T.; Liu, W.; Liu, J.; Du, J.; et al. Effect of narrow row planting patterns on crop competitive and economic advantage in maize soybean relay strip intercropping system. Plant Prod. Sci. 2017, 20, 1–11. [Google Scholar] [CrossRef]
- McCormick, A.J.; Cramer, M.D.; Watt, D.A. Sink strength regulates photosynthesis in sugarcane. New Phytol. 2006, 171, 759–770. [Google Scholar] [PubMed]
- Wu, H.; Tang, H.; Liu, L.A.; Shi, L.; Zhang, W.; Jiang, C. Local weak light induces the improvement of photosynthesis in adjacent illuminated leaves in maize seedlings. Physiol. Plant. 2021, 171, 125–136. [Google Scholar] [PubMed]
- Huang, S.; Ai, Y.; Du, J.; Yu, L.; Wang, X.; Yang, W.; Sun, X. Photosynthetic compensation of maize in heterogeneous light is impaired by restricted photosynthate export. Plant Physiol. Biochem. 2022, 192, 50–56. [Google Scholar]
- Sun, Z.; Geng, W.; Ren, B.; Zhao, B.; Liu, P.; Zhang, J. Responses of the photosynthetic characteristics of summer maize to shading stress. J. Agron. Crop Sci. 2023, 209, 330–344. [Google Scholar] [CrossRef]
- Zhang, M.; Chen, T.; Latifmanesh, H.; Feng, X.; Cao, T.; Qian, C.; Deng, A.; Song, Z.; Zhang, W. How plant density affects maize spike differentiation, kernel set, and grain yield formation in Northeast China? J. Integr. Agric. 2018, 17, 1745–1757. [Google Scholar]
- Gonzalez, V.H.; Lee, E.A.; Lukens, L.L.; Swanton, C.J. The relationship between floret number and plant dry matter accumulation varies with early season stress in maize (Zea mays L.). Field Crops Res. 2019, 238, 129–138. [Google Scholar]
- Hu, J.; Ren, B.; Dong, S.; Liu, P.; Zhao, B.; Zhang, J. Poor development of spike differentiation triggered by lower photosynthesis and carbon partitioning reduces summer maize yield after waterlogging. Crop J. 2022, 10, 478–489. [Google Scholar]
- Rattalino Edreira, J.I.; Budakli Carpici, E.; Sammarro, D.; Otegui, M.E. Heat stress effects around flowering on kernel set of temperate and tropical maize hybrids. Field Crops Res. 2011, 123, 62–73. [Google Scholar]
- Chen, T.; Song, Z.W.; Zhang, M.; Yan, X.G.; Zhu, P.; Ren, J.; Deng, A.X.; Zhang, W.J. Effects of shading and plant density on ear development and plant productivity of spring maize in Northeast China. J. Appl. Ecol. 2016, 27, 3237–3246. [Google Scholar]
- Li, Y.; Huang, S.; Meng, Q.; Li, Z.; Fritschi, F.B.; Wang, P. Pre-silking water deficit in maize induced kernel loss through impaired silk growth and ovary carbohydrate dynamics. Plant-Environ. Interact. 2024, 5, e10141. [Google Scholar]
- Liang, X.; Chen, H.; Pan, Y.; Wang, Z.; Huang, C.; Chen, Z.; Tang, W.; Chen, X.; Shen, S.; Zhou, S. Yield more in the shadow: Mitigating shading-induced yield penalty of maize via optimizing source-sink carbon partitioning. Eur. J. Agron. 2025, 162, 127421. [Google Scholar]
- Zhou, B.; Yue, Y.; Sun, X.; Ding, Z.; Ma, W.; Zhao, M. Maize kernel weight responses to sowing date-associated variation in weather conditions. Crop J. 2017, 5, 43–51. [Google Scholar]
- Gao, Z.; Sun, L.; Ren, J.; Liang, X.; Shen, S.; Lin, S.; Zhao, X.; Chen, X.; Wu, G.; Zhou, S. Detasseling increases kernel number in maize under shade stress. Agric. For. Meteorol. 2020, 280, 107811. [Google Scholar]
- Yang, Y.; Guo, X.; Liu, G.; Liu, W.; Xue, J.; Ming, B.; Xie, R.; Wang, K.; Hou, P.; Li, S. Solar Radiation Effects on Dry Matter Accumulations and Transfer in Maize. Front. Plant Sci. 2021, 12, 727134. [Google Scholar]
