
Orchard Microclimate Control as a Way to Prevent Kiwifruit Decline Syndrome Onset
 ,
,  , , , , and
, , , , and 
Abstract
1. Introduction
2. Materials and Methods
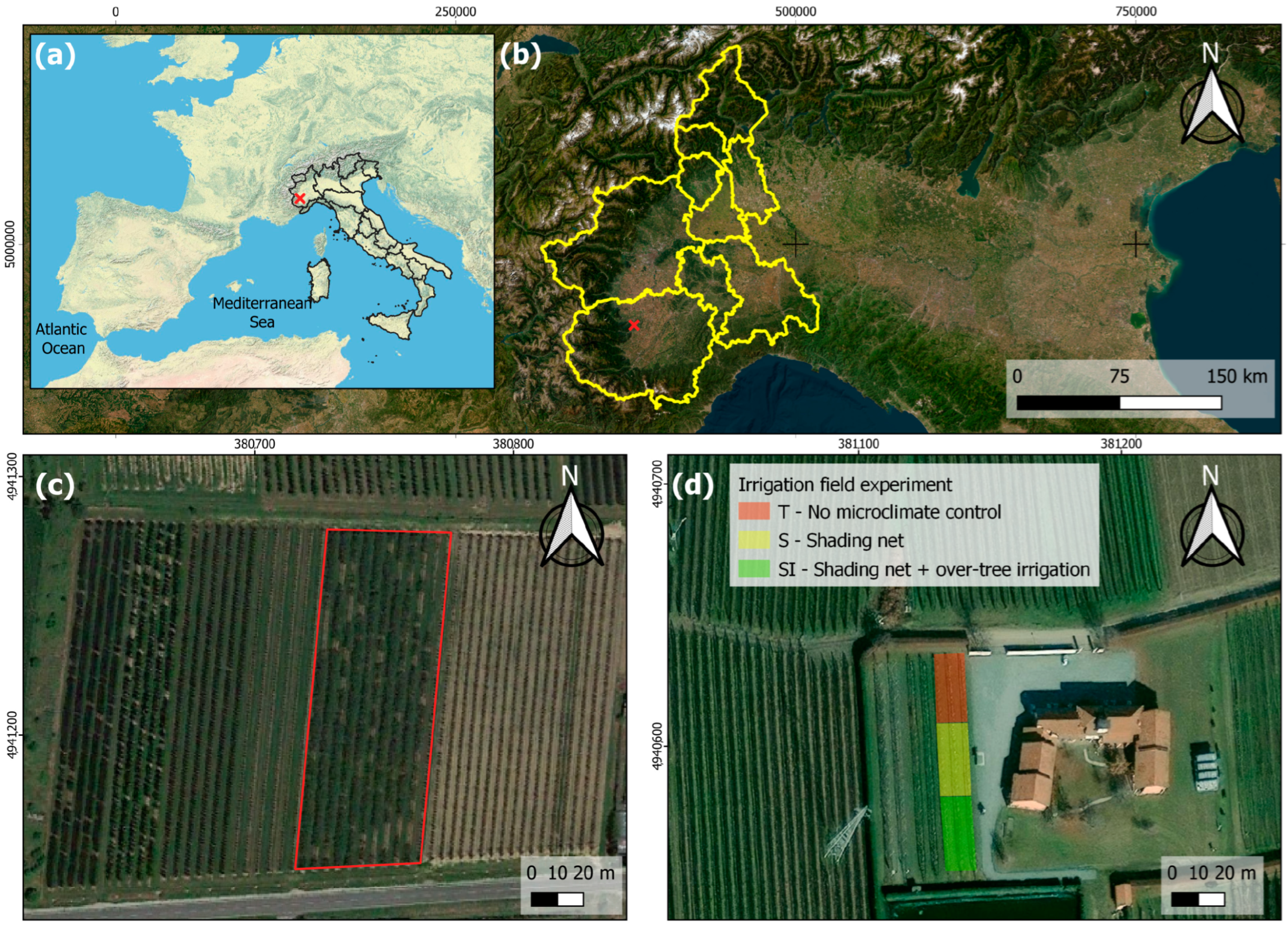
2.1. Study Site and Trial Setup
2.2. Environmental Measurements: Weather Data, Photosynthetically Active Radiation (PAR), and Microclimate
2.3. Physiological and Biometric Measurements
2.3.1. Leaf Gas Exchanges
2.3.2. Leaf Temperature
2.3.3. Stem Water Potential
2.3.4. Stem Growth
2.4. Root Xylem Vessel Characterization
2.5. Root Starch Content
2.6. Statistical Analysis
3. Results
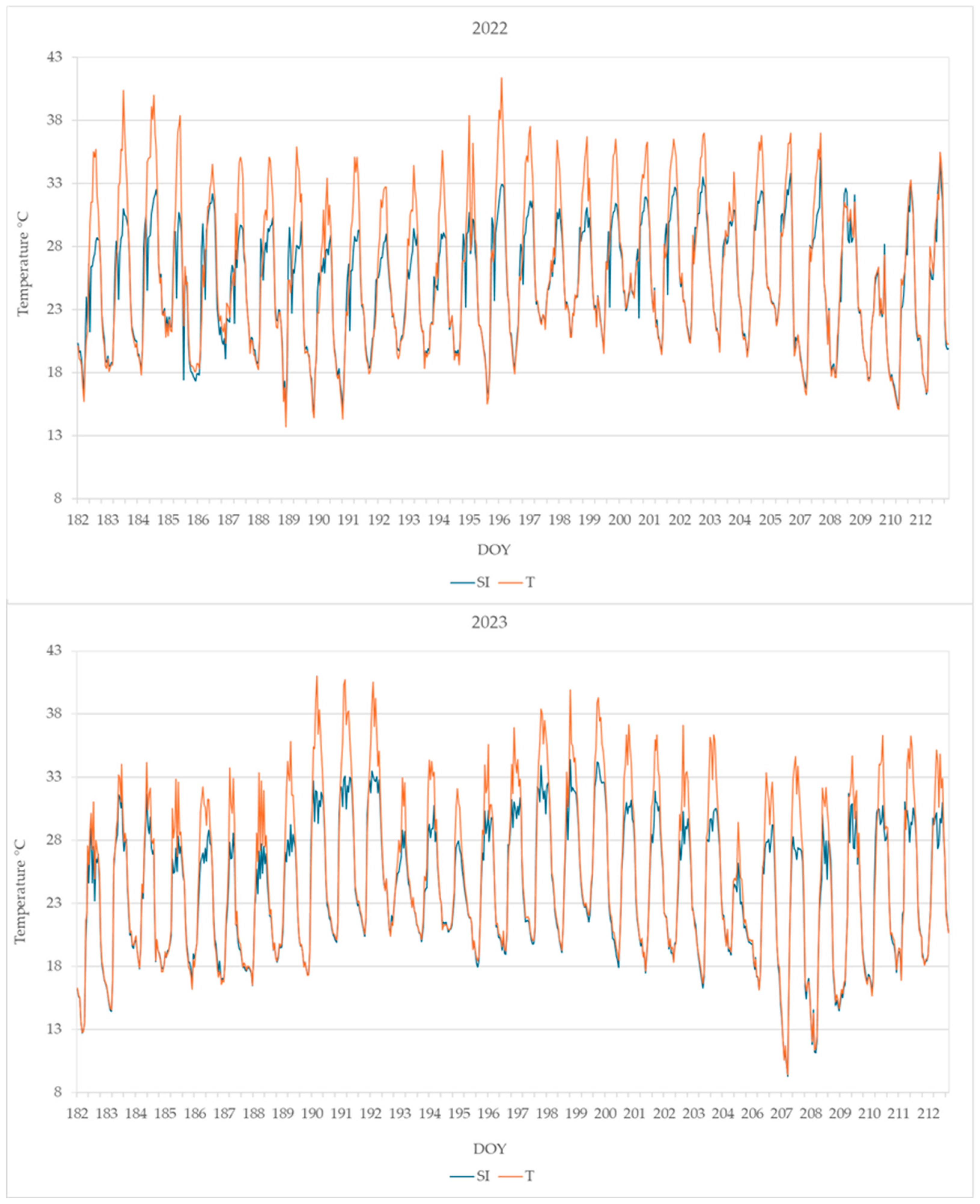
3.1. Weather Data, PAR, and Microclimate Measurements
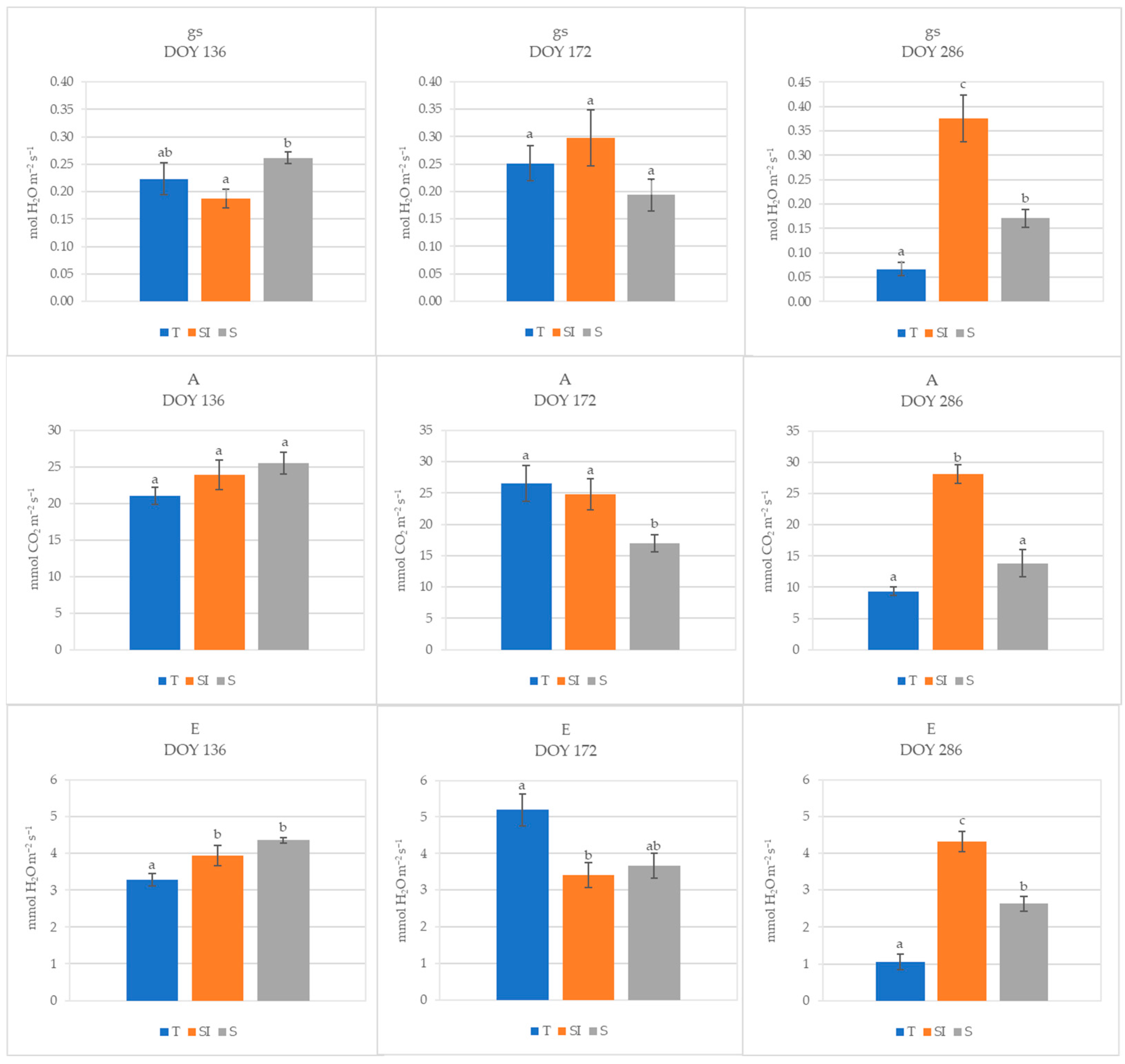
3.2. Leaf Gas Exchanges
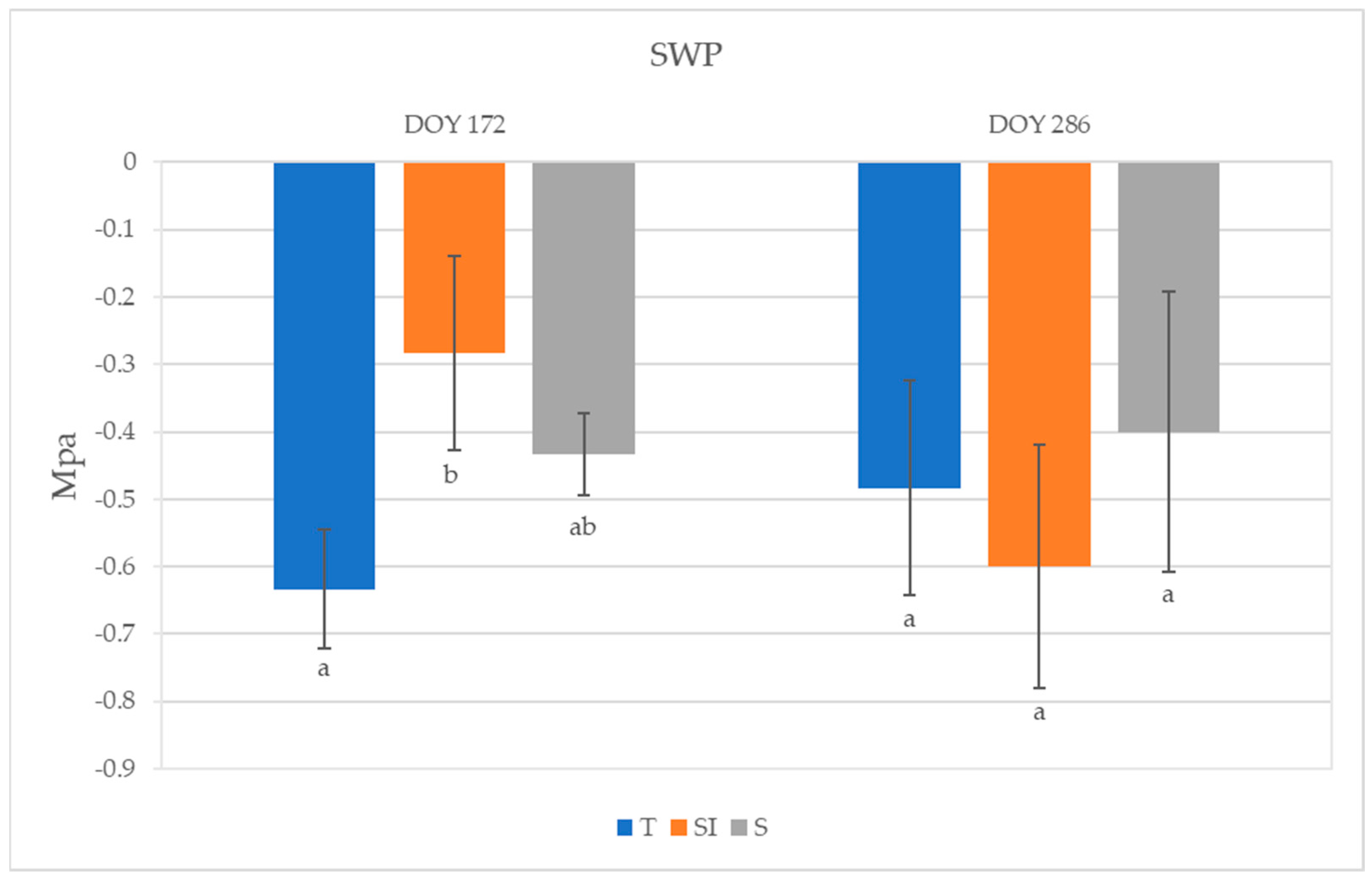
3.3. Stem Water Potential (SWP)
3.4. Leaf Temperature
3.5. Stem Growth
3.6. Roots Xylem Vessel Characterization
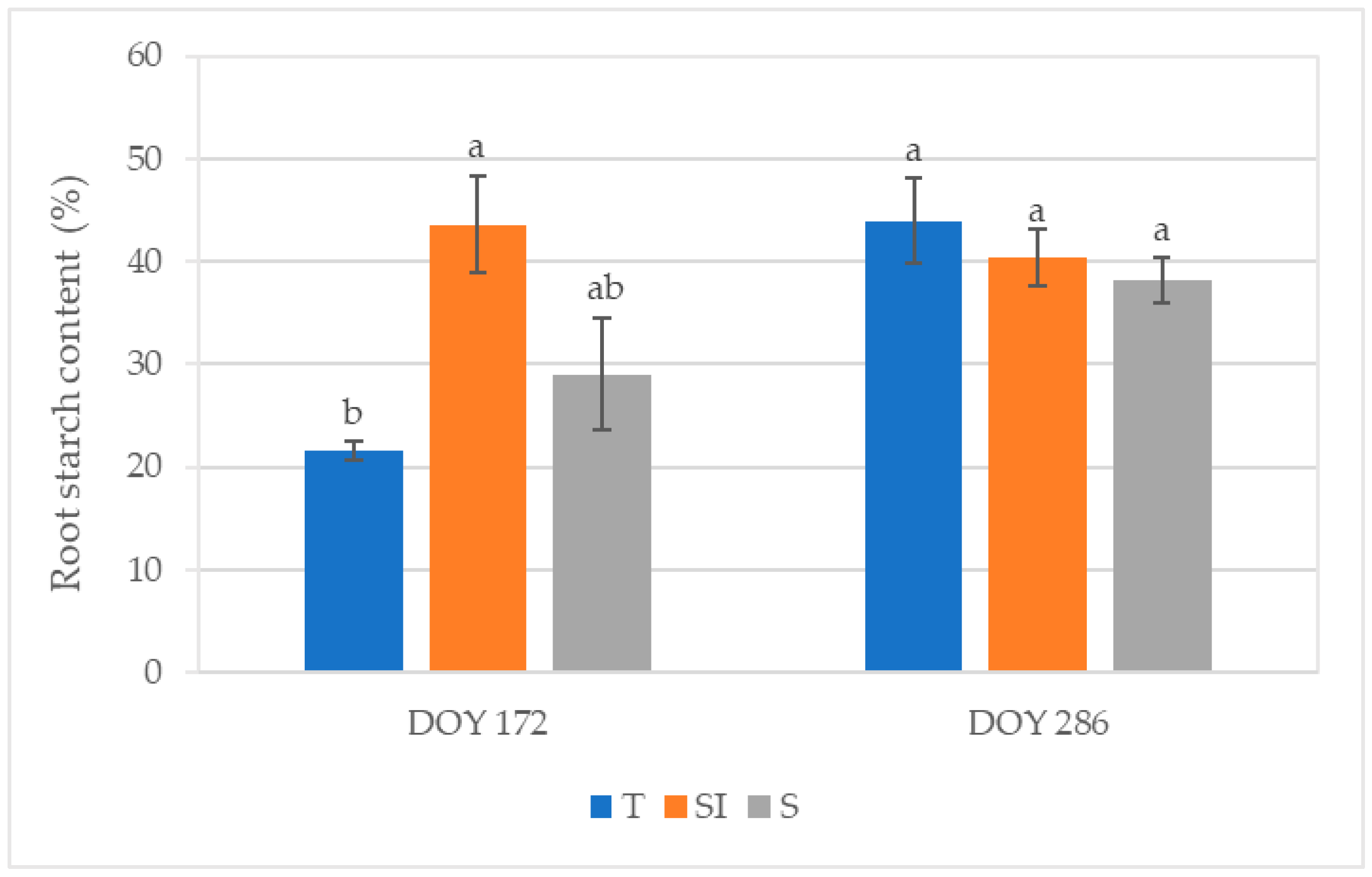
3.7. Root Starch Content
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 3 February 2025).
- Rajan, P.; Natraj, P.; Kim, M.; Lee, M.; Jang, Y.J.; Lee, Y.J.; Kim, S.C. Climate Change Impacts on and Response Strategies for Kiwifruit Production: A Comprehensive Review. Plants 2024, 13, 2354. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, A.R. Kiwifruit: A botanical review. Hortic. Rev. 1984, 6, 1–64. [Google Scholar] [CrossRef]
- Lemon, C.W.; Considine, J.A. Anatomy and Histochemistry of the Root System of the Kiwifruit Vine, Actinidia deliciosa var. deliciosa. Ann. Bot. 1993, 71, 117–129. [Google Scholar] [CrossRef]
- Condon, J.M. Aspects of kiwifruit stem structure in relation to transport. Acta Hortic. 1991, 297, 419–426. [Google Scholar] [CrossRef]
- Medda, S.; Fadda, A.; Mulas, M. Influence of Climate Change on Metabolism and Biological Characteristics in Perennial Woody Fruit Crops in the Mediterranean Environment. Horticulturae 2022, 8, 273. [Google Scholar] [CrossRef]
- Smith, G.S.; Buwalda, J.G.; Green, T.G.A.; Clark, C.J. Effect of oxygen supply and temperature at the root on the physiology of kiwifruit vines. New Phytol. 1989, 113, 431–437. [Google Scholar] [CrossRef]
- Smith, G.S.; Judd, M.J.; Miller, S.A.; Buwalda, J.G. Recovery of kiwifruit vines from transient waterlogging of the root system. New Phytol. 1990, 115, 325–333. [Google Scholar] [CrossRef]
- Bardi, L.; Nari, L.; Morone, C.; Faga, M.G.; Malusà, E. Possible Role of High Temperature and Soil Biological Fertility on Kiwifruit Early Decline Syndrome. Front. Agron. 2020, 2, 13. [Google Scholar] [CrossRef]
- Hall, A.; Snelgar, B. Temperature effects on kiwifruit dry matter at harvest. Acta Hortic. 2008, 803, 155–162. [Google Scholar] [CrossRef]