- Zhou, L.; Cao, J.; Zhang, F.; Li, L. Rhizosphere acidification of faba bean, soybean and maize. Sci. Total Environ. 2009, 407, 4356–4362. [Google Scholar]
- Liang, B.; Ma, Y.; Shi, K.; Chen, G.; Chen, H.; Hu, Y.; Chen, P.; Pu, T.; Wu, Y.; Sun, X.; et al. Appropriate bandwidth achieves a high yield by reducing maize intraspecific competition in additive maize-soybean strip intercropping. Eur. J. Agron. 2023, 142, 126658. [Google Scholar]
- Cui, H.; Camberato, J.J.; Jin, L.; Zhang, J. Effects of shading on spike differentiation and grain yield formation of summer maize in the field. Int. J. Biometeorol. 2015, 59, 1189–1200. [Google Scholar]
- Casal, J.J.; Fankhauser, C. Shade avoidance in the context of climate change. Plant Physiol. 2023, 191, 1475–1491. [Google Scholar]
- Maddonni, G.A.; Otegui, M.a.E.; Andrieu, B.; Chelle, M.; Casal, J.J. Maize leaves turn away from neighbors. Plant Physiol. 2002, 130, 1181–1189. [Google Scholar] [CrossRef]
- Zhou, Y.; Kusmec, A.; Schnable, P.S. Genetic regulation of self-organizing azimuthal canopy orientations and their impacts on light interception in maize. Plant Cell 2024, 36, 1600–1621. [Google Scholar] [CrossRef]
- Yang, Y.; Guo, X.; Liu, H.; Liu, G.; Liu, W.; Ming, B.; Xie, R.; Wang, K.; Hou, P.; Li, S. The effect of solar radiation change on the maize yield gap from the perspectives of dry matter accumulation and distribution. J. Integr. Agric. 2021, 20, 482–493. [Google Scholar]
- Fernández-Milmanda, G.L.; Ballaré, C.L. Shade Avoidance: Expanding the Color and Hormone Palette. Trends Plant Sci. 2021, 26, 509–523. [Google Scholar]
- Elango, T.; Jeyaraj, A.; Dayalan, H.; Arul, S.; Govindasamy, R.; Prathap, K.; Li, X. Influence of shading intensity on chlorophyll, carotenoid and metabolites biosynthesis to improve the quality of green tea: A review. Energy Nexus 2023, 12, 100241. [Google Scholar]
- Dörken, V.M.; Lepetit, B. Morpho-anatomical and physiological differences between sun and shade leaves in Abies alba Mill. (Pinaceae, Coniferales): A combined approach. Plant Cell Environ. 2018, 41, 1683–1697. [Google Scholar] [CrossRef]
- Gao, J.; Liu, Z.; Zhao, B.; Liu, P.; Zhang, J.W. Physiological and comparative proteomic analysis provides new insights into the effects of shade stress in maize (Zea mays L.). BMC Plant Biol. 2020, 20, 60. [Google Scholar] [CrossRef] [PubMed]
- Mathur, S.; Jain, L.; Jajoo, A. Photosynthetic efficiency in sun and shade plants. Photosynthetica 2018, 56, 354–365. [Google Scholar]
- Ren, B.; Yu, W.; Liu, P.; Zhao, B.; Zhang, J. Responses of photosynthetic characteristics and leaf senescence in summer maize to simultaneous stresses of waterlogging and shading. Crop J. 2023, 11, 269–277. [Google Scholar] [CrossRef]
- Taylor, S.H.; Gonzalez-Escobar, E.; Page, R.; Parry, M.A.J.; Long, S.P.; Carmo-Silva, E. Faster than expected Rubisco deactivation in shade reduces cowpea photosynthetic potential in variable light conditions. Nat. Plants 2022, 8, 118–124. [Google Scholar] [PubMed]
- Liang, X.; Gao, Z.; Shen, S.; Paul, M.J.; Zhang, L.; Zhao, X.; Lin, S.; Wu, G.; Chen, X.; Zhou, S. Differential ear growth of two maize varieties to shading in the field environment: Effects on whole plant carbon allocation and sugar starvation response. J. Plant Physiol. 2020, 251, 153194. [Google Scholar] [CrossRef]
- Yang, H.; Nukunya, K.; Ding, Q.; Thompson, B.E. Tissue-specific transcriptomics reveal functional differences in floral development. Plant Physiol. 2021, 188, 1158–1173. [Google Scholar]