- Savé, R.; Serrano, L. Some physiological and growth responses of kiwi fruit (Actinidia chinensis) to flooding. Physiol. Plant. 1986, 66, 75–78. [Google Scholar] [CrossRef]
- Reid, J.B.; Tate, K.G.; Brown, N.S.; Cheah, L.H. Effects of flooding and alluvium deposition on kiwifruit (Actinidia deliciosa): 1. Early vine decline. N. Z. J. Crop Hortic. Sci. 1991, 19, 247–257. [Google Scholar] [CrossRef]
- Reid, J.B.; Tate, K.G.; Brown, N.S. Effects of flooding and alluvium deposition on kiwifruit (Actinidia deliciosa). N. Z. J. Crop Hortic. Sci. 1992, 20, 283–288. [Google Scholar] [CrossRef]
- Bardi, L.; Nari, L.; Morone, C.; Solomita, M.; Mandalà, C.; Faga, M.G.; Migliori, C.A. Kiwifruit Adaptation to Rising Vapor Pressure Deficit Increases the Risk of Kiwifruit Decline Syndrome Occurrence. Horticulturae 2022, 8, 906. [Google Scholar] [CrossRef]
- Mandalà, C.; Monaco, S.; Nari, L.; Morone, C.; Palazzi, F.; Bencresciuto, G.F.; Bardi, L. Xylem Hydraulic Conductance Role in Kiwifruit Decline Syndrome Occurrence. Horticulturae 2024, 10, 392. [Google Scholar] [CrossRef]
- Sengupta, S.; Majumder, A.L. Physiological and genomic basis of mechanical-functional trade-off in plant vasculature. Front Plant Sci. 2014, 5, 224. [Google Scholar] [CrossRef] [PubMed]
- Secchi, F.; Zwieniecki, M.A. Accumulation of sugars in the xylem apoplast observed under water stress conditions is controlled by xylem pH. Plant Cell Environ. 2016, 39, 2350–2360. [Google Scholar] [CrossRef]
- Guaschino, M.; Garello, M.; Nari, L.; Zhimo, Y.V.; Droby, S.; Spadaro, D. Soil, rhizosphere, and root microbiome in kiwifruit vine decline, an emerging multifactorial disease. Front. Microbiol. 2024, 15, 1330865. [Google Scholar] [CrossRef]
- Cardacino, A.; Turco, S.; Balestra, M.G. Seasonal dynamics of kiwifruit microbiome: A case study in a KVDS-affected orchard. Microbiol. Res. 2025, 292, 128044. [Google Scholar] [CrossRef]
- Bergamaschi, V.; Vera, A.; Pirone, L.; Siles, J.A.; López-Mondéjar, R.; Luongo, L.; Vitale, S.; Reverberi, M.; Infantino, A.; Bastida, F. Kiwifruit Vine Decline Syndrome (KVDS) Alters Soil Enzyme Activity and Microbial Community. Microorganisms 2024, 12, 2347. [Google Scholar] [CrossRef]
- Prencipe, S.; Schiavon, G.; Rosati, M.; Nari, L.; Schena, L.; Spadaro, D. Characterization of Phytopythium Species Involved in the Establishment and Development of Kiwifruit Vine Decline Syndrome. Microorganisms 2023, 11, 216. [Google Scholar] [CrossRef]
- Manici, L.M.; Saccà, M.L.; Scotti, C.; Caputo, F. Quantitative reduction of soil bacteria and qualitative microbial changes: Biotic components associated to kiwifruit decline. Plant Soil 2022, 477, 613–628. [Google Scholar] [CrossRef]
- Di Biase, R.; Calabritto, M.; Sofo, A.; Reyes, F.; Mininni, A.; Mastroleo, M.; Xylogiannis, E.; Dichio, B. Assessment of kiwifruit physiological decline: Irrigation and soil management strategy to recover from waterlogging. Acta Hortic. 2023, 1373, 11–18. [Google Scholar] [CrossRef]