- Yang, H.; Chai, Q.; Yin, W.; Hu, F.; Qin, A.; Fan, Z.; Yu, A.; Zhao, C.; Fan, H. Yield photosynthesis and leaf anatomy of maize in inter- and mono-cropping systems at varying plant densities. Crop J. 2022, 10, 893–903. [Google Scholar]
- Assefa, Y.; Carter, P.; Hinds, M.; Bhalla, G.; Schon, R.; Jeschke, M.; Paszkiewicz, S.; Smith, S.; Ciampitti, I. Analysis of Long Term Study Indicates Both Agronomic Optimal Plant Density and Increase Maize Yield per Plant Contributed to Yield Gain. Sci. Rep. 2018, 8, 4937. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.; Li, B.; Deng, T.; Xiao, Z.; Chen, X.; Hu, H.; Zhang, B.; Wu, G.; Li, F.; Zhao, X.; et al. The equilibrium between sugars and ethylene is involved in shading- and drought-induced kernel abortion in maize. Plant Growth Regul. 2020, 91, 101–111. [Google Scholar]
- Wang, J.; Zhao, S.; Zhang, Y.; Lu, X.; Du, J.; Wang, C.; Wen, W.; Guo, X.; Zhao, C. Investigating the genetic basis of maize ear characteristics: A comprehensive genome-wide study utilizing high-throughput phenotypic measurement method and system. Front. Plant Sci. 2023, 14, 1248446. [Google Scholar]
- Allison, J.C.S.; Daynard, T.B. Effect of Change in Time of Flowering, Induced by Altering Photoperiod or Temperature, on Attributes Related to Yield in Maize. Crop Sci. 1979, 19, 1–4. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Treatments | NPE ha−1 | GPE ear−1 | TGW g | M kg·ha−1 |
|---|---|---|---|---|---|
| 2022 | T1 | 56,252.81 a | 616.32 a | 289.52 ab | 10,037.45 a |
| K1 | 55,558.33 a | 603.30 ab | 283.39 ab | 9498.85 b | |
| T2 | 55,836.13 a | 568.03 c | 279.29 b | 8863.392 c | |
| K2 | 56,175.65 a | 604.22 ab | 292.55 a | 9923.72 ab | |
| T3 | 55,731.95 a | 531.15 d | 252.74 c | 7481.48 d | |
| K3 | 56,237.38 a | 597.00 ab | 294.35 a | 9877.86 ab | |
| T4 | 54,913.46 a | 523.96 d | 237.05 d | 6821.01 e | |
| K4 | 56,574.11 a | 581.37 bc | 291.30 ab | 9583.10 ab | |
| 2023 | T1 | 59,080.33 a | 597.73 a | 327.34 a | 11,553.25 ab |
| K1 | 58,300.53 a | 609.14 a | 333.01 a | 11,826.48 a | |
| T2 | 57,463.19 a | 595.66 a | 324.22 a | 11,089.74 ab | |
| K2 | 58,706.64 a | 592.08 ab | 317.92 a | 11,053.02 ab | |
| T3 | 57,141.3 a | 566.41 b | 307.3 a | 9939.21 c | |
| K3 | 58,683.49 a | 616.54 a | 315.14 a | 11,400.43 ab | |
| T4 | 56,499.42 a | 534.12 c | 263.70 b | 7943.78 d | |
| K4 | 55,785.10 a | 597.27 a | 318.29 a | 10,581.99 bc |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, B.; Li, J.; Zhao, X.; Lei, X.; Chen, G.; Pu, T.; Wu, Y.; Yong, T.; Yang, F.; Wang, X.; et al. Effects of Bandwidth on Ear Differentiation and Grain Yield Formation of Maize in Strip Intercropping. Plants 2025, 14, 1081. https://doi.org/10.3390/plants14071081
Liang B, Li J, Zhao X, Lei X, Chen G, Pu T, Wu Y, Yong T, Yang F, Wang X, et al. Effects of Bandwidth on Ear Differentiation and Grain Yield Formation of Maize in Strip Intercropping. Plants. 2025; 14(7):1081. https://doi.org/10.3390/plants14071081
Chicago/Turabian StyleLiang, Bing, Jingjing Li, Xuyang Zhao, Xinhui Lei, Guopeng Chen, Tian Pu, Yushan Wu, Taiwen Yong, Feng Yang, Xiaochun Wang, and et al. 2025. "Effects of Bandwidth on Ear Differentiation and Grain Yield Formation of Maize in Strip Intercropping" Plants 14, no. 7: 1081. https://doi.org/10.3390/plants14071081
APA StyleLiang, B., Li, J., Zhao, X., Lei, X., Chen, G., Pu, T., Wu, Y., Yong, T., Yang, F., Wang, X., & Yang, W. (2025). Effects of Bandwidth on Ear Differentiation and Grain Yield Formation of Maize in Strip Intercropping. Plants, 14(7), 1081. https://doi.org/10.3390/plants14071081