- Donati, I.; Cellini, A.; Sangiorgio, D.; Caldera, E.; Sorrenti, G.; Spinelli, F. Pathogens Associated to Kiwifruit Vine Decline in Italy. Agriculture 2020, 10, 119. [Google Scholar] [CrossRef]
- Calabritto, M.; Mininni, A.N.; Di Biase, R.; Pietrafesa, A.; Dichio, B. Spatiotemporal dynamics of root water uptake and identification of soil moisture thresholds for precision irrigation in a Mediterranean yellow-fleshed kiwifruit orchard. Front. Plant Sci. 2024, 15, 1472093. [Google Scholar] [CrossRef]
- Mulas, M.; Fadda, A.; Zurru, R.; Chessa, G. Effectiveness of over tree irrigation to control temperature stress in SRA 63 clementines. Acta Hortic. 2004, 664, 413–418. [Google Scholar] [CrossRef]
- Iglesias, I.; Salvia, J.; Torguet, L.; Cabús, C. Orchard cooling with overtree microsprinkler irrigation to improve fruit colour and quality of ‘Topred Delicious’ apples. Sci. Hortic. 2022, 93, 39–51. [Google Scholar] [CrossRef]
- Kabir, M.Y.; Nambeesan, S.U.; Díaz-Pérez, J.C. Shade nets improve vegetable performance. Sci. Hortic. 2024, 334, 1113326. [Google Scholar] [CrossRef]
- Moura, L.; Pinto, R.; Rodrigues, R.; Brito, L.M.; Rego, R.; Valín, M.I.; Mariz-Ponte, N.; Santos, C.; Mourão, I.M. Effect of Photo-Selective Nets on Yield, Fruit Quality and Psa Disease Progression in a ‘Hayward’ Kiwifruit Orchard. Horticulturae 2022, 8, 1062. [Google Scholar] [CrossRef]
- Basile, B.; Giaccone, M.; Cirillo, C.; Ritieni, A.; Shahak, Y.; Forlani, M. Photo-selective hail nets affect fruit size and quality in Hayward kiwifruit. Sci. Hortic. 2012, 141, 91–97. [Google Scholar] [CrossRef]
- Basile, B.; Giaccone, M.; Shahak, Y.; Forlani, M.; Cirillo, C. Regulation of the vegetative growth of kiwifruit vines by photo-selective anti-hail netting. Sci. Hortic. 2014, 172, 300–307. [Google Scholar] [CrossRef]
- Calderón-Orellana, A.; Silva, D.I.; Bastías, R.M.; Bambach, N.; Aburto, F. Late-season plastic covering delays the occurrence of severe water stress and improves intrinsic water use efficiency and fruit quality in kiwifruit vines. Agric. Water Manag. 2021, 249, 106795. [Google Scholar] [CrossRef]
- Beck, H.E.; Zimmermann, N.E.; McVicar, T.R.; Vergopolan, N.; Berg, A.; Wood, E.F. Present and Future Köppen-Geiger Climate Classification Maps at 1-Km Resolution. Sci. Data 2018, 5, 180214. [Google Scholar] [CrossRef] [PubMed]
- Kottek, M.; Grieser, J.; Beck, C.; Rudolf, B.; Rubel, F. World Map of the Köppen-Geiger Climate Classification Updated. Meteorol. Z. 2006, 15, 259–263. [Google Scholar] [CrossRef]
- USDA. Keys to Soil Taxonomy, 11th ed.; United States Department of Agriculture, Natural Resources Conservation Service: Washington, DC, USA, 2010.
- FAO/ISRIC/ISSS. World Reference Base for Soil Resources; FAO: Rome, Italy, 1998. [Google Scholar]
- Scholander, P.F.; Bradstreet, E.D.; Hemmingsen, E.A.; Hammel, H.T. Sap Pressure in Vascular Plants. Science 1965, 148, 339–346. [Google Scholar] [CrossRef]
- Scholz, A.; Klepsch, M.; Karimi, Z.; Jansen, S. How to Quantify Conduits in Wood? Front. Plant Sci. 2013, 4, 56. [Google Scholar] [CrossRef]
- Eades, H.W. Iodine as an indicator of sapwood and heartwood. For. Chron. 1937, 13, 470–477. [Google Scholar] [CrossRef]
- Morabito, C.; Orozco, J.; Tonel, G.; Cavalletto, S.; Meloni, G.R.; Schubert, A.; Gullino, M.L.; Zwieniecki, M.A.; Secchi, F. Do the ends justify the means? Impact of drought progression rate on stress response and recovery in Vitis vinifera. Physiol. Plant. 2022, 174, e13590. [Google Scholar] [CrossRef]
- Pagliarani, C.; Casolo, V.; Ashofteh Beiragi, M.; Cavalletto, S.; Siciliano, I.; Schubert, A.; Gullino, M.L.; Zwieniecki, M.A.; Secchi, F. Priming xylem for stress recovery depends on coordinated activity of sugar metabolic pathways and changes in xylem sap pH. Plant Cell Environ. 2019, 42, 1775–1787. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2021, 4, 1–9. [Google Scholar]
- ARPA. Piemonte Regional Agency for Environmental Protection. Available online: https://www.arpa.piemonte.it (accessed on 4 December 2023).
- Blasini, D.E.; Koepke, D.F.; Bush, S.E.; Allan, G.J.; Gehring, C.A.; Whitham, T.G.; Day, T.A.; Hultine, K.R. Tradeoffs between leaf cooling and hydraulic safety in a dominant arid land riparian tree species. Plant Cell Environ. 2022, 45, 1664–1681. [Google Scholar] [CrossRef]
- Gong, X.; Leigh, A.; Guo, J.; Fang, L.D.; Hao, G.Y. Sand dune shrub species prioritize hydraulic integrity over transpirational cooling during an experimental heatwave. Agric. For. Meteorol. 2023, 336, 109483. [Google Scholar] [CrossRef]
- Eller, C.B.; Lima, A.L.; Oliveira, R.S. Foliar uptake of fog water and transport belowground alleviates drought effects in the cloud forest tree species, Drimys brasiliensis (Winteraceae). New Phytol. 2013, 199, 151–162. [Google Scholar] [CrossRef]
- Li, Z.; Gong, X.; Wang, J.L.; Chen, Y.D.; Liu, F.Y.; Li, H.P.; Lü, G.H. Foliar water uptake improves branch water potential and photosynthetic capacity in Calligonum mongolicum. Ecol. Indic. 2023, 146, 109825. [Google Scholar] [CrossRef]
- Carlquist, S. Ecological Strategies of Xylem Evolution. In Ecological Strategies of Xylem Evolution; University of California Press: Berkeley, CA, USA, 1975; ISBN 978-0-520-32056-7. [Google Scholar] [CrossRef]
- Hacke, U.G.; Sperry, J.S. Functional and Ecological Xylem Anatomy. Perspect. Plant Ecol. Evol. Syst. 2001, 4, 97–115. [Google Scholar] [CrossRef]
- Cochard, H.; Froux, F.; Mayr, S.; Coutand, C. Xylem Wall Collapse in Water-Stressed Pine Needles. Plant Physiol. 2004, 134, 401–408. [Google Scholar] [CrossRef]
- Grossiord, C.; Buckley, T.N.; Cernusak, L.A.; Novick, K.A.; Poulter, B.; Siegwolf, R.T.W.; Sperry, J.S.; McDowell, N.G. Plant Responses to Rising Vapor Pressure Deficit. New Phytol. 2020, 226, 1550–1566. [Google Scholar] [CrossRef]
- Gould, N.; Clearwater, M.J. Vine Canopy and Root Physiology. In Kiwifruit; CABI: Oxfordshire, UK, 2023; pp. 255–272. [Google Scholar] [CrossRef]
- Nari, L.; Berra, L.; Bardi, L.; Monaco, S.; Mandalà, C.; Morone, C.; Spadaro, D.; Guaschino, M.; Giorgi, V.; Neri, D. Misure Agronomiche Unica Strada Possibile per Prevenire la Morìa. Frutticoltura 2023, 7, 32–38. [Google Scholar]
- Grossiord, C.; Sevanto, S.; Borrego, I.; Chan, A.M.; Collins, A.D.; Dickman, L.T.; Hudson, P.J.; McBranch, N.; Michaletz, S.T.; Pockman, W.T.; et al. Tree Water Dynamics in a Drying and Warming World. Plant Cell Environ. 2017, 40, 1861–1873. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Site Description | |
|---|---|
| Coordinates | N 44.609001, E 7.501853 |
| Area | 2625 m2 |
| Planting distance | 5 m × 3 m |
| Year of establishment | 2020 |
| Cultivar | Actinidia chinensis var. deliciosa (Hayward) |
| Training system | T-Bar (pergola) |
| Shading net | Black-yellow anti-hail shading net |
| Propagation material | Cuttings |
| Soil preparation | Soil ridging |
| Previous crop | Kiwifruit (Hayward) |
| 2022 | 2023 | |||
|---|---|---|---|---|
| Treatment | SI | T | SI | T |
| June | 32.5 | 38.9 | 35.6 | 39.0 |
| July | 35.1 | 41.4 | 35.2 | 41.7 |
| August | 34.1 | 35.4 | 35.9 | 40.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mandalà, C.; Palazzi, F.; Bencresciuto, G.F.; Migliori, C.A.; Morabito, C.; Morone, C.; Nari, L.; Monaco, S.; Bardi, L. Orchard Microclimate Control as a Way to Prevent Kiwifruit Decline Syndrome Onset. Plants 2025, 14, 1049. https://doi.org/10.3390/plants14071049
Mandalà C, Palazzi F, Bencresciuto GF, Migliori CA, Morabito C, Morone C, Nari L, Monaco S, Bardi L. Orchard Microclimate Control as a Way to Prevent Kiwifruit Decline Syndrome Onset. Plants. 2025; 14(7):1049. https://doi.org/10.3390/plants14071049
Chicago/Turabian StyleMandalà, Claudio, Francesco Palazzi, Grazia Federica Bencresciuto, Carmela Anna Migliori, Cristina Morabito, Chiara Morone, Luca Nari, Stefano Monaco, and Laura Bardi. 2025. "Orchard Microclimate Control as a Way to Prevent Kiwifruit Decline Syndrome Onset" Plants 14, no. 7: 1049. https://doi.org/10.3390/plants14071049
APA StyleMandalà, C., Palazzi, F., Bencresciuto, G. F., Migliori, C. A., Morabito, C., Morone, C., Nari, L., Monaco, S., & Bardi, L. (2025). Orchard Microclimate Control as a Way to Prevent Kiwifruit Decline Syndrome Onset. Plants, 14(7), 1049. https://doi.org/10.3390/plants14071049